黃連NRAMP5基因的克隆與生物信息學分析*
2022-09-29 15:20:14王文斌朱海蘭胡志剛
世界科學技術-中醫藥現代化 2022年5期
王文斌,朱海蘭,莫 靜,劉 迪,胡志剛**,汪 波
(1. 湖北中醫藥大學藥學院 武漢 430065;2. 湖北省藥品監督檢驗研究院 武漢 430075;3. 湖北大學生命科學學院 武漢 430062)
1 引言
從廣義上說,重金屬是指金屬密度大于5g/cm3的金屬[1]。按照這個定義,有53 種天然存在的金屬屬于重金屬。重金屬在高濃度下對植物均有毒害,但其中一些,例如鋅(Zn)、鐵(Fe)和銅(Cu)等,在低濃度時是必要的營養素,而其他一些,例如鎘(Cd)、鉻(Cr)和鉛(Pb),即使在極低的濃度下也會對植物造成危害。那些在低濃度下對植物表現出毒性的重金屬通常被稱為重金屬[2]。鎘被認為是對人體健康最危險的重金屬元素之一。人體攝入鎘過量會導致以腎小管病變為主的腎臟損害、以肺炎和肺水腫等為代表的呼吸系統損傷,造成骨骼疏松、萎縮、甚至斷裂等,嚴重危害人類健康。鎘不是植物生長的必需元素[3],對植物也有很強的毒害作用,鎘大量積累會影響植物的生長發育,導致植物葉片黃化枯敗、株高降低等現象,甚至使植物死亡[4-5]。植物在進化過程中,已經發展出了獨特的方法來獲取必需的營養元素,抑制或富集土壤中的有毒金屬元素[6]。對重金屬在植物中累積途徑的深入了解將對植物生長發育研究和人類健康具有重要意義,而金屬轉運蛋白的克隆則是關鍵的第一步。
在植物吸收金屬后,金屬離子與載體蛋白結合形成螯合物進入到細胞內[7-8]。植物重金屬轉運蛋白包括鋅鐵轉運蛋白、重金屬ATP 酶、自然抗性相關巨噬細胞蛋白等,其中大量研究表明自然抗性相關巨噬細胞蛋白可以特異性轉運鎘和錳[9]。NRAMP 家族蛋白是一類高度保守膜蛋白,在細菌、真菌、植物和動物體內都廣泛存在。NRAMP 蛋白在不同物種間表現出功能差異和特異性表達。MmNRAMP1 最早在小鼠身上被發現,它主要通過對金屬離子吸收而調節對分枝桿菌的吞噬作用[10]。而植物中普遍存在多個NRAMP家族基因,模式植物擬南芥的NRAMP家族成員有6 個,水稻中有7 個,煙草有9 個。Bozzi 等[11]發現,NRAMP能夠結合或運輸生物所需的二價金屬(Mn2+,Fe2+,Co2+,Ni2+,Cu2+,Zn2+)和有毒金屬(Cd2+,Pb2+,和Hg2+),但不能運輸二價堿土金屬離子(Mg2+和Ca2+)。超積累型東南景天有6個NRAMP家族基因,具有重金屬離子吸收和轉運功能[12-13]。如蘋果MhNRAMP1基因能夠在根部轉運鎘離子,并抑制了抗細胞死亡相關基因的表達[14-15]。大豆GmNRAMP1可能參與鎘離子吸收轉運[16]。萊茵衣藻NRAMP3的表達可能具有減少萊茵衣藻細胞內Cd 的積累的作用[17]。超積累型東南景天受到鎘脅迫時,SaNRAMP6基因葉片表達量下降,根系表達量顯著增加,表明SaNRAMP6可能直接參與鎘的吸收 和 轉 運 過 程[18]。 Cailliatte 等[12]認 為 擬 南 芥AtNRAMP6是一種細胞內金屬轉運體,主要作用于液泡內膜,并能影響鎘在細胞中的分布和可利用性。辣椒NRAMP1和NRAMP3基因主要參與Cd 從根部轉運到地上部的過程,NRAMP2、NRAMP5、NRAMP6基因參與辣椒根對Cd的吸收過程[19]。
