兩種生態型香根草對鎘的耐受和積累特性比較
2022-10-01 15:22:28董名揚馮曉暉
西北植物學報 2022年8期
董名揚,孫 瑤,馮曉暉,汪 毅,胡 斌,周 強,2*
(1 吉首大學植物資源保護與利用湖南省高校重點實驗室,湖南吉首 416000;2 錳鋅礦業重金屬污染綜合防治技術湖南省工程實驗室,湖南吉首 416000)
隨著人類生產活動和社會發展,大量的重金屬被排放到環境中,導致土壤污染嚴重。鎘是對人體毒性最大的重金屬之一,其在《全國土壤污染調查公報》的重金屬污染元素中排名最高[1],中國約有7%的土壤存在不同程度的鎘污染。在《中國耕地地球化學報告》中,中國耕地重金屬超標位點占比為8.2%,其中多數為鎘、鎳等有毒元素復合污染[2]。土壤中的鎘被作物吸收并積累在體內,會影響植物的光合作用、新陳代謝、生長發育等過程,并降低其產量和品質。作物體內的鎘還能通過食物鏈等途徑進入人體,影響人類健康[3]。因此,土壤鎘污染治理刻不容緩。植物修復在治理鎘等重金屬污染領域有著廣闊的前景,與傳統的物理和化學修復技術相比,其具有處理費用低、對環境干擾小、不會產生二次污染等優勢[4]。比較不同生態型植物對重金屬的積累特點,可以篩選修復效果更佳的種質資源和揭示其積累機制,這是目前研究的熱點之一。有研究表明不同生態型東南景天(SedumalfrediiHance)[5]、天竺葵(PelargoniumhortorumBailey)[6]和多花黑麥(LoliummultiflorumLam.)[7]對鎘的耐受和積累存在明顯的差異,為其應用于鎘污染治理提供了充分的科學依據。
香根草[Chrysopogonzizanioides(L.) Roberty]為禾本科多年生草本植物,具有根系發達、生物量大、適應性廣和抗逆性強等特點,對多種重金屬均有一定的富集作用,具有用于植物修復的潛力[8]。香根草作為世界知名的水土保持植物,已選育推廣了數十種生態型。周強等[9]研究發現不同生態型香根草的光合特性存在明顯差異,Monto等4種生態型香根草的光合日變化呈雙峰曲線,而Veriveria nermorlis等2種生態型香根草的光合日變化呈單峰曲線。但不同生態型香根草對鎘的耐受能力和積累特性是否存在差異及其耐受鎘脅迫的機制都尚不明確。因此,本研究比較了Monto和Veriveria nermorlis兩種生態型香根草的鎘耐受和積累特征,旨在為香根草應用于鎘污染土壤的植物修復提供更加堅實的理論依據。
1 材料和方法
1.1 實驗材料
試驗在湖南省吉首市進行,Monto(MT)和Veriveria nermorlis(VN)兩種生態型香根草引種自中國科學院華南植物園,并長年種植在吉首大學的試驗苗圃中。在試驗準備階段,將分蘗繁殖得到的香根草幼苗單株培養,挑選長勢健壯的香根草(苗齡5個月)置于1/2 Hoagland營養液中培養,待其恢復生長后進行鎘脅迫處理。
1.2 試驗設計
