中藥有效成分對膀胱癌信號通路的調(diào)控作用*
2022-09-29 15:20:00王思宇
王思宇,王 宇
(1. 陜西中醫(yī)藥大學(xué)基礎(chǔ)醫(yī)學(xué)院 咸陽 712046;2. 陜西中醫(yī)藥大學(xué)醫(yī)學(xué)科研實驗中心 咸陽 712046)
膀胱癌是泌尿系中常見的惡性腫瘤之一,在人類最常見腫瘤中排第十位,多發(fā)生于男性患者,其發(fā)病率男女比例為4:1,在男性腫瘤相關(guān)死亡原因中排第九位。膀胱癌的發(fā)病受多種因素影響,其中包括遺傳、環(huán)境、飲食、基因等多種原因。目前,膀胱癌的臨床治療多以手術(shù)、放射治療、系統(tǒng)化療和免疫治療相結(jié)合為主,但預(yù)后仍相對較差[1-3]。近年來,中醫(yī)藥防治腫瘤逐漸引起了人們的廣泛關(guān)注。在中醫(yī)古籍中并未明確提出“膀胱癌”這一病名,但根據(jù)其臨床表現(xiàn)可將其歸屬于“癃閉”“尿血”“溺血”“血淋”等范疇[4]。中藥有效成分在腫瘤的治療上具有毒副作用小、多靶點、多通路的特點。目前,膀胱癌的治療研究需關(guān)注腫瘤中所涉及的每個分子改變,而信號通路的調(diào)控與腫瘤的發(fā)生發(fā)展密切相關(guān),研究表明,中藥有效成分可以通過調(diào)控膀胱癌的多種信號通路,對膀胱癌細胞具有阻滯細胞周期、抑制細胞增殖、誘導(dǎo)細胞凋亡和抑制細胞遷移等多種治療作用。本文根據(jù)已有的實驗研究,就中藥有效成分對膀胱癌信號通路的調(diào)控作用和對膀胱癌組織細胞的影響予以綜述,并展望中藥有效成分在抗膀胱癌治療中的應(yīng)用前景。
1 中藥有效成分對膀胱癌磷脂酰肌醇3 激酶/蛋白激酶B信號通路的調(diào)控作用
1.1 PI3K/AKT信號通路概述
PI3K/AKT 信號通路在腫瘤的發(fā)生發(fā)展中具有至關(guān)重要的作用,是細胞應(yīng)激過程中存在的關(guān)鍵調(diào)節(jié)因子[5]。PI3K 可以被受體酪氨酸激酶(receptor protein tyrosine kinase, RTKs)、G 蛋白偶聯(lián)受體(G Protein-Coupled Receptors,GPCR)或RAS 激活,將4,5-二磷酸磷脂酰肌醇(phosphatidylinositol 4,5-bisphosphate,PIP2)轉(zhuǎn)化為 3, 4, 5- 三磷酸磷脂酰肌醇(phosphatidylinositol 3,4,5-triphosphate, PIP3)。PIP3為3-磷酸肌醇依賴性蛋白激酶1(3-phosphoinositidedependent protein kinase 1, PDK1)和丙酮酸脫氫酶激酶2(pyruvate dehydrogenase kinase 2,PDK2)提供對接位點,從而分別磷酸化AKT 活性中心的Thr308和Ser-473,使AKT 被磷酸化激活。