生態恢復對石漠化地區土壤有機碳累積特征及其機制的影響
2022-10-12 13:50:42龍啟霞藍家程姜勇祥
生態學報 2022年18期
關鍵詞:生態
龍啟霞,藍家程,*,姜勇祥
1 貴州師范大學喀斯特研究院, 貴陽 550001 2 國家喀斯特石漠化防治工程技術研究中心, 貴陽 550001 3 福建師范大學地理科學學院, 福州 350007
石漠化是指由于人類活動導致植被破壞、土壤退化,并轉為裸露的巖石景觀的過程[1],被認為是中國西南喀斯特地區最為嚴重的生態環境問題之一[2]。為控制西南喀斯特地區水土流失、恢復和改善退化的喀斯特生態系統,國家實施了一系列的退耕還林生態工程治理措施。植被恢復期間土壤生態功能和質量可能會得到改善[3]。土壤有機碳(SOC) 參與了土壤的多種物理化學和生物過程[4],是土壤質量的重要組成部分[5]。同時,全球土壤有機碳庫為1500—2300 Pg,約為大氣碳庫的2倍,其微小變化都會對大氣CO2濃度的平衡和調節氣候變化有重要作用[6]。土壤有機質參與土壤團聚體的形成和破環,是團聚體穩定的重要因素[7]。土壤團聚體穩定性被認為是反映土壤質量的一個重要指標[8],其團聚過程反過來促進了SOC的物理保護[9],并在維持土壤肥力和質量方面起著至關重要的作用[10]。因此,深入理解喀斯特地區生態恢復下SOC及組分對團聚體及其結合有機碳的影響,可為石漠化地區土壤生態功能恢復、控制水土流失和土地退化提供理論支撐。
植被恢復被認為是增加SOC固存的有效措施[11]。然而,在植被恢復過程中,SOC積累仍存在很大的不確定性。植被恢復可以顯著增加[12]、減少[13]或沒有顯著改變[14]SOC含量。這可能歸因于退耕造林以后的植被恢復類型、土壤深度和退耕時間等[15—16],尤其是SOC復雜的化學或物理組分的變化[17]及其與團聚體的相互關系。由耕地到原始森林的植被自然演替,喀斯特地區SOC含量逐漸增加[18],但耕地轉變為人工林SOC含量略有下降[19]。因此,深刻認識生態恢復后的土壤有機碳庫變化與團聚體關系對制定喀斯特地區合理的生態恢復策略具有重要意義。土壤SOC包含不同穩定性和周轉率的組分。土壤易氧化有機碳(Easily oxidizable organic carbon, EOC) 是土壤有機碳庫中易被氧化且活性較高的組分[20],SOC與EOC之差可以用來表征非活性有機碳(Non-liable organic carbon, NLOC) 或惰性有機碳。土壤SOC按顆粒分組可分為顆粒有機碳(Particulate organic carbon, POC) 和礦物結合有機碳(Mineral associated organic carbon, MOC),POC是新鮮動植物殘體和腐殖化有機碳過渡的碳庫,也可以表征土壤活性有機碳[21],MOC則屬于惰性有機碳庫,在一定程度上可以表征土壤碳庫穩定性[22]。SOC和植物殘體被認為是團聚體形成的重要膠結劑,顯著影響團聚體穩定性[23],反之,團聚體在形成過程中伴隨著新鮮有機物的不斷輸入,對活性碳組分的保護和固定發揮重要的作用[24—25]。一些研究主要聚焦于團聚體穩定性與不穩定和活性碳組分的關系。土壤活性有機碳組分與團聚體穩定性密切相關[26]。Wang等[27]認為SOC的活性組分(EOC) 和生物活性組分與>2 mm團聚體含量和團聚體穩定性呈正相關,同時活性有機碳通過改善土壤團聚體結構來增強團聚體對有機碳的物理保護[28]。POC構成大團聚體的核心,參與團聚體的周轉和形成,并受到團聚體的物理保護[24]。然而,穩定有機碳組分與團聚體組成和穩定性的關系較少受到關注。Wu等[10]研究發現較不穩定有機碳組分,化學穩定性有機碳組分對團聚體穩定性的影響更顯著,穩定性有機碳促進>2 mm團聚體的形成,>2 mm團聚體的物理保護降低了SOC的損失。可見,土壤有機碳組分對團聚體組成和穩定性的影響存有一些爭議。此外,不同粒徑團聚體有機碳對SOC貢獻大小存在差異[29]。羅曉虹等[30]認為有機碳主要儲存于<0.053 mm粒徑中,以<0.053 mm粒徑對SOC貢獻率最大。相反,周方亮等[31]認為>2 mm團聚體有機碳含量最大,Shen等[32]研究表明>2 mm團聚體有機碳對SOC的貢獻率最高。然而,已有研究缺乏從土壤碳組分對團聚體及其結合有機碳的影響的角度去解析生態恢復下SOC積累特征及其機制。