將香根草植株固定于塑料盆中,以1/2 Hoagland營養液作為培養液,設置3種濃度的硝酸鎘溶液作為輕度(200 μmol·L-1)、中度(800 μmol·L-1)、重度(2 000 μmol·L-1)鎘脅迫處理,以1/2 Hoagland營養液作為對照(無鎘添加),并將pH調節至5.8。香根草于植物培養箱中培育7 d,每日更換處理溶液。培養期間晝夜溫度分別為(28±2)℃和(22±2)℃;白晝16 h·d-1;光照強度為1 000 μmol·m-2·s-1。每處理設置3次重復。培養結束取樣進行葉片葉綠素熒光參數和葉綠素含量測量后,植株用20 mmol·L-1EDTA-2Na浸泡根部20 min,再用去離子水反復沖洗,擦干表面水分后,分葉片和根系取樣。一部分新鮮樣品用于測定亞細胞組分鎘含量和不同化學形態鎘含量,另一部樣品105 ℃殺青30 min后,70~80 ℃烘干至恒重用于鎘含量的測定。
1.3 測定指標及方法
1.3.1 葉片葉綠素熒光參數香根草植株暗適應30 min后,采用PAM-2500(Walz,德國)進行葉片葉綠素熒光參數測量。其初始熒光(Fo)、最大熒光(Fm)、PSⅡ最大光化學效率(Fv/Fm)數值從儀器直接讀取,PSⅡ潛在活性(Fv/Fo)按照公式(Fm-Fo)/Fo計算[10]。
1.3.2 葉片葉綠素含量使用SPAD-502便攜式葉綠素儀(Konica Minolta Sensing,日本)測量香根草葉片的相對葉綠素含量(SPAD值)。
1.3.3 葉片和根部鎘含量稱取0.1 g香根草葉片或根系的干燥粉末放置于消解管中,并加入HNO3和HClO4(V∶V=4∶1)混合酸液3 mL,靜置6~8 h,然后用石墨消解儀消煮至溶液澄清,冷卻并定容至25 mL,再經0.22 μm微孔濾膜過濾。采用石墨爐原子吸收光譜儀(AA-7050型原子吸收分光光度計)測定鎘含量。據測定結果分別計算富集系數(bioconcentration factor, BF)和轉移系數(translocation factor,TF)[11]。
BF=植株根或葉片中鎘濃度/溶液中鎘濃度
TF=葉片中鎘濃度/根中鎘濃度
1.3.4 葉片和根系亞細胞組分的鎘含量亞細胞組分的分離采用差速離心法[12]。稱取0.2 g植物新鮮樣品剪碎,用預先配置好的Tris-HCl緩沖液(M∶V=1∶10,0.25 mol·L-1蔗糖、Tris-HCl 緩沖液[pH 7.5]和1 mmol·L-1二硫赤蘚糖醇)研磨成勻漿,轉入離心管中,1 000 g離心15 min,得到的沉淀為細胞壁組分;上清液于10 000 g離心30 min,得到的沉淀為細胞器組分;上清液為可溶性組分。所得各亞細胞組分按照1.3.3中的方法消解并測定鎘含量。
1.3.5 葉片和根不同化學形態鎘含量采用連續提取法[13],分別用下列提取液依次進行不同化學形態鎘提取:80%乙醇→去離子水→1 mol·L-1NaCl→2%乙酸→0.6 mol·L-1HCl。稱取0.2 g新鮮樣品,加入20 mL 80%乙醇提取液研磨、勻漿,并轉入離心管中,25 ℃恒溫振蕩22 h 后,于5 000 g離心10 min,倒出上清液;向沉淀加入20 mL 80%乙醇提取液于25 ℃恒溫繼續振蕩2 h,再于5 000 g 下離心10 min,倒出上清液。合并2 次上清液,蒸發至近干(2~3 mL),用于后續消解處理。離心得到的沉淀用下一種提取液進行提取,重復以上震蕩、離心提取過程。最后,經0.6 mol·L-1HCl 提取后得到的沉淀直接消解,即為殘渣態。提取得到的樣品按照1.3.3中的方法消解并測定鎘含量。