活化的AKT 可以作用于下游信號分子,如哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin, mTOR),進一步調(diào)節(jié)細胞的生長增殖以及遷移[6,7]。PI3K/AKT 信號通路在腫瘤的生長中具有重要意義,針對抑制PI3K/AKT 信號途徑開發(fā)新型抗腫瘤藥物是目前的研究熱點,故可將其作為膀胱癌治療靶點,尋找可以抑制該信號通路的中藥。
1.2 涉及PI3K/AKT信號通路的抗膀胱癌中藥有效成分
黃芩素,又稱黃芩苷元,是從唇形科植物高黃芩根部提取分離出的天然黃酮類化合物,化學(xué)名稱為5,6,7-三羥基黃酮(5,6,7-trihydroxyflavone),其結(jié)構(gòu)式見圖1[8,9]。研究表明,黃芩素對膀胱癌細胞具有抑制作用。王建[10]等以40、80 μmol·L-1的黃芩素處理膀胱癌T24 和5637 細胞24 h 后發(fā)現(xiàn),黃芩素可以抑制膀胱癌細胞增殖,誘導(dǎo)細胞G0/G1期阻滯并促進其凋亡。進一步研究發(fā)現(xiàn),處理后的膀胱癌細胞中Bax/Bcl-2 比值上升,PI3K、p-AKT 及p-mTOR 的表達水平降低。表明黃芩素通過抑制PI3K/AKT/mTOR 信號通路從而對膀胱癌起抑制作用。
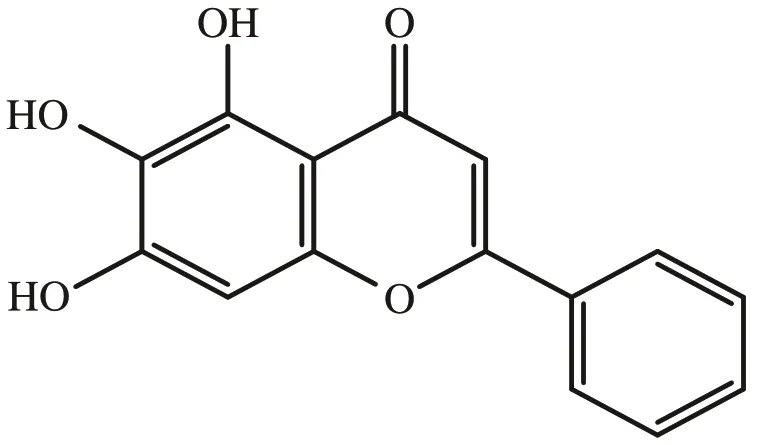
圖1 黃芩素化學(xué)結(jié)構(gòu)圖
姜黃素是從姜黃根莖中提取出來的一種脂溶性多酚類化合物,分子式為C21H20O6,相對分子質(zhì)量368.38,其結(jié)構(gòu)式見圖2[11,12]。研究表明,姜黃素可以通過抑制AKT/PI3K 信號通路降低MMP-2 和MMP-9 的表達,從而抑制T24細胞增殖及遷移[13]。
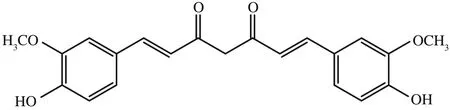
圖2 姜黃素化學(xué)結(jié)構(gòu)圖
此外,其他中藥有效成分同樣可以通過調(diào)控PI3K/AKT 信號通路抑制膀胱癌生長。宋廈[14]等研究發(fā)現(xiàn)200 μmol·L-1的青蒿素可以顯著降低T24 細胞中PI3K 的表達及AKT 和mTOR 的磷酸化水平,通過抑制PI3K/AKT/mTOR 信號通路抑制膀胱癌細胞增殖、遷移并促進其凋亡。吳磊[15]等研究發(fā)現(xiàn)青藤堿對T24 細胞的增殖、遷移及侵襲起抑制作用,并可誘導(dǎo)T24細胞凋亡。此外,以2.0 mmol·L-1的青藤堿處理膀胱癌T24細胞24 h 后發(fā)現(xiàn),青藤堿可以通過抑制PI3K/AKT 信號通路抑制T24細胞發(fā)生EMT。
