烏拉爾甘草TIFY基因家族鑒定及調控分析*
2022-09-29 15:19:48秦振芬孟祥霄朱雪雯BotirKhaitovAtiatulWahab
世界科學技術-中醫藥現代化 2022年5期
秦振芬,孟祥霄,溫 東,朱雪雯,王 艷,Botir Khaitov,Atia tul Wahab,孫 偉**
(1. 中國中醫科學院中藥研究所/中藥鑒定與安全性評估北京市重點實驗室 北京 100700;2. 中國醫學科學院北京協和醫學院藥用植物研究所 北京 100193;3. 卡拉奇大學國際化學與生命科學研究院化學研究所卡拉奇 75270;4. 國際生物鹽農業中心 塔什干 100084;5. 卡拉奇大學國際化學與生命科學研究院分子醫學與藥物研究所 卡拉奇 75270)
中藥材甘草是指甘草屬(Glycyrrhiza)多年生草本植物烏拉爾甘草(G. uralensisFisch.)、光果甘草(G.glabraL.)和脹果甘草(G. inflataBatalin.)的干燥根及根莖,性平味甘,歸脾、胃、心、肺經,為臨床常用藥,具有補中益氣、潤肺止咳、清熱解毒、緩急止痛、調和諸藥之功。因其能調和百藥、解百毒,故有“國老”之稱[1]。我國是對甘草認識、研究最早的國家,甘草最早出現于《神農本草經》中,并被列為上品[2]。烏拉爾甘草主要活性成分包括甘草皂苷類、甘草黃酮類以及甘草多糖,常用于治療多方面疾病如心腦血管疾病、炎癥等[3]。其中甘草酸的含量是《中國藥典》中衡量甘草藥材品質優劣的主要指標。甘草酸具有抗癌、抗病毒、免疫調節等藥理作用,還具有促腎上腺皮質激素樣的作用,同時在臨床上可作為消炎藥使用[4]。因此深入研究甘草酸的轉錄調控機制,對于培育高質量植株,促進甘草酸生物合成及開發甘草酸類藥物等具有重要意義。
三萜皂苷的生物合成首先通過氧化角鯊烯環化酶(OSCs)將共同前體2,3-氧化角鯊烯環化成各種各樣的三萜骨架模式。這些三萜骨架在細胞色素P450單加氧酶(P450)的催化下進行位點特異性氧化,形成不同的三萜苷元或非糖基化形式。甘草酸苷是齊墩烷型三萜皂苷,來源于β-香樹脂醇,近年來,隨著對甘草酸代謝途徑研究的不斷深入,其生物合成酶包括P450酶、UGT 合成酶和細胞合成酶介導的葡萄糖醛酸基轉移酶大多已被鑒定[5],同時,研究人員已在酵母中成功合成了甘草酸的前體物質甘草次酸[6](圖1)。相較烏拉爾甘草藥效成分的合成途徑解析,轉錄調控研究相對較少,如bHLH 類轉錄因子GubHLH3 可以調控烏拉爾甘草中大豆皂苷的合成[7],因此需要我們更深入研究烏拉爾甘草藥效成分的轉錄調控機制。
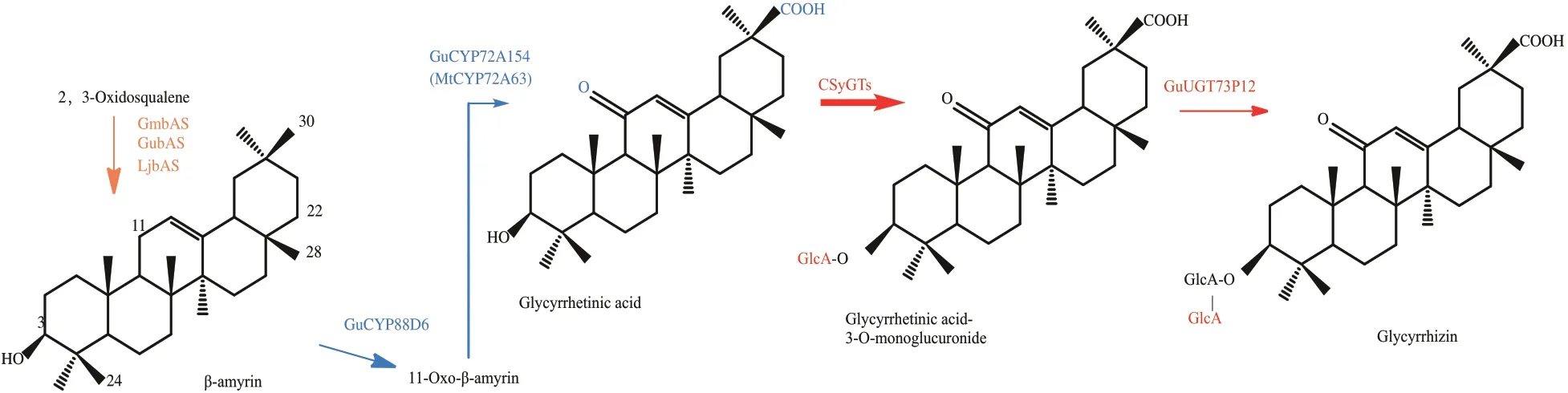
圖1 甘草酸的代謝途徑