1.4 數據分析
文中所列數據為3次重復的平均值,數據表示為平均值±標準誤。采用 SPSS 20.0軟件進行數據統計和方差分析,并用Duncan法進行多重比較(α=0.05),Excel 2019制作圖表。
2 結果與分析
2.1 鎘脅迫對兩種生態型香根草葉片生長狀況、葉綠素熒光參數和SPAD值的影響
圖1顯示,800 μmol·L-1鎘處理時,VN型香根草最外側葉片開始變黃,而MT型香根草的葉片無明顯受害癥狀;2 000 μmol·L-1時,VN型香根草的葉片全部變黃、枯萎,而MT型香根草僅外側的葉片開始枯黃,中間的葉片仍為綠色,受害較輕。
同時,通過比較鎘脅迫下兩種生態型香根草葉片葉綠素熒光參數和SPAD值的變化(表1)發現,與對照(CK)相比,兩種生態型香根草葉片的葉綠素熒光參數Fo、Fm、Fv/Fm、Fv/Fo值以及相對葉綠素含量(SPAD值)均隨著鎘脅迫濃度的增加呈降低趨勢。其中,MT型香根草的Fo降幅始終未達到顯著水平,其Fm、Fv/Fm、Fv/Fo也均僅在2 000 μmol·L-1鎘處理下才比對照顯著降低,其SPAD值在各處理間均存在顯著性差異。VN型香根草的Fm在200 μmol·L-1鎘處理下就已經比對照顯著降低,Fv/Fm和Fv/Fo則在800 μmol·L-1鎘脅迫處理下就顯著降低,而其Fo降幅僅在2 000 μmol·L-1鎘處理下才達到顯著水平,其SPAD值在各處理間也均存在顯著性差異。在相同鎘脅迫濃度下,葉片各參數值基本表現為MT型高于VN型;在對照條件下,各參數在兩者之間均差異不顯著;在200 μmol·L-1鎘處理下,僅Fm在兩者間有顯著差異;在800 μmol·L-1鎘處理下,Fm、Fv/Fm和Fv/Fo均在兩者間差異顯著;在2 000 μmol·L-1鎘處理條件下,各參數在生態型間均有顯著性差異,此時MT型葉片Fo、Fm、Fv/Fm、Fv/Fo及SPAD值分別比相應對照顯著降低14.94%、31.15%、8.18%、25.63%和45.38%,VN型則分別顯著降低49.14%、79.04%、46.15%、77.79%和58.93%。可見,在鎘脅迫條件下,MT型香根草葉片生長狀況更好,葉片的熒光參數和葉綠素相對含量下降幅度小而緩慢,耐鎘性更強。
2.2 鎘脅迫對兩種生態型香根草葉片和根中鎘積累量的影響