孫超群[16]等研究發(fā)現(xiàn)氧化苦參堿可以抑制T24 細胞增殖,并促進其凋亡。進一步研究發(fā)現(xiàn),氧化苦參堿可以上調(diào)T24 細胞中PTEN 的表達,下調(diào)PI3K、AKT及mTOR 的表達,通過調(diào)控PTEN/PI3K/AKT/mTOR 信號通路發(fā)揮抗膀胱癌作用。Li T[17]等研究發(fā)現(xiàn)紅景天苷可以誘導(dǎo)T24 細胞自噬,促進p62 和LC3 的表達,抑制PI3K、p-AKT 和MMP-9 的表達,上調(diào)Bax 的表達。這表明紅景天苷通過自噬/PI3K/AKT 和MMP-9 信號通路降低T24細胞活性,誘導(dǎo)其凋亡。
此外,枸杞多糖可以通過抑制PI3K/AKT 信號通路抑制膀胱癌BUI87 細胞的增殖和遷移[18]。毛蕊異黃酮呈時間劑量依賴性抑制膀胱癌細胞的增殖,且對T24 細胞的抑制作用明顯高于T739 細胞。其作用機制主要是通過抑制PI3K/AKT 信號通路活化從而促進Bax 的表達,抑制T24細胞增殖并誘導(dǎo)其凋亡[19]。張智慧[20]以20、40、80 μmol·L-1的蒲公英萜醇刺激膀胱癌T24 細胞48 h 后,發(fā)現(xiàn)其降低細胞中PI3K 和AKT 的磷酸化水平,這表明蒲公英萜醇可以通過抑制活化抑制膀胱癌細胞的增殖,并促進其凋亡。
2 中藥有效成分對膀胱癌核轉(zhuǎn)錄因子-κB 信號通路的調(diào)控作用
2.1 NF-κB信號通路概述
慢性炎癥與腫瘤的發(fā)生具有密切聯(lián)系,NF-κB 是主要的炎癥因子之一,其介導(dǎo)的信號通路可以通過調(diào)節(jié)影響細胞增殖、轉(zhuǎn)移、血管生成和耐藥性基因表達,在腫瘤的發(fā)生和發(fā)展中發(fā)揮作用[21]。NF-κB 是由Rel家族的同源二聚體組成,可以通過調(diào)節(jié)大量功能不同的基因的表達,介導(dǎo)細胞對各種不同刺激的反應(yīng)。NF-κB 主要包括p65,RelB,cRel,p50,p52 五種蛋白質(zhì),通過Rel 同源結(jié)構(gòu)域(Rel homology domain, RHD)進行DNA 結(jié)合和二聚[22]。細胞中主要存在兩種NFκB 激活通路:經(jīng)典NF-κB 信號通路和非經(jīng)典NF-κB信號通路。經(jīng)典通路主要導(dǎo)致IκBα 的磷酸化和p65的異二聚體的核轉(zhuǎn)運,由促炎信號介導(dǎo),依賴于IKKβ和NEMO。而非經(jīng)典通路主要導(dǎo)致p52-RelB 復(fù)合物的生成,由TNF 細胞因子家族的特定成員如CD40 配體、BAFF 和淋巴毒素-β 誘導(dǎo),依賴于IKKα 介導(dǎo)的RelB 相關(guān)的p100 磷酸化[23,24]。NF-κB p50/p65 異源二聚體是腫瘤發(fā)生發(fā)展的重要中介。在膀胱癌中,許多調(diào)節(jié)腫瘤增殖、轉(zhuǎn)移、凋亡和血管生成的相關(guān)蛋白由NF-κB靶基因編碼,并隨著NF-κB信號通路的激活而上調(diào)。研究表明,抑制NF-κB 活化可抑制體外和體內(nèi)膀胱癌細胞的增殖和轉(zhuǎn)移,并促進其凋亡[25,26]。研究發(fā)現(xiàn)中藥有效成分可以通過調(diào)控NF-κB 信號通路對膀胱癌起抑制作用。