TIFY 家族基因是近年發現的能增強植物抗逆性的新基因。TIFY 蛋白是植物中一類特異性轉錄因子,最初在擬南芥中鑒定出來,屬于植物特異的GATA 轉錄因子家族,TIFY 家族的特征是一個高度保守的基序(TIF[F/Y]XG),所以被命名為TIFY 基因家族[8]。通過系統發育和結構分析,該家族的基因可分為:TIFY、JAZ、ZML 和PPD[9]。其中,最典型的成員是JAZ 基因,是植物特異性基因家族的成員,在JA通路中發揮著關鍵作用[10]。茉莉酸類化合物廣泛存在于植物體內,是一種植物內源激素,在植物各個生長階段以及抵抗逆境過程中均起到重要作用[11]。Nishii 等從擬南芥中鑒定出首個TIFY 基因,隨后TIFY 基因家族陸續在水稻、大麻、人參、鐵皮石斛、杜仲等中被鑒定出來[12-16]。近期研究表明,TIFY 基因家族參與眾多生物學過程,例如擬南芥中過表達AtTIFY1(ZIM)基因導致葉柄和下胚軸加長[17]、AtTIFY4a(PPD1)和AtTIFY4b(PPD2)基因有助于擬南芥葉片同步生長[18];Wang Y[19]在木本模式植物毛果楊中發現,TIFY 基因可積極響應干旱、鹽分、極端溫度等非生物脅迫;Chini A[20]在番茄中發現,TIFY 基因對鹽脅迫,滲透脅迫和ABA 處理都有反應;在藥用活性成分長春花堿的代謝調控研究中顯示,缺少茉莉酸甲酯刺激下JAZ 蛋白可以有效地抑制CrMYC2和BIS1表達從而控制長春花堿的合成[21];在藥用植物丹參中JAZ3 蛋白可以和SmWD40-170 轉錄因子蛋白形成復合體,從而阻礙其正向調控丹參酮合成的作用[22]。
由此可見,TIFY 蛋白在植物生長發育、代謝物質調控上具有重要作用,因此本研究在已有的烏拉爾甘草全基因組的基礎上,采用生物信息學方法對烏拉爾甘草TIFY 基因家族的序列特征、分類、系統發育和基因的組織差異表達進行可視化分析,為研究TIFY基因家族成員參與甘草酸代謝調控提供數據支持,為甘草的優良品種選育等工作提供參考。
1 材料
烏拉爾甘草的全基因組數據和相關注釋在http://ngs-data-archive.psc.riken.jp/Gur-genome/download.pl網站獲取,其品種為308-19 品系烏拉爾甘草[23]。擬南芥TIFY 基因家族蛋白數據下載自TAIR 網站(http://www.arabidopsis.org/browse/genefamily/index.jsp)。
2 方法
2.1 烏拉爾甘草GurTIFY基因家族鑒定
烏拉爾甘草全部蛋白序列通過TBtools 軟件進行分析提取,與擬南芥TIFY基因家族蛋白序列進行blast比對,獲得候選烏拉爾甘草TIFY 基因序列,用TBtools去除重復,然后包含TIFY 結構域(PF06200)的蛋白通過HMMERsearch 獲得,其他數據刪除,確定烏拉爾甘草中的TIFY 基因家族成員。利用ExPASy 網站(http://web.expasy.org/protparam/)對烏拉爾甘草TIFY等電點、蛋白分子量等理化性質進行分析。亞細胞定位通過WoLFPSORT 網站(http://www.genscript.com/wolfpsort.html)進行分析。
2.2 烏拉爾甘草GurTIFY基因家族系統進化分析