表2顯示,兩種生態型香根草根部和葉片鎘積累量隨著鎘處理濃度的升高而顯著增加,但MT型的鎘積累量大多顯著低于相同濃度處理的VN型。其中,在200~2 000 μmol·L-1鎘處理下,兩種生態型香根草葉和根中的鎘含量分別為1.52~36.31 mg·kg-1和64.47~354.09 mg·kg-1,分別比對照增加了2.49~33.99倍和42.98~236.06倍,即根中鎘含量及其增幅遠高于葉片;除200 μmol·L-1處理組根中鎘含量以外,VN型葉片和根中的鎘含量均顯著高于MT型。同時,隨著鎘脅迫濃度的增加,兩種類型香根草葉片的鎘富集系數(BF)均呈先降低后升高的變化趨勢,而根中鎘的BF值則表現出逐漸顯著下降的趨勢,且在各濃度處理間多存在顯著差異。此外,兩種類型香根草鎘轉移系數(TF)呈先降低后增加的趨勢,且在2 000 μmol·L-1處理時大幅度增加,各處理濃度間多存在顯著性差異。就兩種生態型香根草而言,MT型香根草的鎘富集系數和轉移系數在大多數情況下要小于VN型。

表2 不同濃度鎘脅迫下兩種生態型香根草葉片和根部鎘積累量、富集系數和轉移系數的變化
2.3 兩種生態型香根草葉片和根中鎘的亞細胞分布特征
由表3可知,在200和800 μmol·L-1鎘脅迫下,兩種生態型香根草葉片細胞壁組分、可溶組分和細胞器組分中的鎘含量變化不顯著; 在 2 000 μmol·L-1鎘脅迫下,VN型葉片中3種組分的鎘含量和MT型葉片中細胞壁組分的鎘含量才顯著增加;VN型葉片中可溶組分和細胞器組分中的鎘含量顯著高于相應的MT型。同時,在200~2 000 μmol·L-1鎘脅迫處理下,VN型葉片中細胞壁組分的鎘占比由18.87%增加到75.93%,可溶組分的鎘占比由78.48%下降到23.84%;而MT型葉片中2種組分的鎘占比在200~800 μmol·L-1鎘處理下基本穩定,分別小于40%和小于60%,僅在2 000 μmol·L-1鎘處理時,其細胞壁組分的鎘占比大幅增加到94.62%,而可溶組分的鎘占比下降到5.26%(圖 2,A)。
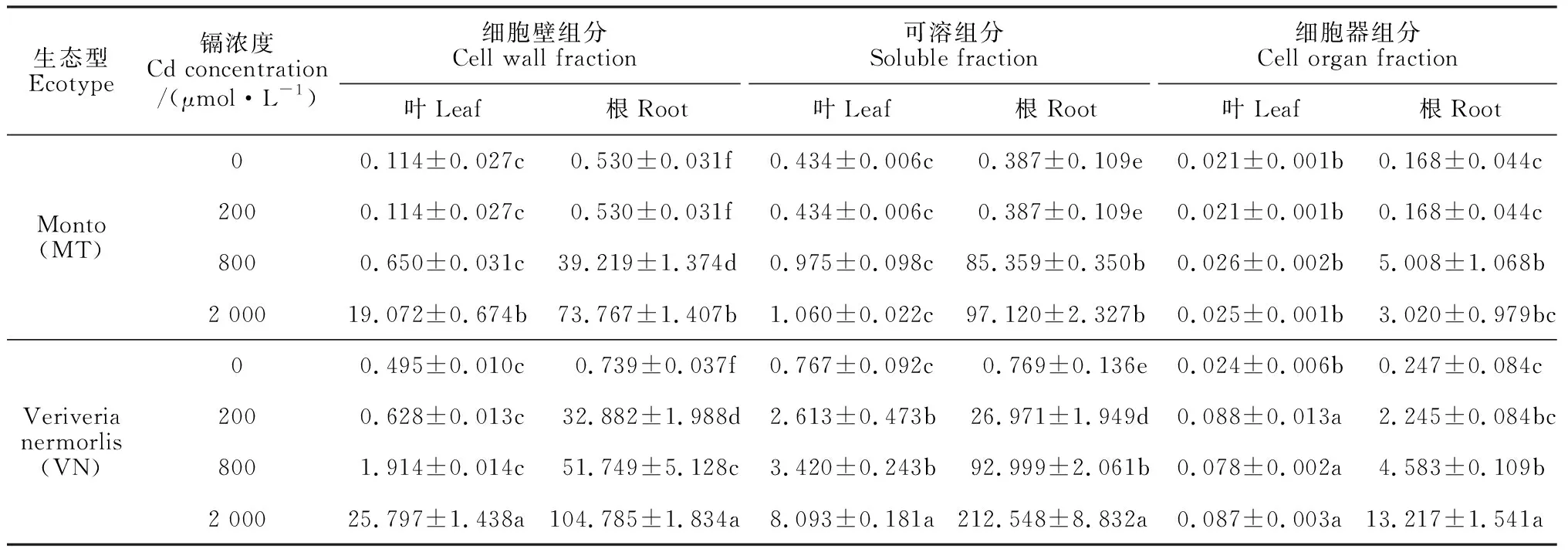
表3 不同鎘濃度脅迫下兩種生態型香根草葉片和根中鎘的亞細胞分布
在200~2 000 μmol·L-1鎘處理下,兩種生態型香根草根中的細胞壁組分和可溶組分的鎘含量均顯著增加(表 3);但MT型根中細胞壁組分的鎘占比由27.31%逐步增加至42.42%;可溶組分的鎘占比則由70.53%逐步下降至55.85%;但VN型的表現則相反,細胞壁組分的鎘占比由52.95%下降到31.70%,可溶組分的鎘占比由43.43%增加至64.30%(圖 2,B)。同時,兩種生態型香根草根中細胞器組分的鎘含量大多變化不顯著(表 3)。總體而言, 除200 μmol·L-1處理的可溶組分鎘含量外,VN型香根草根中細胞壁組分和可溶組分鎘含量顯著高于相應的MT型。
2.4 兩種生態型香根草葉片和根中各化學形態鎘的分布特征
由表4和圖3,A 可知,在200~2 000 μmol·L-1鎘脅迫下,兩種生態型香根草葉片中均以氯化鈉提取態鎘含量最高,占比均在90%左右(僅VN型在2 000 μmol·L-1鎘處理組為57.80%)。MT型的乙醇提取態、氯化鈉提取態、殘留態鎘含量在3種濃度鎘處理間無顯著差異;MT型的乙酸提取態、鹽酸提取態鎘含量在800和2 000 μmol·L-1鎘處理下保持穩定,并顯著高于200 μmol·L-1鎘處理組。VN型的乙醇提取態、氯化鈉提取態和鹽酸提取態鎘含量在2 000 μmol·L-1鎘處理下顯著高于其他鎘濃度處理組;同時,VN型的上述3種提取態和去離子水提取態鎘含量也顯著高于相應的MT型,尤其是其乙醇提取態、去離子水提取態鎘含量分別為MT型的13.2和14.3倍。