2.2 涉及NF-κB信號通路的抗膀胱癌中藥有效成分
隱丹參酮可以抑制膀胱癌J82 細胞增殖并誘導(dǎo)其凋亡。研究者通過Western blot檢測發(fā)現(xiàn),隱丹參酮可以下調(diào)p65 的表達,并上調(diào)Caspase-3 和Caspase-9 的表達。這表明隱丹參酮可以通過抑制NF-κB 信號通路抑制J82 細胞增殖,并誘導(dǎo)其凋亡[27]。Tian F[28]等研究發(fā)現(xiàn)葒草素可以通過抑制NF-κB 進而抑制Hedgehog 信號通路,發(fā)揮對膀胱癌T24 細胞增殖的抑制作用,并促進細胞凋亡。曾文[29]等以20 mg·kg-1的白藜蘆醇灌注膀胱癌大鼠模型14 天后,檢測發(fā)現(xiàn),白藜蘆醇可以下調(diào)Bcl-2 的表達,上調(diào)大鼠NF-κB、RIP2、Caspase-3 的表達,可能通過抑制NF-κB/RIP2 信號通路抑制膀胱癌的生長。
姜黃素除了可以通過調(diào)控AKT/PI3K 信號通路在體外抑制膀胱癌細胞增殖和遷移,還可以通過抑制NF-κB 信號通路在體內(nèi)抑制膀胱癌細胞的增殖。吳金生[30]等研究發(fā)現(xiàn)姜黃素處理后的膀胱癌大鼠,其NF-κB 信號通路的活化被顯著抑制,Bax 的表達升高,Bcl-2 的表達顯著下降,進而抑制N-甲基亞硝基脲誘發(fā)膀胱癌大鼠腫瘤細胞的增殖,并誘導(dǎo)其凋亡。
劉軍[31]等分別以0、5、25、50、100 μmol·L-1的蛇床子素處理膀胱癌T24 細胞48 h 后,EMSA 檢測發(fā)現(xiàn),蛇床子素呈劑量依賴性抑制NF-κB 的活化。此外,蛇床子還可以增加Caspase-3 活性,對膀胱癌細胞起抑制作用。大黃素可以通過下調(diào)NF-κB、XIAP 的表達,并增強膀胱癌細胞中Caspase-3 蛋白表達,從而促進腫瘤細胞凋亡,增強順鉑對T24 細胞移植瘤凋亡的促進作用[32]。張麗瑞[33]等發(fā)現(xiàn)紫鉚因可以抑制T24 細胞的增殖,其作用機制是通過阻斷ERK1/2 和NF-κB 信號通路。曾星[34]等研究發(fā)現(xiàn)豬苓多糖可以通過TLR4 下調(diào)膀胱癌T739 細胞中IKBKB、NF-κB p65、CCL2 及ICAM1的表達,對膀胱癌起抑制作用。
3 中藥有效成分對膀胱癌分泌性糖蛋白/β-鏈蛋白信號通路的調(diào)控作用
3.1 Wnt/β-catenin信號通路概述
