油葵苗期抗旱相關性狀的QTL定位及候選基因篩選
2022-10-02 10:57:06石慧敏侯建華蘇飛燕王艷霞李丹丹周佳嶺
西北植物學報 2022年8期
石慧敏,侯建華,蘇飛燕,王艷霞,李丹丹,周佳嶺
(內蒙古農業大學 農學院, 呼和浩特 010019)
干旱是世界范圍內限制農作物產量的主要因素之一[1]。在干旱頻發地區,種植耐旱性較強的作物顯得尤為重要[2]。向日葵是干旱半干旱地區一種十分重要的油料作物,根系發達,具有良好的抗旱潛力[3-5],但其分布和產量仍受干旱的影響很大[6-11]。苗期是向日葵生長發育的關鍵時期,苗期受旱對向日葵形態建成、生長發育以及后期產量和品質形成都具有很大的影響,因此開展向日葵苗期抗旱性的研究至關重要。由于農作物抗旱的遺傳機制非常復雜,因此常規育種方法在該領域若想取得重大進展比較困難[12]。近年來,分子標記技術已成功地應用于數量性狀位點的鑒定(QTL),且在挖掘作物對干旱脅迫響應的重要性狀基因位點中已經有廣泛的應用[12-13]。HERVé等[14]報道了4個葉綠素濃度QTL和1個相對含水量 QTL,分別解釋了53%和9.8%的表型變異。Nishtman等[15]從PAC2×RHA266雜交的123個自交系及其親本中選取70個自交系為材料,在淹水狀態下分別檢測到葉綠素濃度和相對含水量的3個和6個QTL。在水分脅迫條件下,鑒定出7個和2個QTL。并發現葉綠素濃度和相對含水量的QTL在連鎖群10和16上存在重疊現象。最終認為兩種水分條件下不同性狀的共同QTL位點在連鎖群10上顯得更為重要。張永虎[16]構建了1張包含17個連鎖群并分布有738個標記的遺傳連鎖圖譜,兩種水分處理下檢測到30個QTL。盡管前人對向日葵干旱脅迫的研究取得了一定的進展,但是由于上述用于QTL定位的遺傳圖譜所涉及到的標記數量少、密度低,因此無法揭示向日葵耐旱性的綜合遺傳機制。隨著向日葵基因組的公布,為挖掘向日葵抗旱相關的重要候選基因提供了可能。
基于此,本研究利用前期構建的高密度分子標記遺傳連鎖圖譜,通過對150個株系組成的重組自交系群體苗期抗旱相關性狀的表型及遺傳統計分析,進行了QTL定位和候選基因的挖掘,為油葵遺傳改良奠定基礎。
1 材料和方法
1.1 材 料