使用MEGA7軟件對擬南芥和烏拉爾甘草的TIFY蛋白進行多序列比對,使用鄰接法(Neighbor-Joining,NJ)構建系統發育樹(bootstrap 重復次數為1000),并用iTOL(http://itol.embl.de/)網站對進化樹進行美化,根據系統發育樹對烏拉爾甘草GurTIFY基因亞家族分類。
2.3 烏拉爾甘草GurTIFY 基因保守基序及基因結構分析
通 過MEME(https://meme-suite. org/meme/)對GurTIFY的蛋白保守基序進行預測(motif 參數設為10),使用CDD 數據庫(https://www.ncbi.nlm.nih.gov/cdd)對GurTIFY的蛋白結構域進行預測。將蛋白保守基序、保守結構域和其系統進化樹使用TBtools軟件整合及可視化作圖。
2.4 烏拉爾甘草GurTIFY基因順式作用元件預測
通過基因組序列和結構注釋文件分別提取18 個GurTIFY基因上游各2Kb 序列作為啟動子區域。然后對啟動子區的順式作用元件的預測通過PlantCare(https://www.plantcare.co.uk/)進行,通過TBtools 軟件進行可視化作圖。
2.5 烏拉爾甘草GurTIFY基因差異表達分析
采用兩年生烏拉爾甘草根和葉的轉錄組數據,使用STAR(version 2.7.3)軟件進行轉錄組數據比對,使用RSEM(version 1.33)軟件計算各樣品的表達量FPKM 值。使用DEseq2(version 1.22.2)軟 件 計 算GurTIFY在不同組織中差異表達情況。最終使用烏拉爾甘草GurTIFY在不同組織的FPKM 值,繪制TIFY 基因表達水平的柱形圖。轉錄組原始數據為已上傳至國家基因組科學數據中心(https://ngdc.cncb.ac.cn/),GSA編號:CRA005924。
篩選表達量較高,且不同組織間具有顯著差異的基因進行實時熒光定量PCR(qRT-PCR)分析,使用華越洋試劑盒(0416-50)提取樣品RNA,全式金試劑盒(AT311-02)反轉錄合成cDNA,采用啟衡星試劑盒(FS-Q1002)進 行qRT-PCR 反應,內參基因為βtubulin,使用NCBI 設計引物(表1),qRT-PCR 反應體系:cDNA 模板1 μL,上下游引物各0.4 μL、SYBR qPCR Ready Mix 熒光染料10 μL,ddH2O 8.2 μL。反應程序:95℃預變性3 min;95℃變性30 s,56℃退火20 s,72℃延伸20 s,40 次循環。每個樣品設3 次生物學重復,采用2-ΔΔCT法分析基因表達量。
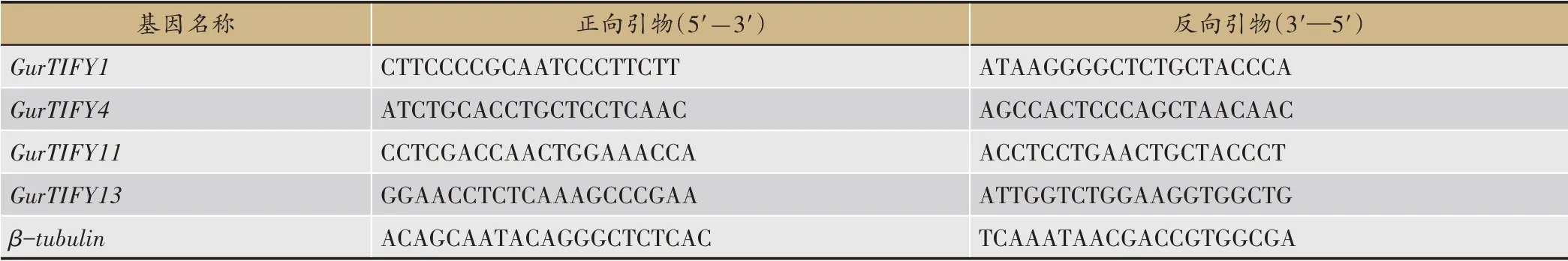
表1 烏拉爾甘草GurTIFY基因qRT-PCR引物序列
3 結果與分析
3.1 烏拉爾甘草GurTIFY基因家族成員的鑒定