Wnt/β-catenin 信號通路是胚胎發(fā)育、細胞生長、增殖、分化及凋亡等過程的重要通路之一,該通路的異常對腫瘤的發(fā)生發(fā)展具有重要意義。在Wnt/βcatenin 信號通路中,β-catenin 信號可以被降解復(fù)合物阻斷。該降解復(fù)合物的主要由支架蛋白Axin、結(jié)腸腺瘤性息肉病蛋白(adenomatous polyosis coli, APC)、酪蛋白激酶1(casein kinase1,CK1)以及糖原合成酶激酶3(glycogen synthase kinase, GSK3)組成。在沒有Wnt信號刺激的情況下,降解復(fù)合物可以通過有效地驅(qū)動β-catenin 的降解,抑制Wnt/β-catenin 通路的激活。而當(dāng)細胞受到Wnt 信號刺激后,它們結(jié)合并激活由Frizzled 和LRP5/6 蛋白組成的異聚受體復(fù)合物,通過散亂蛋白(dishevelled, Dsh)啟動信號反應(yīng),最終抑制降解復(fù)合物。β-catenin 在細胞質(zhì)中積累后轉(zhuǎn)入細胞核,與TCF/LEF-1 相互作用并將其轉(zhuǎn)化為靶基因的轉(zhuǎn)錄激活因子,調(diào)節(jié)Cylin D1、C-myc、Survivin 和MMP-7等下游靶基因的表達[35,36]。以往研究表明Wnt/βcatenin 信號通路參與膀胱癌的病理過程,能夠促進細胞增殖、遷移和侵襲,并抑制其凋亡[37]。由于Wnt/βcatenin 信號通路在膀胱癌發(fā)生發(fā)展過程中發(fā)揮的重要作用,故可以將其可以作為膀胱癌的治療靶點,通過抑制Wnt/β-catenin 信號通路,發(fā)揮藥物對膀胱癌的治療作用。
3.2 涉及Wnt/β-catenin 信號通路的抗膀胱癌中藥有效成分
倪建華[38]等使用10、20、40 mg·L-1的二氫丹參酮處理膀胱癌T24細胞24 h后,PCR 檢測發(fā)現(xiàn),處理后的T24 細胞中CyclinD1 mRNA 表達降低,GSK3β、βcatenin、E-cadherin mRNA 表達增加。這表明二氫丹參酮可能通過GSK3β/Wnt 信號通路對T24 細胞增殖和侵襲起抑制作用,并阻滯細胞周期于S 期。漢防己堿可抑制β-catenin 正向調(diào)控分子DVL 的表達,并增加β-catenin 負向調(diào)控分子GSK3β 的活性增加,從而導(dǎo)致β-catenin 蛋白水平降低,通過抑制Wnt/βcatenin 信號通路,發(fā)揮抗膀胱癌作用[39]。李家仁[40]等通過研究漢防己丙素對亞硝基胺致大鼠膀胱癌的作用,觀察其膀胱癌組織發(fā)現(xiàn)漢防己丙素可以通過上調(diào)大鼠膀胱癌組織中KLF4 的表達,下調(diào)CyclinD1 的表達,上調(diào)p21 的表達,從而抑制膀胱癌細胞增殖。同時,通過抑制KLF4/Wnt4/β-catenin信號通路抑制膀胱癌細胞上皮細胞-間充質(zhì)轉(zhuǎn)化進程,從而抑制膀胱癌細胞的遷移和侵襲。
4 中藥有效成分對膀胱癌絲裂原活化蛋白激酶信號通路的調(diào)控作用
4.1 MAPK信號通路概述