本試驗所用材料,是內蒙古農牧業科學院提供的2個親本材料K55(弱抗旱性)和K58(強抗旱性),雜交后由內蒙古農業大學通過單粒傳法構建的150個株系組成的F7代重組自交系群體。
1.2 材料種植和指標測定
2018年春季,選擇籽粒飽滿的向日葵種子,用氯化汞溶液消毒浸泡5 min,用蒸餾水多次沖洗,在水分合適的培養皿內室溫催芽2 d,種植于塑料盆(250 mm×190 mm×160 mm),每盆中裝4 kg土壤(75%砂:20%營養土壤:5%蛭石),種植前澆透水,每盆留苗8株,置于內蒙古農業大學人工智能溫室中。溫室內的溫度設置為25 ℃,濕度設置為40%。試驗設置干旱脅迫和正常澆水(對照)2個處理,每個處理設置3個重復。在向日葵幼苗生長到3片葉時開始進行干旱脅迫處理,脅迫期間每3 d 測定1次土壤含水量,干旱脅迫土壤含水量控制在5%~10%,正常澆水土壤含水量控制在15%~20%,對照條件下以最高土壤含水量20%,脅迫條件下以最高土壤含水量10%來計算每次澆水量,澆水量=(土壤最高含水量-土壤測定含水量)×土壤風干重量,土壤含水量和澆水量見表1。
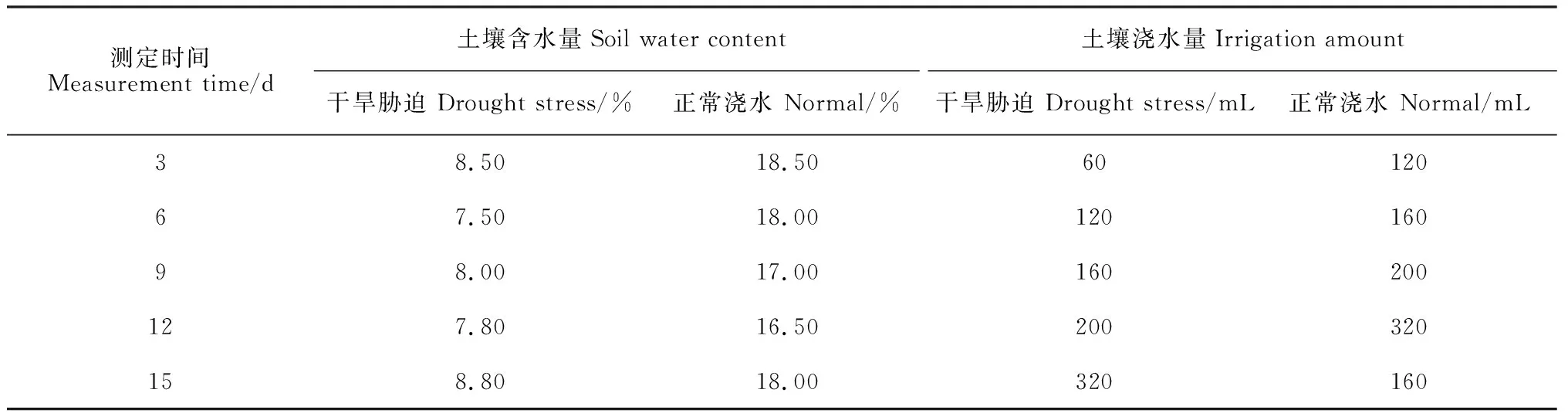
表1 土壤含水量及澆水量對照表
脅迫15 d之后,每個材料隨機選取5株,對正常澆水和干旱脅迫兩種水分條件處理下的向日葵幼苗的葉片相對電導率、葉綠素含量、葉面積、葉片相對含水量、根長共5個性狀進行測定。根長用根系掃描儀(萬深 LA-S) 測定,葉綠素含量(SPAD值)使用葉綠素儀(TYS-A) 測定,葉面積用葉面積系數法測定,葉片相對電導率用DDS-11A電導儀測定;采用飽和稱重法測定葉片的相對含水量[17],計算公式為:
RWC=[(Wf-Wd) / (Wt-Wd)]×100%
其中,Wf為葉片鮮重,Wd為葉片干重,Wt為被水飽和后的葉片重量。
1.3 數據統計分析
運用Excel 2019軟件對表型數據進行錄入整理,利用SPSS23.0軟件進行表型性狀分析、相關分析等,利用JoinMap4.0,采用CIM(復合區間作圖法)定位方法對向日葵幼苗性狀進行QTL定位。
1.4 QTL定位