黃連為湖北省“一縣一品”的道地藥材,湖北省利川市及周邊地區是黃連的道地產區,在我國傳統中醫藥中具有悠久的使用歷史,最早可追溯到我國已知現存最早的本草著作《神農本草經》。黃連作為我國著名的傳統中藥,不僅在臨床上大量使用,還出口到國外,使其需求量大大增加。但是,黃連對生長環境要求嚴格,生長期長,導致黃連市場供需矛盾加劇,野生黃連被過度采挖逐漸變成三級瀕危植物,現市場上使用的主要是人工種植黃連。目前栽培黃連主產地為重慶石柱縣、湖北利川市,以及四川、湖南等地也有種植。而近年來黃連藥材出口檢測中發現部分重金屬超標,特別是鎘超標,嚴重影響黃連產業發展[20]。對擬南芥、水稻等模式植物的研究結果表明,NRAMP 蛋白家族在植物對鎘的吸收和轉運中起著重要作用。
2 研究思路與方法
本文旨在克隆分析黃連NRAMP基因特性,探討黃連Cd 吸收轉運分子機制。以黃連基因組[21]測序結果為基礎,通過比對篩選出與擬南芥NRAMP基因同源性較高的黃連NRAMP5基因,通過PCR 技術克隆該基因的開放讀碼框,并通過生物信息學分析黃連NRAMP5蛋白的理化性質、亞細胞定位、跨膜結構域、系統進化關系等(圖1),對基因功能以及編碼蛋白的性質作出初步的判斷和預測,為研究黃連體內重金屬鎘轉運機制以及基因的功能作用提供參考依據,以期為后續的CcNRAMP5基因表達分析和功能驗證研究提供基礎。
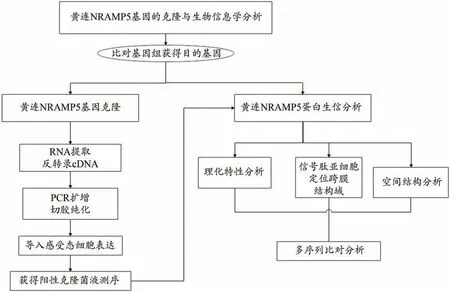
圖1 技術路線圖
3 研究與分析
3.1 材料、試劑與儀器
3.1.1 材料
黃連植株采自湖北省神農架林區,種植于湖北省藥品監督檢驗研究院分子生物實驗室,經DNA條形碼鑒定為毛茛科植物味連(Coptis chinensis)。實驗材料系實驗室水培培養3周長勢良好的黃連葉片。
3.1.2 試劑
pEASY?-Blunt 克隆載體和Trans1-T1 大腸桿菌感受態細胞購自北京全式金生物科技有限公司。HLingene 瓊脂糖凝膠回收/PCR 產物純化試劑盒和Thermo Fisher cDNA Synthesis Kit 反轉錄試劑盒購于上海惠凌生物技術有限公司。IPTG 溶液(50 mg·mL-1)、X-Gal 溶液(20 mg·mL-1)、Amp 溶液(100 mg·mL-1)購自索萊寶生物科技有限公司。高效率PCR 酶KOD Dash 購于東洋紡(上海)生物科技有限公司。植物總RNA 提取試劑盒購于天根生化科技(北京)有限公司。引物合成及測序由武漢天一華煜基因科技有限公司完成。
3.1.3 儀器
冷凍高速離心機(Thermo Fisher Scientific);Nano drop-2000 超微量分光光度計(Thermo Fisher Scientific);T100 型PCR 擴增儀(美 國Bio-Rad);GGI54TW 蒸汽消毒鍋(美國ZEALWAY 公司);BCD-225TMPM 冰箱(海爾);MX-RL-E 旋轉混懸儀(北京大龍興創);DYY-10C 電泳儀(北京六一);Gel Doc XR+凝膠成像系統(美國Bio-Rad 公司);LWB-26 雙列六孔電熱恒溫水浴鍋(上海龍躍);UF55 干燥箱(德國Memmert 美墨爾特);2.5μL、10μL、20μL、100μL、200μL、1000μL 移液器(eppendorf,德國艾本德股份公司)等分子生物實驗室常規儀器等。