通過AtTIFY的blastn 同源比對,在烏拉爾甘草中共得到37 條候選序列,去除重復并確認結構域(PF06200)后 得 到GurTIFY成 員18 個,命 名 為GurTIFY1-GurTIFY18,核苷酸長度在444-1266 bp 之間,氨基酸長度在147-421 aa 之間,蛋白分子量大小在16741.11-44426.5 Da 之間,等電點介于4.57-9.68之間,與此前已發表的擬南芥,大麻等的TIFY 基因家族相近(表2)。

表2 烏拉爾甘草TIFY基因家族基本信息
3.2 烏拉爾甘草GurTIFY 基因的蛋白多重序列分析和系統進化分析
為了解烏拉爾甘草和擬南芥TIFY 基因家族的進化關系,烏拉爾甘草和擬南芥TIFY 基因共37 個,根據擬南芥TIFY 家族分類方法[24]被分為4 個亞類(圖2)。烏拉爾甘草18 個TIFY 基因中,JAZ 亞家族基因10 個,將JAZ亞家族分為3個亞組,Ⅰ組有GurTIFY8,Ⅱ組由GurTIFY4、GurTIFY17、GurTIFY18組 成,Ⅲ組 由GurTIFY1、 GurTIFY2、 GurTIFY10、 GurTIFY11、GurTIFY13、GurTIFY15組成;ZML亞家族基因5個分別為GurTIFY12、GurTIFY6、GurTIFY16、GurTIFY9、GurTIFY7;TIFY 亞家族基因2 個分別為GurTIFY5和GurTIFY3以及PPD 亞家族基因1 個為GurTIFY14,每個亞家族的結構域特點與擬南芥一致。

圖2 烏拉爾甘草(紅色字體)和擬南芥(黑色字體)TIFY轉錄因子系統發育樹
3.3 烏拉爾甘草GurTIFY 基因保守基序及基因結構分析
將烏拉爾甘草TIFY蛋白保守基序、保守結構域和其系統進化樹使用TBtools 軟件進行結合并做可視化作圖分析。結果顯示,烏拉爾甘草的18 個TIFY 基因中含有10 個保守基序和4 個保守結構域,18 個GurTIFY中 均 含 有 TIFY 結 構 域,GurTIFY9、GurTIFY16、GurTIFY7、GurTIFY6、GurTIFY12含有CCT和GATA 結構域,除GurTIFY9、GurTIFY16、GurTIFY7、GurTIFY6、GurTIFY12、GurTIFY3、GurTIFY5外均含有Jas結構域(圖3)。

圖3 烏拉爾甘草TIFY家族功能結構域
為進一步分析甘草GurTIFY基因之間的進化關系,利用TBtools 軟件構建了烏拉爾甘草GurTIFY的基因結構圖。結果顯示,烏拉爾甘草的18 個GurTIFY中內含子的數量為2-9 個(圖4),其中GurTIFY9和GurTIFY16所含內含子數量最多為9 個,GurTIFY14含有2 個內含子,數目最少,其余成員中GurTIFY1、GurTIFY2、GurTIFY11、GurTIFY13、GurTIFY15含內含子4 個,GurTIFY4、GurTIFY6、GurTIFY12、GurTIFY17、GurTIFY18含內含子6 個,GurTIFY3和GurTIFY5含內含子5 個,GurTIFY10、GurTIFY7和GurTIFY8內含子數量分別為3、7、8個。