本研究定位所使用的高密度遺傳連鎖圖譜是呂品等[18]于2017年以油用向日葵強抗旱自交系K58為父本,弱抗旱自交系K55為母本進行雜交獲得150個F7重組自交系(RIL)群體所構建的。該圖譜包含4 912個SNP標記和93個SSR標記,分布于向日葵17個連鎖群上,總長2 425.05 cM,相鄰2個標記間的平均距離0.49 cM。 采用JoinMap4.0軟件對RIL定位群體的所有基因型數據進行連鎖分析。使用復合區間映射(CIM)方法[19]來識別QTL。用PT檢驗1 000次設定閾值,首先考慮0.99置信度對應的LOD閾值,若沒有定位區間則考慮0.95置信度對應的LOD閾值;若沒有定位區間則考慮0.90置信度的閾值。若仍沒有結果則沒有考慮PT檢驗的結果,手動降低閾值到3.0;若3.0沒有區間則降到2.5或2.0。檢測到的各性狀QTL位點命名方法為:q+所測性狀的英文名縮寫+連鎖群位置+QTL編號。
1.5 候選基因篩選
利用遺傳連鎖圖譜中的所有SNP和SSR標記,確定了物理圖譜和遺傳圖譜的比對關系。在基因組上檢測到的與QTL的置信區間一致的區域被認為是QTL區域,位于QTL內的基因被定義為QTL的候選基因,如Chao等[20]的方法所述。本研究選擇掃描標記區間內基因組區域對應的候選基因,以2017年發布的向日葵基因組作為參考基因組進行比對[21],并利用GO、KEGG、COG、NR、pam、Swiss-Prot等數據庫進行功能注釋。
2 結果與分析
2.1 表型性狀分析
圖1顯示,兩個親本和RIL群體中的150個材料在正常澆水和干旱脅迫條件下5個抗旱相關性狀的正態分布結果。從圖1可以看出,所有性狀均呈連續分布,且呈現正態分布或偏正態分布,符合QTL定位的要求。
從表2可以看出,在正常灌水條件下,兩親本間的平均值差異不是很明顯;強抗旱種質K58葉片相對電導率、葉綠素含量、葉片相對含水量、葉面積和根長等5個性狀均高于K55(弱抗旱種質),RIL群體的變異系數為7.6%~45.4%。在干旱條件下,K58(強抗旱種質)的所有性狀也均高于K55(弱抗旱種質),RIL群體的變異系數在12.0%~42.2%之間。
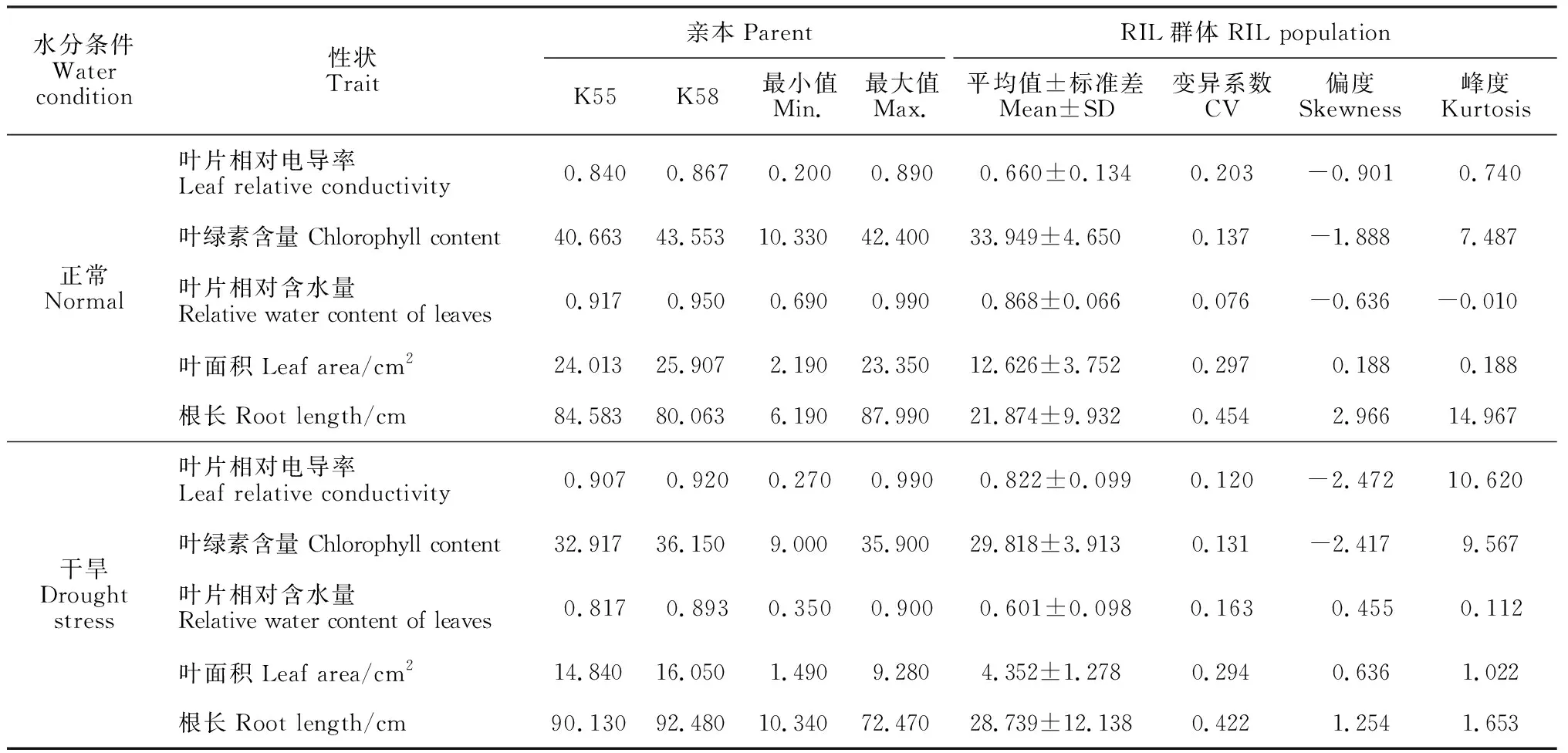
表2 正常澆水和干旱脅迫條件下親本和RIL群體各性狀指標