3.2 方法
3.2.1 目的基因的獲得
以擬南芥、水稻、東南景天等有文獻報道的NRAMP 蛋白序列為參考序列,比對黃連基因組,通過blastp比對尋找黃連NRAMP家族基因(e值≤e-10),并且將它們在Pfam 蛋白質家族數據庫(http://pfam.xfam.org/search)中進行檢驗以確保其包含NRAMP 保守結構域(PF01566),得到黃連NRAMP5基因。
3.2.2 總RNA的提取、檢測及cDNA的合成
黃連葉片用清水沖洗洗凈,濾紙吸干,并用液氮研磨成粉末,使用天根植物總RNA提取試劑盒提取總RNA。使用瓊脂糖凝膠電泳和超微量分光光度計檢測提取的RNA 質量和濃度。然后以提取的總RNA 為模板,采用試劑盒反轉錄合成cDNA,50℃水浴20 min,85℃水浴10 min,滅活逆轉錄酶,儲存在-20℃。
3.2.3 黃連NRAMP5基因的克隆
依據已測得的黃連基因組數據,查找該基因上下游2000 bp 長度,分別設計擴增引物(上游引物LF:AAGGAGGAGTGCATCCATTG,LR: CTGCATCTTTGG GTGTCAT;下游引物RF: ATGCAGTGATCTTACTCTC AATTCGGC, RR: TGTGGCAATGTGTGATTGTG)將目的序列片段擴增,以確定黃連NRAMP5基因編碼序列。根據獲得的黃連NRAMP5基因序列,設計特異性擴增引物NRAMP-5F:5’-ATGGAAAGTGAAAATAC CCAAAAGG-3’,NRAMP-5R:5’-CTATTTATTCTGA TGTGTCATTTCAGC-3’。以反轉錄得到的黃連葉片cDNA 為模板進行PCR 擴增,獲得NRAMP5的完整編碼序列。PCR 反應體系為:1μL KOD Dash,5μL 10 ×Buffer for KOD Dash,5μL 2 mM dNTPs,3μL cDNA,34μL ddH2O,1μL Forward Primer(10μmol·L-1),1μL Reverse Primer(10μmol·L-1),共50μL。PCR 反應程序為:95℃5 min;95℃30 s,62℃30 s,74℃1 min 30 s,反應進行32個循環;74℃10 min。
PCR 產物經切膠回收后,純化產物與pEASYBlunt 克隆載體連接并轉化至Trans1-T1 感受態細胞,隨機挑選陽性克隆經菌液PCR 驗證長度無誤后送武漢天一華煜基因科技有限公司測序。
3.2.4 黃連NRAMP5基因的生物信息學分析