圖4 烏拉爾甘草GurTIFY基因的基因結構
3.4 烏拉爾甘草GurTIFY基因順式作用元件預測
順式元件對基因表達具有重要的調控作用,烏拉爾甘草TIFY 基因家族成員的順式作用元件預測和可視化分析結果表明(圖5),GurTIFY中每個成員的啟動子區都含有眾多順式元件,如光響應元件,脫落酸響應元件,干旱誘導元件等。光響應元件數量最多為274 個,分布在每個成員中,其次為脫落酸響應元件數量為68個。另外,烏拉爾甘草抗逆性和抗干旱的茉莉酸甲酯和干旱響應元件也在大多數烏拉爾甘草TIFY基因的啟動子上存在。表明烏拉爾甘草TIFY 不同成員能夠廣泛且差異性的參與烏拉爾甘草生長發育及非生物脅迫等多種生物過程的響應。

圖5 烏拉爾甘草GurTIFY基因的啟動子區順式作用元件
3.5 烏拉爾甘草GurTIFY基因差異表達分析
為了探究GurTIFY在烏拉爾甘草不同組織中的表達模式,對兩年生烏拉爾甘草根和葉的轉錄組數據進行分析,結果表明(圖6),GurTIFY15、GurTIFY4、GurTIFY1、 GurTIFY11、 GurTIFY5、 GurTIFY13、GurTIFY8、GurTIFY16、GurTIFY14在烏拉爾甘草葉和根中的表達水平具有顯著差異性。GurTIFY15、GurTIFY4在各個組織表達量都相對較高,而GurTIFY16、GurTIFY3和GurTIFY14在各個組織幾乎不表達。且GurTIFY15、GurTIFY4、GurTIFY1、GurTIFY11、GurTIFY13在葉中的表達遠遠高于在根中的表達。在所有成員中GurTIFY15在葉表達量最高,GurTIFY2 在根表達量最高。以上在根和葉中表達量較高的基因大多為JAZ 亞家族的成員,表明烏拉爾甘草的GurTIFY基因家族成員中的JAZ 亞家族可能參與了不同組織器官的調控。篩選表達量較高,且組織間具有顯著差異的GurTIFY1、GurTIFY4、GurTIFY11、GurTIFY13四個基因進行qRT-PCR 驗證,結果顯示(圖7),在相同組織中四個基因的qRT-PCR 相對表達趨勢與轉錄組結果相似,相同基因在不同組織間的qRT-PCR表達差異與轉錄組分析結果相一致。

圖6 烏拉爾甘草不同組織中GurTIFY基因的差異表達分析

圖7 qRT-PCR驗證烏拉爾甘草GurTIFY基因表達
4 討論
烏拉爾甘草是重要的中藥材,有著豐富的藥用功能,但跟其他中藥材一樣,烏拉爾甘草種植區域仍有許多制約因素,如鹽害、干旱和堿性土壤等,嚴重影響烏拉爾甘草的產量和質量,此外,由于全球變暖和適宜種植的土地面積減少,迫切需要進行甘草抗逆品種選育,并改善甘草育種的重要性狀和品質[25],因此,有必要對甘草的發育及其對環境因素的響應進行深入研究。TIFY 家族基因是近年發現的能增強植物抗逆性的新基因。TIFY 基因參與調控多種次生代謝產物的生物合成,但在中藥植物中報道較少。