從表3可知,正常澆水條件下,葉片相對電導率與葉綠素含量呈極顯著正相關,與葉片相對含水量呈顯著性負相關,但未達到極顯著水平;干旱脅迫條件下,葉片相對電導率與葉綠素含量仍呈極顯著正相關;此外,根長與葉面積呈顯著性正相關,但未達到極顯著水平。

表3 正常澆水和干旱脅迫條件下RIL群體各性狀間的相關分析
2.2 QTL定位
兩種水分條件下向日葵苗期抗旱相關性狀的QTL定位結果顯示,在RIL群體中共有11個QTL位點,正常澆水條件下5個QTL,干旱脅迫條件下6個QTL。定位于5號連鎖群上的位點最多,為3個QTL位點。各性狀的表型貢獻率處于0.768%~7.547%之間(表4,圖2)。
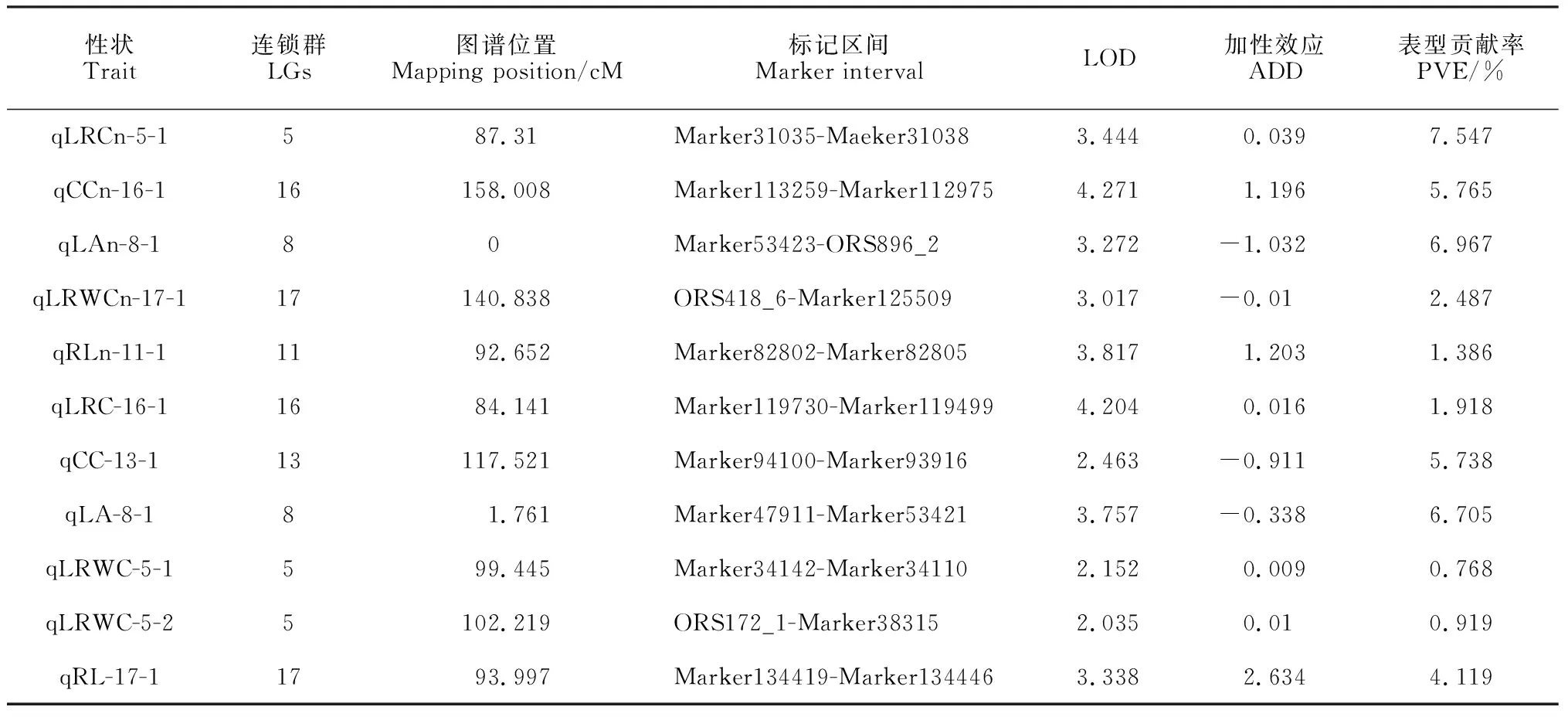
表4 兩種水分條件下苗期各性狀的QTL定位結果
正常灌水條件下檢測到的5個QTL位點中,葉片相對電導率、葉綠素含量、葉面積、葉片相對含水量和根長各檢測到一個QTL位點。位于第16連鎖群上與葉片相對電導率緊密關聯的qCCn-16-1位點LOD值最大,為4.271,表型貢獻率為5.765%;第5連鎖群上與葉綠素含量緊密相關的qLRCn-5-1位點表型貢獻率最大,為7.547%。
干旱脅迫條件下檢測到的6個QTL位點中,葉片相對含水量檢測到2個位點,葉片相對電導率、葉綠素含量、葉面積和根長各1個QTL位點。位于第8連鎖群上與葉面積緊密相關的qLA-8-1的位點表型貢獻率最大,為6.705%;位于第16連鎖群上與葉綠素含量緊密相關的QTL位點LOD值最大,為4.204。
2.3 與干旱相關的候選基因
本研究將檢測到的位點對應的QTL區間序列與NCBI上發表的向日葵基因組[21]進行比對,利用GOG、GO、KEGG、NR等數據庫對候選基因進行基因功能注釋。COG(同源蛋白簇)主要注釋到以下幾條:3條參與翻譯后修飾、蛋白質更新、伴侶;1條參與脂質運輸和代謝;5條參與無機離子的運輸與代謝;3條參與信號轉導機制;1條參與翻譯、核糖體結構和生物發生;1條參與防御機制;1條參與復制、重組和修復;1條參與次生代謝產物的生物合成、轉運和分解代謝;3條參與碳水化合物的運輸和代謝;1條參與輔酶的運輸和代謝。在兩種水分條件下,共注釋和篩選到62個重要的候選基因,候選基因位于3個連鎖群(8、13、16)的3個QTL內,與3個性狀(葉片相對電導率、葉面積、葉綠素)相關(表5)。