序列組裝完成后,利用TBtools 軟件[22]查找黃連NRAMP5基因的開放閱讀框并翻譯成氨基酸序列,在NCBI 數據庫中對氨基酸序列進行Blastp 比對;利用ExPASy 在線軟件分析NRAMP5編碼氨基酸序列的組成和理化性質;利用ProtScale 在線工具進行蛋白疏水性分析;利用SignalP 進行蛋白信號肽預測;利用ProtComp 在線軟件對NRAMP5的氨基酸序列進行亞細胞定位;利用TMHMM 通過隱馬爾可夫模型來預測蛋白跨膜區域判斷其是否為跨膜蛋白;利用SPOMA分析NRAMP5的蛋白質二級結構并進行預測,以了解局部空間結構;利用SWISS-MODEL 預測CcNRAMP5蛋白的三級結構;使用MEGA 6.05 軟件對氨基酸序列進行多重比對,采用Neighbor-Joining 法構建蛋白質系統發育樹,Bootstrap 統計檢驗次數為500,其他參數設置為默認參數。
3.3 結果
3.3.1 黃連RNA的提取和黃連NRAMP5基因的克隆
根據試劑盒說明書,從黃連葉片中提取總RNA,選取凝膠電泳條帶清晰、吸光度與濃度均滿足實驗要求的RNA 作為模板RNA。用反轉錄試劑盒將RNA 樣品合成cDNA第一條鏈。
以黃連葉片cDNA 為模板,針對NRAMP5基因設計的引物擴增了約1500 bp 的PCR 產物,與預期大小一致。將該片段切膠回收后與pEASY- Blunt 載體連接,轉化至大腸桿菌感受態細胞,PCR 鑒定為陽性克隆的菌液進行測序,最后將序列提交至GenBank,基因登錄號為KAF9609000.1。
用CodonCode Aligner 軟件拼接測序結果的峰圖,得到1611 bp 的cDNA 序列,經TBtools 軟件分析發現其為一個完整的開放閱讀框,共編碼536 個氨基酸。通過NCBI blastp 將該氨基酸序列與NR 數據庫進行比對,結果表明,與GenBank 已報道的其他植物的NRAMP5基因具有較高的相似性,將該基因命名為CcNRAMP5。利用Pfam 分析氨基酸序列的保守結構域,發現CcNRAMP5 蛋白的第63-425 位氨基酸殘基是典型的NRAMP結構域(PF01566),同時在第279-282 氨基酸殘基處有一個N-糖基化位點(Nglycosylation site),屬于NRAMP超家族。
3.3.2CcNRAMP5基因編碼蛋白的序列分析
(1) 理化特性分析
利用ExPASy 數據庫的ProtParam 工具分析CcNRAMP5基因轉化的蛋白序列,發現該蛋白質的分子量為58.19 kD,等電點為7.62,帶負電荷的殘基(Asp+Glu)為37,帶正電荷的殘基(Arg+Lys)為38,含有20種氨基酸,亮氨酸含量最高,占比14%,色氨酸含量最低,占比1.3%,分子式為C2683H4257N667O729S21。蛋白質不穩定系數為33.7,屬于穩定蛋白質。
使用EXPASY 的Protscale 在線工具預測NRAMP5蛋白的親水性和疏水性(圖2),發現在第35 個氨基酸位置,最低峰值是-3.022,在第184 個氨基酸位置,最高峰值是3.711,平均親水性為0.574,這表明該蛋白是疏水性蛋白。

圖2 CcNRAMP5蛋白的親疏水性預測結果
(2) CcNRAMP5 蛋白信號肽、亞細胞定位、跨膜結構域預測