中國西南喀斯特地區是全球碳酸鹽巖集中分布區面積最大,以石漠化為特征的土地退化嚴重。區域尺度上的生態恢復工程促進了植被恢復,顯著提高了區域植被碳儲量,表明中國西南喀斯特地區固碳潛力巨大[33]。已有的研究主要關注喀斯特地區植被恢復/土地利用對SOC及其活性組分、團聚體有機碳的分布特征的影響[18,34—37],一些研究發現,喀斯特地區植被自然恢復可以促進SOC積累[18,38],并促進團聚體形成和提高團聚體的穩定性[39—40]。鮮有將人工恢復和自然恢復進行對比,研究喀斯特石漠化地區生態恢復下土壤SOC組分變化(尤其是POC和MOC) 與團聚體關系的報道。貴州花江小流域為典型喀斯特高原峽谷干熱河谷石漠化地區,自1999年以來,該地區開始實施退耕還林工程,使得大量坡耕地退耕為造林地。其中自然退耕的次生林和人工種植的花椒林為該地區典型的主要生態恢復模式。鑒于此,本文主要探究土壤SOC組分與團聚體及其結合有機碳的關系,以期更好地從團聚體尺度去理解喀斯特石漠化地區生態恢復后SOC積累特征及其機制,為石漠化地區植被恢復重建和改善土壤質量提供理論支撐。
1 材料與方法
1.1 研究區概況
研究區域位于中國貴州省安順市關嶺自治縣和貞豐縣交界的花江干熱河谷小流域(25°39′13″—25°41′00″ N, 105°36′—105°46′107 30″ E, 圖1)。研究區為亞熱帶河谷氣候,年平均氣溫18.4℃,年降水量約1100 mm,降雨時空分布不均,集中在5—10月份。花江示范區主要發育在三疊系碳酸鹽巖組為主的法郎向斜構造上,地勢西高東低,海拔500—1200 m,為典型的喀斯特高原峽谷地貌。區域植被覆蓋率低,巖石裸露率高達85%以上,為典型的中—強度石漠化地區。區域母巖以白云質石灰巖和泥質灰巖為主,土壤類型以鈣質石灰土為主,土層薄且分布不均。研究區土地利用以林地和耕地為主。林地主要為自然退耕的次生林和人工種植的經濟林。其中次生林為坡耕地退耕15—20 a,主要樹種為欒樹(Koelreuteriapaniculata)、香椿(Toonasinensis)、漿果楝(Cipadessabaccifera)、毛桐(Mallotusbarbatus)、八角楓(Alangiumchinense)、油桐(Verniciafordii)、粗糠柴(Mallotusphilippensis) 等,受人為干擾較少。花椒林是當地兼顧生態與經濟效益的主要經濟林,為坡耕地退耕后連續種植15—20年的純林,已成為當地主要造林樹種。花椒林的經營管理較為粗放,使用較多的除草劑、農藥和復合肥[41],每年定期進行適當修剪管理,以保證花椒有效地采光與通風。耕地主要是種植歷史50年以上的玉米,施用化肥、復合肥和農家肥混合,定期進行人為翻土、除草,采用傳統耕作方式進行土壤管理。
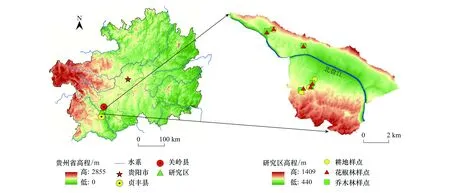
圖1 研究區采樣地理位置Fig.1 The geographic location of the sample in the study area
1.2 樣地設置及采樣
2018年11月在示范區內設置了6個樣地,每個樣地設置包括耕地、花椒林和次生林。每個樣地中的次生林和花椒林與耕地相鄰。每個樣地每種土地利用分別設置3個樣方,每個樣方按照5點混合采樣法,分別采集0—10 cm和10—20 cm土層樣品,共108個土樣。采用小鏟子分土層采集大約2 kg土壤樣品,裝入密封袋混合成一個土樣,帶回實驗室自然風干測定土壤理化性質。使用100 cm3不銹鋼環刀在每層采集約1 kg原狀土壤,為保證土壤原狀結構,使用塑料盒帶回實驗室,取出樣品沿自然結構面掰成約<10 mm的土壤顆粒,自然風干后去除土樣中動植物殘茬和礫石,進行顆粒分級及團聚體測定。土壤容重采用環刀法進行取樣。
1.3 實驗分析與測試
土壤團聚體測定[42]:稱取100 g風干土樣,置于2 mm、0.25 mm和0.053 mm組成的套篩上,進行2 min的上下均勻震蕩(30次/min),殘留在不同篩子上的土壤用蒸餾水洗到鋁盒中,將鋁盒中的土壤置于60℃的烘箱中烘干至恒重,最后獲得4個團聚體粒徑組分,分別稱重并計算其占全土的百分比后,參考SOC的測定方法,將分離后的各粒徑土壤團聚體有機碳含量進行測定。