表5 與干旱脅迫相關的重要候選基因

續表5 Continued Table 5
基于基因功能注釋,發現許多基因與干旱脅迫有關。位于8號染色體上的QTL qLA-8-1區間內的候選基因rna22964編碼熱擊蛋白90-5,該基因參與翻譯后修飾和蛋白質更新;rna23294編碼WRKY轉錄因子2;rna22215編碼細胞色素P450 90A1;rna23271編碼細胞色素P450 82G1;rna23019和rna23004均編碼乙烯反應性轉錄因子RAP2-7;rna22100編碼E3泛素蛋白連接酶RHC2A;rna22868編碼WRKY轉錄因子26;rna22783編碼水通道蛋白TIP2-1,參與碳水化合物的運輸和代謝;rna22224編碼脯氨酸轉運蛋白3;rna22878編碼細胞色素P450 71A1;rna22577編碼小熱激蛋白;rna22661和rna22193分別編碼脫落酸受體PYL2和PYL4;rna22948編碼ABC運輸者G家庭成員31。位于13號染色體上與葉綠素含量緊密關聯的QTL位點qCC-13-1區間內的候選基因rna40140編碼水通道蛋白TIP4;1,參與碳水化合物的運輸和代謝;rna40077編碼ABC轉運蛋白G家庭成員15,該基因參與防御機制。位于16號染色體上的QTL位點qCCn-16-1的區間內的候選基因rna49909編碼光調節蛋白。
3 討 論
3.1 表型性狀分析
作物抗旱性是一種綜合性狀,單一指標往往不具有代表性。因為所有單個抗旱的生物學過程最終都將反映在植物生長及其最終產品(產量)上[22],因此,選擇多個指標來綜合評價作物的抗旱性才較為客觀。本研究選取了5個與向日葵抗旱相關的指標進行QTL定位;干旱脅迫下,K58所有性狀均高于K55,說明K58具有較強抗旱性。此外,本研究利用5個與抗旱相關的性狀對RIL群體進行抗旱性評價,但是有些性狀間相關性不大,這可能是試驗誤差造成的。由表2可以發現,兩種水分條件下葉片相對電導率與葉綠素含量均呈極顯著正相關,且干旱脅迫下,相關系數增大,說明二者與干旱脅迫具有密切的關系,因此認為這兩個指標是衡量向日葵干旱脅迫的重要指標,這與張海燕等[23]在甘薯中的研究結論一致。