本研究在全基因組的水平上共鑒定出18 個烏拉爾甘草TIFY 基因,全部分布于細胞核中。基于CCT、Jas、GATA、TIFY4 種保守結構域,將烏拉爾甘草的TIFY 基因家族劃分為TIFY、JAZ、PPD、ZML 四個亞家族。其中JAZ 亞家族基因占比最多。近年來,人們對JAZ 蛋白進行了深入研究,主要是因為它們在植物發育和針對生物和非生物脅迫的防御反應等許多方面起到了作用。茉莉酸作為重要的植物激素,在植物生長、繁殖和抗逆等諸多方面起重要的作用。茉莉酸途徑的抑制因子JAZ(Jasmonate ZIM-domain, JAZ)蛋白(JAZs)是調節茉莉酸(JA)激素應答的關鍵因子,在茉莉酸(JA)信號傳導中,JAZ 蛋白通過與轉錄因子MYC2 結合來調節JA 下游基因的表達,其在廣闊的激素調節網絡中起中心樞紐的作用[26-27]。Li[22]在實驗中發現丹參TIFY家族中屬于JAZ亞家族的SmJAZ3蛋白在JA信號通路中與SmWD40-170相互作用,并在次級代謝產物的積累中發揮重要作用。因此,推測JAZ 基因在烏拉爾甘草生長發育的調控過程發揮了重要的作用,是增強烏拉爾甘草抗逆性的主要基因之一。
順式作用元件分析表明,在烏拉爾甘草的TIFY基因啟動區中光響應元件和激素誘導類元件數量較多,表明該基因與烏拉爾甘草的生長發育密切相關,環境和激素可影響其表達。此外,在烏拉爾甘草TIFY基因啟動子中均預測到大量不同功能的順式作用元件,表明TIFY 基因可能參與了烏拉爾甘草的生長和發育的許多過程。根據差異表達分析,烏拉爾甘草TIFY基因對烏拉爾甘草根和葉器官的生長發育發揮一定的作用,其中GurTIFY15、GurTIFY4、GurTIFY1、GurTIFY11、GurTIFY5、 GurTIFY13、 GurTIFY8、 GurTIFY16、GurTIFY14在烏拉爾甘草葉和根中的表達水平具有顯著差異性,屬于JAZ 亞家族的GurTIFY15、GurTIFY4、GurTIFY1、GurTIFY2和GurTIFY11中葉的表達量較高,而有研究表明擬南芥TIFY 的JAZ 亞家族一些成員基因參與葉片的生長調控[9]。擬南芥PPD 亞家族成員對葉片形態具有調節作用[18],AtTIFY1(ZIM)可以促進葉柄和下胚軸的延伸[17],與赤霉素和油菜素類固醇無關[28]。丹參TIFY 基因家族在不同組織差異表達分析結果得知TIFY 家族整體在葉中的表達較高且其大部分屬于JAZ亞家族[22]。因此,推測GurTIFY在葉的生長發育調控中起到了重要的作用,起調控作用的可能就是JAZ亞家族。
TIFY 基因家族是一個植物特有的基因家族,現已在大部分陸生植物中檢測到了TIFY 基因家族[29-30],目前在一些中藥中也已經有所研究報道,然而該家族在烏拉爾甘草中的表達和功能分析卻很少。對GurTIFY的研究可以為抗逆性遺傳改良提供理論基礎,擴大烏拉爾甘草種植范圍,提高其產量。本研究為進一步探究烏拉爾甘草TIFY 基因家族功能和烏拉爾甘草轉錄調控研究以及利用該基因家族改良烏拉爾甘草抗逆性奠定了基礎。