MAPK 家族主要包括四個蛋白成員:細胞外信號調(diào)節(jié)激酶1/2(extracellular regulated protein kinases,ERK1/2)、細胞外信號調(diào)節(jié)激酶5(extracellular regulated protein kinases, ERK5)、c-Jun 氮基末端激酶(c-Jun N-terminal kinase, JNK)、以及p38 絲裂原活化蛋白激酶(p38 mitogen-activated protein kinases,p38)[41]。MAPK 信號通路在腫瘤細胞增殖、遷移、侵襲、血管生成、轉(zhuǎn)移和凋亡等過程中發(fā)揮重要作用[42]。研究表明,在膀胱癌細胞中p38 高表達是膀胱癌不良預(yù)后指標(biāo),與膀胱癌高分期及遠處轉(zhuǎn)移相關(guān)。而抑制ERK1/2 和p38 MAPK 可以誘導(dǎo)膀胱癌細胞周期阻滯,抑制其生長和增殖[43,44]。故尋找有效的MAPK 通路抑制劑對于腫瘤的治療具有重要意義。
4.2 涉及MAPK信號通路的抗膀胱癌中藥有效成分
姚澤欽[45]等給予膀胱癌小鼠模型腹腔注射15 mg·kg-1的丹參酮ⅡA 2 周后,發(fā)現(xiàn)小鼠組織中Cleaved PARP、Bax顯著升高,Caspase-3、Bcl-2明顯降低,同時p-p38 MAPK、p-ERK顯著降低。這表明丹參酮ⅡA可以通過抑制p38 MAPK/ERK信號通路抑制腫瘤增殖并調(diào)控凋亡相關(guān)因子。
此外,和厚樸酚可以通過促進細胞產(chǎn)生ROS,激活MAPK 通路,上調(diào)p38、ERK、JNK 的表達,從而誘導(dǎo)BIU87 細胞凋亡[46]。苦豆堿可以下調(diào)Bcl-2 的表達水平,上調(diào)Bax 的表達水平,抑制ERK1/2 的磷酸化促進膀胱癌EJ細胞凋亡[47]。伍連春[48]等研究發(fā)現(xiàn)黃芩素聯(lián)合U0126 可以通過抑制膀胱癌T24 細胞增殖,并誘導(dǎo)其凋亡。進一步探尋其作用機制,二者聯(lián)合應(yīng)用可以更加顯著的下調(diào)Bcl-2 的表達,上調(diào)Bax 的表達,通過調(diào)控ERK1/2 及p38 信號通路發(fā)揮對T24 細胞的抑制作用。此外,芒柄花黃素可以通過下調(diào)Bcl-2 的表達和活化p38,誘導(dǎo)膀胱癌T24和BIU87細胞凋亡[49]。
TAK1是MAPKKK 蛋白家族成員,其活化后,可以通過激活MKK4/7 活化JNK,進一步調(diào)控細胞凋亡。槲皮黃酮可以通過抑制TAK1/JNK 信號通路,下調(diào)Bal-2、Bal-xL 的表達,從而誘導(dǎo)膀胱癌BIU87 細胞凋亡[50]。
5 中藥有效成分對膀胱癌Janus 激酶/信號轉(zhuǎn)導(dǎo)和轉(zhuǎn)錄激活因子信號通路的調(diào)控作用
5.1 JAK/STAT信號通路概述