3.2 油葵抗旱性的遺傳基礎和QTL
近幾十年中,QTL定位是植物數量性狀位點挖掘中十分重要的方法之一,并且已經開發了多種遺傳結構不同的群體進行定位。RIL是永久性分離群體,故常作為 QTL定位群體[24]。在本研究中,對RIL群體進行表型分析,并在正常澆水和干旱脅迫兩種條件下對油葵苗期的5個抗旱相關性狀進行QTL定位,共檢測到油葵5條染色體上與抗旱相關的11個QTL位點。位于8號染色體上與葉面積緊密相關的2個位點qLAn-8-1和qLA-8-1在兩種水分條件下被重復檢測到,表明所檢測到的QTL是穩定的。尚未定位到與前人研究一致的抗旱相關性狀的QTL位點。本研究在干旱條件下于5號染色體檢測到與葉片相對含水量相關的QTL,HERVé等[14]于5號染色體上也定位到了控制葉片含水量相關的1個QTL位點。Nishtman ABDI等[15]在正常澆水和干旱條件下分別在16號染色體上定位到控制葉綠素含量1個和2個QTL,而本研究也在正常澆水條件下也于16號染色體上檢測到1個控制葉綠素含量的QTL,這2個性狀雖然與前人研究定位到同一個染色體上,但物理位置卻不相同,QTL標記區間的差異可能是由于圖譜之間的標記種類和密度不一致所造成的。此外,qLRWC-5-1、qLRWC-5-2和qLRCn-5-1均定位于5號染色體上;qLRC-16-1和qCCn-16-1均定位于16號染色體上,qLRWCn-17-1和qRL-17-1均定位于17號染色體上,這些QTL之間雖然控制著不同的性狀,但是卻定位到同一染色體上,這些結果從各性狀間的相關分析中也可以看出,兩種水分條件下葉片相對電導率與葉綠素含量均呈極顯著正相關;正常澆水條件下葉片相對電導率與葉片相對含水量呈顯著性負相關,由此也可以看出相關性較高的性狀間,可能是由同一染色體調控的,且該結果與Nishtman等利用RIL群體在水分脅迫下獲得的結果一致[15]。
3.3 候選基因