SignalP-5.0 Server軟件分析結果顯示,含有Sec信號肽(Sec/SPI)的可能性為0.0016,不含有信號肽(Other)的可能性為0.9984,預測CcNRAMP5蛋白不含信號肽序列,不屬于分泌型蛋白。使用ProtComp 9.0 對CcNRAMP5的亞細胞定位進行預測,預測結果表明CcNRAMP5 蛋白質定位于質膜。利用TMHMM Server v. 2.0 預測CcNRAMP5 蛋白的跨膜結構域(transmembrane domain,TMD),對應的氨基酸殘基區域分別為78-110、121-143、147-169、178-200、220-242、263-285、323-345、365-387、391-413、426-448、463-485。結果表明,CcNRAMP5 蛋白含有11 個跨膜區域,具有膜蛋白的重要特征(圖3)。
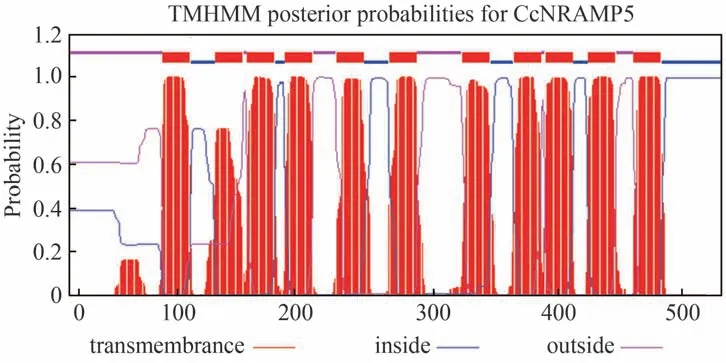
圖3 黃連NRAMP5蛋白跨膜結構域分析
(3) CcNRAMP5蛋白二級結構預測
經PRABI 網站的SOPMA 程序對CcNRAMP5 蛋白二級結構進行分析,預測發現CcNRAMP5 蛋白主要由α 螺旋(a helix)、延伸鏈(Extended strand)、β 轉角(Beta turn)和無規則卷曲(Random coi)四部分結構組成,分別占比47.57%、13.99%、2.99%和35.45%。α-螺旋與無規則卷曲是多肽鏈的主要結構元件。
(4) 三維建模
三級結構預測分析顯示(圖4),CcNRAMP5 蛋白與葡萄球菌錳轉運突變體MntH 的晶體結構(二價金屬陽離子轉運體MntH)具有37%的序列相似性,以該蛋白(SMTL ID:5m8k.1.A)為模板,通過同源建模構建CcNRAMP5 蛋白的三級結構,用于建模的氨基酸殘基范圍為36-493位,占編碼氨基酸總數的85.26%。
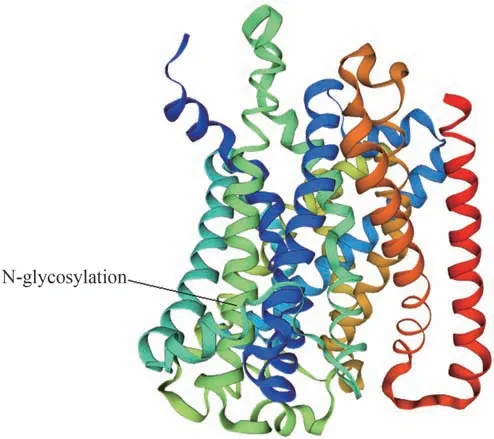
圖4 黃連NRAMP5蛋白的三級結構
3.3.3 多序列比對分析
為研究黃連CcNRAMP5 蛋白在物種中的進化位置和親緣關系,利用MEGA 6.05對水稻(Oryza sativa)、擬南芥(Arabidopsis thaliana)、歐洲油菜(Brassica napus)、波蘭小麥(Triticum polonicum)、可可(Theobroma cacao)、大麥(Hordeum vulgare)等植物的NRAMP5蛋白序列進行進化樹分析比較(圖5)。結果表明,黃連CcNRAMP5與同為毛茛目的博落回McNRAMP5(Macleaya cordata, accession no.OVA12788.)、罌 粟PsNRAMP5(Papaver somniferum,accession no. XP_026445592.)親緣關系最近,與桑(Morus alba)、蘋果(Malus domestica)、可可(Theobromacacao)、香橙(Citrus junos)、甜橙(Citrus sinensis)等聚為一個大類。水稻與大麥、小麥聚為一支,可能與其均為單子葉植物相關。
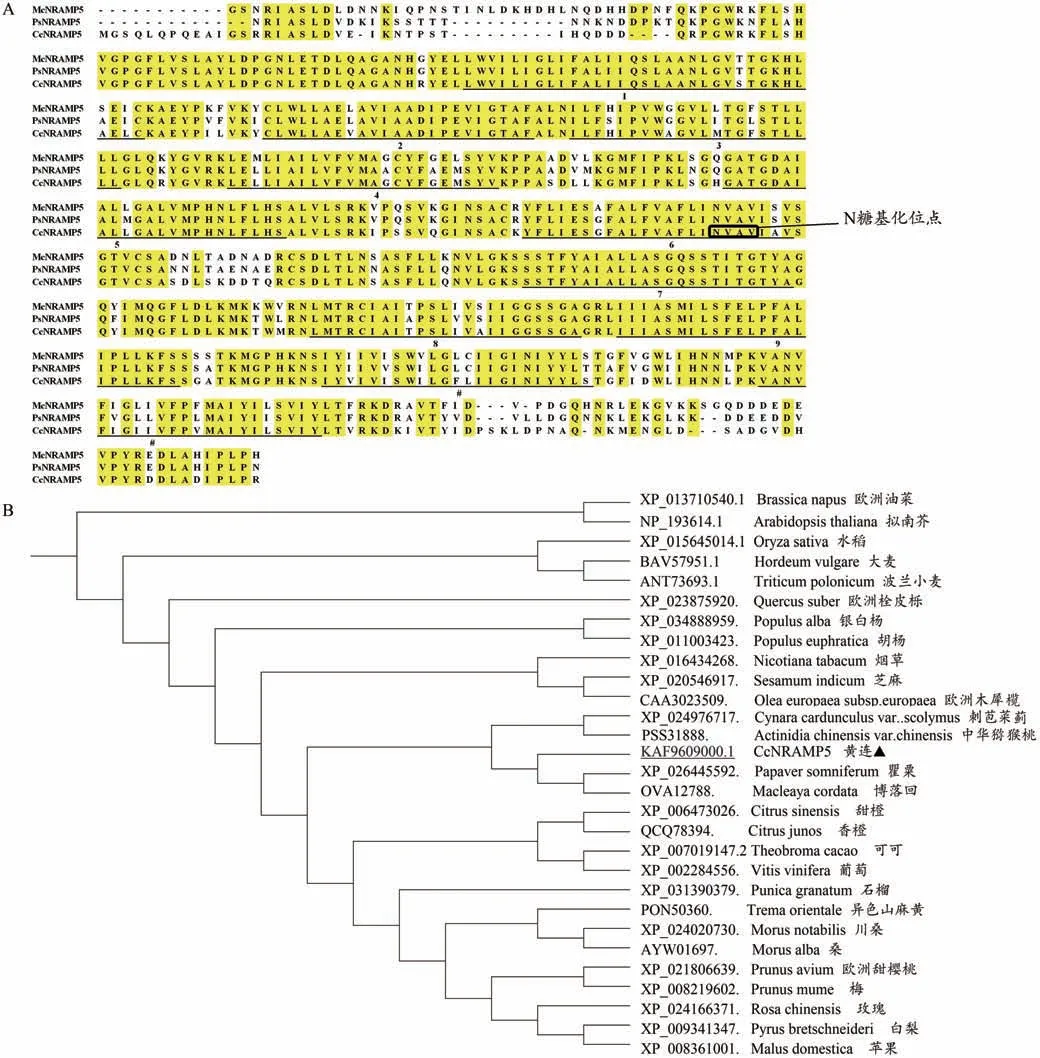
圖5 黃連NRAMP5蛋白的多序列比對分析
4 討論