JAK/STAT 信號通路在細胞增殖、分化及凋亡等多種生物學(xué)過程中發(fā)揮作用,在多種惡性腫瘤中被激活。JAK/STAT 信號通路主要由三個部分組成:酪氨酸激酶相關(guān)受體、JAK 和STAT。其中JAK 家族是由JAK1、JAK2、JAK3 及酪氨酸激酶(tyrosine kinase 2,TYK2)四個成員組成,可選擇性的與不同的受體鏈相結(jié) 合。STAT 家 族 主 要 由STAT1、STAT2、STAT3、STAT4、STAT5A、STAT5B 和STAT6 七個家族成員組成,STAT3 和STAT5A/B 的激活可以促進腫瘤發(fā)生,而STAT1的激活則具有相反的作用[51,52]。
在腫瘤細胞中,JAK/STAT 信號通路的激活方式有多種,其中包括自分泌/旁分泌細胞因子的產(chǎn)生,激活受體、JAK 或其他上游致癌基因的突變,進而激活STATs,以及激活STATs 本身的突變[53]。有研究表明,JAK/STAT 信號通路的活性與膀胱癌細胞的遷移和侵襲能力呈正相關(guān)[54]。此外,JAK/STAT 信號通路在維持膀胱癌干細胞的干性、自我更新以及增殖潛能方面也發(fā)揮著重要作用[55]。因此,抑制JAK/STAT信號通路在抑制膀胱癌發(fā)生發(fā)展過程中發(fā)揮著重要作用。
5.2 涉及JAK/STAT信號通路的抗膀胱癌中藥有效成分
于汝通[54]等使用5、10、25、50 μmol·L-1的黃芩素處理膀胱癌T24 細胞48 h 后,通過Tanswell 檢測發(fā)現(xiàn),黃芩素呈濃度依賴性抑制T24細胞的遷移和侵襲。其中50 μmol·L-1黃芩素處理的T24細胞,可以下調(diào)uPA、MMP-9的表達,上調(diào)TIMP-1、TIMP-2的表達,同時p-JAK2、p-STAT3 的表達顯著降低。而JAK/STAT 激活劑逆轉(zhuǎn)了黃芩素對T24 細胞遷移和侵襲的抑制作用。這表明黃芩素通過抑制JAK2/STAT3 的活化發(fā)揮抗膀胱癌作用。此外,白花蛇舌草可能通過抑制JAK2/STAT3 信號通路抑制膀胱癌細胞增殖,誘導(dǎo)細胞凋亡。并且呈濃度依賴性抑制T24 細胞Bcl-2 的表達,上調(diào)Bax、Caspase-3的表達[56]。
6 中藥有效成分對膀胱癌其他信號通路的調(diào)控作用
此外,中藥有效成分對其他膀胱癌信號通路具有調(diào)控作用。黃晨[57]等使用濃度為100、200 mg·L-1的三七總皂苷處理膀胱癌T24 細胞48 h 后,發(fā)現(xiàn)三七總皂苷可以下調(diào)抗凋亡蛋白Bcl-2 的表達,上調(diào)cleaved Caspase-3 的表達,同時下調(diào)Hippo-YAP 信號通路相關(guān)蛋白YAP1 和TEAD1 的表達,上調(diào)LATS1 的表達。這表明三七總皂苷可以通過激活Hippo-YAP 信號通路抑制膀胱癌T24 細胞增殖、降低其遷移和侵襲能力并促進細胞凋亡。倪建華[58]等研究發(fā)現(xiàn)鼠尾草酚可以通過上調(diào)PPARγ mRNA 及蛋白的表達,激活PPARγ信號通路,進而下調(diào)β-catenin mRNA 和蛋白的表達,以此抑制膀胱癌T24 細胞增殖、降低其遷移和侵襲能力并促進細胞凋亡。中藥有效成分對膀胱癌信號通路的調(diào)控作用的總結(jié)如下(表1)。