從大量候選基因中選擇重要的候選基因往往是一項困難的任務,然而,QTL信息和基因表達變異的整合是一種常見策略[25-26]。在此基礎上本研究推測了一些可能與干旱脅迫密切相關的重要候選基因。位于8號染色體上與葉面積緊密關聯的QTL位點上注釋到的rna23019和rna23004均編碼乙烯反應性轉錄因子RAP2-7;Somayeh Najaf等[27]對向日葵的AP2/ERF基因進行全基因組識別后,選取9個AP2/ERF基因,通過qPCR驗證所選基因在不同非生物脅迫條件下葉片和根組織中的表達情況證實AP2/ERFs基因能有效抵抗非生物脅迫。rna22661和rna22193分別編碼脫落酸受體PYL2和PYL4;已知PLY參與ABA響應干旱脅迫的信號傳導過程[28]。研究發現ABA受體PYL4先前被檢測到參與了植物對各種脅迫的響應。過表達TaPYL4可以提高小麥的抗旱性[29]。rna23294編碼WRKY轉錄因子2,研究發現CsWRKY2參與了茶樹的干旱脅迫,當使用外源ABA后CsWRKY2的表達得到了增強,當使用ABA合成抑制劑時,CsWRKY2的表達受損[30]。另外,ThWRKY2可在干旱脅迫下啟動ThERF1(乙烯響應因子)基因的表達,該基因編碼一種新的乙烯響應因子,并且對干旱脅迫在內的非生物脅迫進行負調控。在芥菜中過表達ThERF1增加了植物的蒸騰速率,導致植物對干旱脅迫更敏感[26]。rna22783和rna40140分別編碼水通道蛋白TIP2;1和TIP4;1。有研究結果表明,HvTIP2;1和HvTIP4;1在大麥對干旱脅迫條件的適應過程中具有重要作用[31]。另有研究者在水稻中發現OsTIP2;1在根部強烈表達,幾乎沒有在葉片中表達,這說明水通道蛋白調控生理過程使植物獲得抗旱性可能具有特異性[32-33]。
下一步,我們將開展精細定位,對這些候選基因進行克隆和功能鑒定,并通過群體的擴大等途徑,繼續發掘與其他性狀相關的候選基因,為加強向日葵種質資源中優異基因的深度發掘和其在育種中的利用奠定基礎。
本研究表明葉片相對電導率和葉綠素含量可以作為向日葵苗期抗旱性評價的重要指標。此外,本研究對向日葵苗期抗旱相關的5個性狀進行QTL定位,于兩種水分條件下共得到11個QTL位點,并對這些位點對應的基因組區域進行候選基因的功能注釋,共篩選到62個與干旱脅迫相關的候選基因,這些基因可作為后期重點研究的對象。