黃連屬于毛茛科植物,是現存雙子葉植物的重要基部類群之一,其根莖作為中藥使用已有2000多年的歷史,不但是中醫處方最常用品種,也是中成藥的重要原料。據《全國中成藥品種目錄》統計,以黃連作原料制生產的中成藥有黃連上清片、香連丸等108 種之多[23]。黃文麗等人[24]利用ICP-MS、細胞分餾、連續萃取等方法分析鎘在黃連中的分布及化學形態,發現黃連中Cd 的累積分布為:根>莖>葉>子實,大部分富集在根部,少量遷移到地上部,且Cd主要積累在周皮、皮層、髓和根際維管束中,外周皮和皮層中Cd含量較高,髓和根際維管束中Cd含量較低,同時還發現在黃連中Ca 與Cd 的時空分布位置相似,認為Ca2+通道是Cd 進入植物的一條途徑,對二價陽離子轉運蛋白的挖掘研究對解析其重金屬鎘富集機制,對推動其無公害種植具有重要的現實意義。
NRAMP 家族基因在進化過程中高度保守,廣泛分布于哺乳動物、酵母和高等植物中,并參與了金屬轉運和抗微生物感染等多種過程。到目前為止,該家族的幾個成員已經從擬南芥、水稻和番茄中分離出來。在本研究中,從黃連中克隆獲得可能參與鎘吸收轉運的NRAMP基因CcNRAMP5,并以此為基礎對其基本理化性質、亞細胞定位以及結構組成等方面進行了分析。CcNRAMP5 蛋白具有11 個跨膜區,在第279-282 位氨基酸殘基處存在一個N-糖基化位點,含有一個NRAMP 特征結構域,亞細胞定位在質膜上,這符合NRAMP 蛋白的重要特征。序列相似性比對和系統進化樹分析表明,黃連CcNRAMP5 蛋白與博落回、罌粟等植物的NRAMP5 蛋白親緣關系最近,序列相似性高,同時推測CcNRAMP5 蛋白可能與這些蛋白具有相似功能。
在水稻中,OsNRAMP5表達于質膜中,參與水稻錳、鐵、鎘的轉運,在水稻根表皮、外皮層、皮層外層以及木質部周圍組織中均有表達[25-27]。大麥HvNramp5定位在質膜上,是鎘和錳的金屬轉運體[28]。擬南芥AtNRAMP3和AtNRAMP5位于液泡膜上,在擬南芥的根和葉中均有表達,作為鎘轉運蛋白,他們參與鎘離子向細胞質的外排,起重金屬解毒作用[29]。波蘭小麥TpNRAMP5可以運輸Cd、Co、Mn,但對鐵和鋅含量沒有影響[30]。煙草中NRAMP基因在生物脅迫響應和次生代謝途徑中具有特異性。可可中TcNRAMP3和TcNRAMP5可以轉運生長必需的二價金屬陽離子(Fe、Mn)和Cd2+,敲除TcNRAMP5有望獲得低Cd 品種可可[31]。這說明NRAMP家族基因在維持植物體對金屬的吸收和運輸中起重要作用,尤其是對二價金屬離子,如Fe、Mn、Zn、Cu 和Cd。CcNRAMP5 是第一個從黃連植物中分離出來的NRAMP 金屬轉運蛋白家族成員,與桑(Morus alba)[32]、蘋果(Malus domestica)[15]、可可(Theobroma cacao)、香橙(Citrus junos)[33]親緣關系較近,可能與黃連植物體內包括Cd在內的二價金屬離子的吸收和轉運有關,其機制和功能在進一步的功能驗證工作中將進行探討。
5 結論
綜上所述,本次研究首次通過TA 克隆得到黃連NRAMP5基因,并運用生物信息學方法對其理化性質、結構特征等進行了分析預測,綜合前人研究結果,筆者推測黃連NRAMP5基因可能在黃連Cd 的吸收轉運過程發揮重要作用。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44