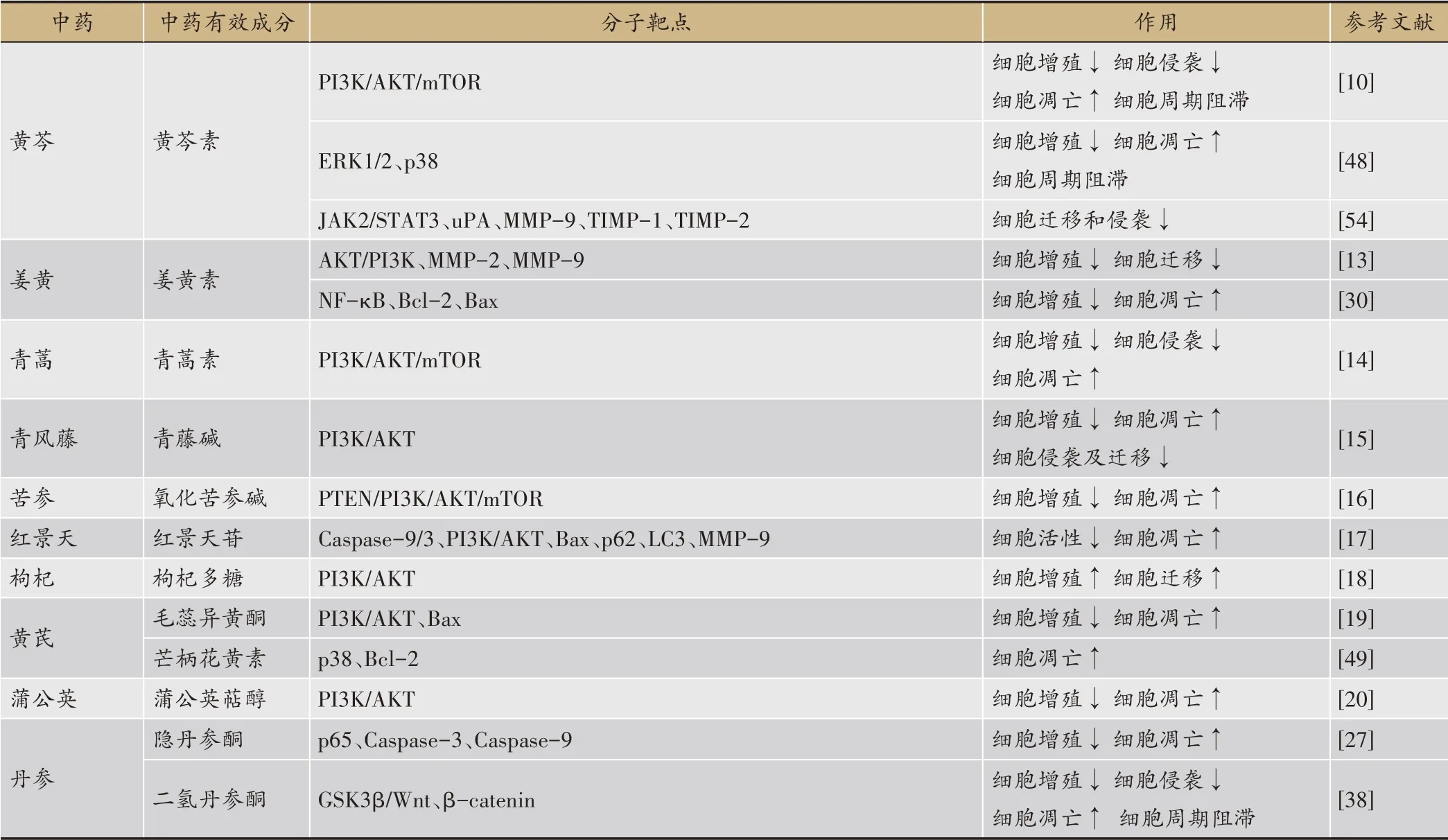
表1 中藥有效成分對膀胱癌信號通路的調(diào)控作用

續(xù)表
7 問題與展望
綜上所述,中藥有效成分可以通過調(diào)控PI3K/AKT、NF-κB、Wnt/β-catenin、MAPK 和JAK/STAT 等多種信號通路,激活或抑制其中關(guān)鍵因子,進而抑制膀胱癌細胞增殖、遷移和侵襲、阻滯細胞周期并誘導(dǎo)腫瘤細胞凋亡,發(fā)揮治療作用。
雖然已有大量研究指出中藥有效成分對膀胱癌的治療作用,但仍存在一些問題。首先,目前對于中藥有效成分抗膀胱癌作用機制的研究多針對單一的信號通路,而本文總結(jié)發(fā)現(xiàn)部分中藥有效成分可以調(diào)控多種信號通路,如黃芩素可以通過調(diào)控PI3K/AKT、MAPK、JAK2/STAT3 等多種信號通路對膀胱癌細胞發(fā)揮抑制作用[10,48,54]。此外,不同信號通路間存在串?dāng)_影響。但相關(guān)研究相對較少。其次,根據(jù)已有的研究,Notch[59]、TGF-β/Smads[60]、HIF-1α/VEGF[61]、PD-1/PD-L1[62]等信號通路與膀胱癌的發(fā)生發(fā)展具有密切聯(lián)系,但缺少中藥有效成分通過調(diào)控此類信號通路發(fā)揮抗膀胱癌作用的研究。此外,臨床上的腫瘤治療方案以多種抗癌藥物聯(lián)合使用為主,而多種中藥有效成分或中藥有效成分與化療藥物聯(lián)合應(yīng)用能否增強治療效果。在今后的研究中,更多有關(guān)中藥有效成分治療膀胱癌的研究有待展開,研究內(nèi)容可以是同一中藥有效成分針對多種分子途徑的機制研究,或多種中藥有效成分對同一信號通路調(diào)控作用的研究。中藥復(fù)方中最常用于治療膀胱癌的中藥有半枝蓮、薏苡仁、白花蛇舌草、茯苓、白術(shù)、生地黃、澤瀉、黃柏、豬苓、黨參、黃芪、甘草等[63],可以就以上藥物的有效成分為切入點,展開新的研究,明確其抑制膀胱癌的作用機制。中藥治療膀胱癌的作用機制的闡明,對今后其應(yīng)用于臨床,提高膀胱癌患者生活質(zhì)量具有重要意義。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年4期)2021-12-01 11:19:40
中老年保健(2021年4期)2021-08-22 07:08:32
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
金橋(2020年7期)2020-08-13 03:07:00
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
基層中醫(yī)藥(2018年6期)2018-08-29 01:20:20