表5和圖3,B顯示,在200~2 000 μmol·L-1鎘處理下,兩種生態型香根草根中鹽酸提取態和殘留態鎘含量均小于0.3 mg·kg-1,占比均小于2%,它們的去離子水提取態和乙酸提取態鎘含量均小于25 mg·kg-1,占比均大多小于10%(VN型的去離子水提取態800 μmol·L-1鎘處理組除外);兩種生態型香根草根中4種化學形態的鎘含量及占比在各濃度鎘處理后整體變化較小,其中的乙醇提取態和氯化鈉提取態的鎘含量和占比較高,并在MT型和VN型間差異顯著。MT型和VN型各濃度處理根系的乙醇提取態鎘占比分別為69%~77%和27.5%~37.8%,氯化鈉提取態鎘占比分別為12.22%~19.59%和47.78%~54.75%;MT型200和800 μmol·L-1鎘處理組乙醇提取態鎘含量約為相應VN型的2.4倍和3.0倍,MT型的氯化鈉提取態鎘含量僅為VN型的17.7%~29.7%。
3 討 論
3.1 兩種生態型香根草葉片生長狀況、葉綠素熒光參數和鎘積累對不同濃度鎘脅迫的響應
植物體內積累過量的鎘,會降低葉綠素的合成效率、破壞葉綠體的微體結構、抑制光合系統活性,進而影響植物的生長。葉綠素熒光參數能體現植物光合作用的能力,其中Fv/Fo反映PSⅡ捕獲激發能的傳能效率,Fv/Fm反映PSⅡ反應中心原初光能轉化效率,其可以反映PSⅡ受傷害的程度,常作為植物脅迫抗性的重要評價指標[14]。BORGES等[15]研究表明鎘脅迫會抑制葉綠素的合成從而降低其含量,并能通過增加熱耗散、影響電子流傳遞等方式降低PSⅡ的活性。周蛟等[3]也發現鎘脅迫導致兩種龍葵的葉綠素含量和葉綠素熒光參數顯著下降,但鎘耐受能力更強的龍葵(SolanumnigrumL.)的Fv/Fm和Fv/Fo要顯著高于少花龍葵(SolanumamericanumMill.)。本研究表明,在800 μmol·L-1鎘處理下,MT型生態型香根草葉片無明顯受害癥狀,Fo、Fm、Fv/Fm和Fv/Fo等葉綠素熒光參數基本保持穩定,而VN型香根草最外側葉片開始變黃枯萎,以上葉綠素熒光參數也出現明顯下降;同時,在2 000 μmol·L-1鎘處理下,MT型的葉片生長狀況、葉綠素熒光參數和葉綠素含量均顯著高于VN型。這表明MT型香根草對鎘的耐受能力要強于VN型。
研究表明,不同種類、生態型或基因型植物的鎘耐受能力和鎘積累特性存在明顯差異[5]。本研究發現,MT型香根草植株根和葉中的鎘含量、富集系數和轉移系數均明顯低于VN型,這可能是MT型生態型香根草鎘耐受能力更強的主要原因。本研究中MT型和VN型香根草將大部分的鎘積累在根中,向其地上部轉移較少,轉移系數遠小于1;但其根和葉中積累的鎘含量遠高于鎘耐受能力較強的多花黑麥草(LoliummultiflorumLamk.)[7]和一年蓬[Erigeronannuus(L.) Pers]等[16]。尤其是兩種生態型香根草根部 的鎘富集和鎘積累能力與商陸(PhytolaccaacinosaRoxb.)、青葙(CelosiaargenteaL.)等鎘超富集植物[17]相當。一般而言,植物修復重金屬污染的效率取決于2個方面,即植物體對環境中重金屬的吸收富集能力以及該種植物生物量的大小[18]。香根草生物量大、根中的鎘積累量高且富含具有經濟價值的香根油,其在鎘污染修復領域具有廣闊的應用前景。
3.2 兩種生態型香根草葉和根中鎘亞細胞分布和化學形態對不同濃度鎘脅迫的響應
鎘被細胞壁或液泡區室化以減弱其活性,是植物對鎘毒害的應激性反應[19]。研究表明,秋茄[Kandeliacandel(L.)Druce]葉片和旱柳(SalixmatsudanaKoidz.)根和葉中細胞壁上積累的鎘含量占比達到 55%~70%[20-21]。這主要是因為鎘離子帶正電荷,細胞壁帶負電荷,所以鎘 容易在細胞壁處被絡合而累積[22]。也有研究表明煙草(NicotianatabacumL.)[23]、小麥(TriticumaestivumL.)[24]細胞中的鎘主要分布在可溶部分。本研究的兩種生態型香根草 葉片中大部分鎘積累在細胞壁和細胞可溶部分;可溶性組分的鎘含量占比在200和800 μmol·L-1鎘處理時超過60%,此時可能更多的鎘被區隔在葉片細胞的液泡中;但鎘脅迫濃度增加到2 000 μmol·L-1時,可溶性組分的鎘占比下降至5.26%~23.82%,此時更多的鎘被阻擋在細胞壁中,未進入細胞內。隨著鎘脅迫濃度的增加,MT型香根草根中細胞壁組分的鎘占比逐步上升,可溶組分的鎘占比逐步下降,而VN型香根草根中的鎘亞細胞分布的變化趨勢則相反。這表明MT型香根草可將更多的鎘結合到根部細胞的細胞壁中,以減輕鎘的毒害作用。
鎘等有毒重金屬被吸收到植物體內后,以不同的化學結合形態存在于植物中。不同的提取劑可提取不同形態的重金屬化合物。去離子水主要提取與水溶性物質形成的重金屬化合物,如有機酸鹽等;80%乙醇主要提取無機鹽、氨基酸鹽等可溶鹽類重金屬化合物;氯化鈉可提取與蛋白質結合或吸著態的重金屬以及果膠酸鹽重金屬化合物等;醋酸提取難溶于水的重金屬磷酸鹽;鹽酸提取草酸鹽等重金屬化合物[25]。在本研究中,氯化鈉提取態的鎘在兩種香根草葉片中均占絕對優勢,這表明進入香根草葉片內的鎘多以果膠酸鹽和蛋白質結合態存在。MT型香根草根中以乙醇提取態的鎘為主要部分,其占比為69%~77%,并始終高于VN型(27%~37%)。這暗示MT型香根草更多的鎘可能是以無機鹽、氨基酸鹽等形式被區隔在根部細胞的液泡中,以減輕鎘脅迫的傷害[19],這可能是MT型香根草對鎘耐受能力更強的另一重要原因。
4 結 論
Monto和Veriveria nermorlis 兩種生態型香根草均表現出較強的鎘耐受能力和鎘積累特性,且Monto的鎘耐受性明顯強于Veriveria nermorlis,這可能主要與其根、葉中的鎘含量、鎘富集系數和鎘轉移系數相對較低,以及葉片細胞壁組分中鎘含量占比和根中乙醇提取態鎘占比相對較高有關。