土壤樣品研磨過0.25 mm篩,并用稀HCl去除無機碳后,土壤SOC含量采用元素分析儀—穩定同位素質譜儀(Vario ISOPOTE Cube-Isoprime Elementar公司,德國) 測定。POC的測定如下[43]:稱取過2 mm風干土樣20 g,放入250 mL三角瓶中,加入濃度為5 g/L的六偏磷酸鈉60 mL,用手搖約10—15 min后使用往復式震蕩器振蕩18 h(18℃, 90 rpm/min),震蕩后的分散液倒在53 μm篩上,用純水洗至篩下水為澄清,篩上的土壤顆粒即為顆粒有機質,分離后于鋁盒中60℃下過夜烘干(24 h),并進行稱重和計算其占全土的百分比。將>53 μm土壤顆粒磨碎過0.149 mm篩,取一定重量樣品測定其有機碳含量,乘以其占全土的百分比計算出POC含量(g/kg)。土壤MOC含量為SOC與POC之差。土壤EOC采用333 mM KMnO4測定[44],土壤NLOC為SOC與EOC之差。
1.4 數據計算
土壤有機碳儲量計算公式[45]如下:
OCs=BD×D×OC×10-2
式中,OCs為土壤SOC、POC、EOC、MOC、NLOC的儲量(kg/m2),BD為土壤容重(g/cm3),D為土層厚度(cm),OC為土壤SOC、POC、EOC、MOC、NLOC的含量(g/kg)。
土壤有機碳組分分配比例計算公式[46]如下:
式中,Fc為土壤有機碳組分分配比例(%);OCi為土壤有機碳i組分含量(g/kg);SOC為土壤有機碳含量(g/kg)。
土壤團聚體有機碳貢獻率、MWD和MGD計算公式[36]如下:
式中,Fa為土壤團聚體有機碳貢獻率(%);ASOCi為i級粒經團聚體有機碳含量(g/kg);Ai為i級粒經團聚體含量(%);SOC為土壤有機碳含量(g/kg);MWD為平均重量直徑(mm);MGD為幾何平均直徑(mm);xi為i級團聚體平均直徑(mm);wi為i粒徑團聚體重量百分比(%)。
1.5 統計分析
使用SPSS 26 軟件進行統計分析,采用單因素方差分析(One-way ANOVA) 和多重比較LSD法分析不同土地利用土壤碳組分含量和儲量、團聚體組成和穩定性、團聚體有機碳含量及有機碳貢獻率之間是否顯著差異(P<0.05)。采用皮爾遜相關分析團聚體組成、穩定性、團聚體有機碳和土壤碳組分之間的關系(P<0.05)。
2 結果與分析
2.1 土壤碳組分特征
生態恢復后,SOC及其組分含量和儲量顯著提高(MOC除外)(圖2)。SOC及其組分含量和儲量的變化模式在兩個土層(0—10 cm和10—20 cm) 相似,基本呈現次生林>花椒林>耕地。在0—10 cm土層,花椒林和次生林的土壤SOC、POC、EOC、NLOC含量分別增加了16.6%、68.2%、26.8%、14.7%和56.5%、151.7%、98.2%、48.5%,儲量分別增加了12.7%、62.0%、22.2%、11.0%和47.5%、134.5%、86.3%、40.2%。在10—20 cm土層中,土壤碳組分的變化趨勢與0—10 cm土層相似,但相對于0—10 cm土層,10—20 cm土層碳組分的含量和儲量增加幅度較小。次生林和花椒林的土壤MOC含量與耕地均無顯著差異。花椒林和次生林下的POC和EOC等活性有機碳增加幅度大于SOC,而MOC和NLOC等穩定性有機碳增加的幅度小于SOC。
生態恢復增加了活性有機碳組分的比例,減小了非活性或惰性有機碳組分的比例(圖2)。在0—10 cm土層,花椒林和次生林的POC分配比(POC/SOC)、EOC分配比(EOC/SOC) 分別顯著增加了38.3%、7.8%和58.3%、26.2%,MOC、NLOC分配比(MOC/SOC、NLOC/SOC) 的變化趨勢與POC/SOC、EOC/SOC相反,分別下降了15.3%、1.5%和23.3%、5.1%。在10—20 cm土層,EOC/SOC、NLOC/SOC的趨勢與0—10 cm土層一致,花椒林的POC/SOC最高,但其MOC/SOC最低。
2.2 土壤團聚體組成及穩定性
在0—10 cm土層,由耕地轉為花椒林和次生林顯著增加了>2 mm團聚體含量,顯著減少了0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體含量(圖3),花椒林和次生林的>2 mm團聚體含量比耕地分別增加了162.2%和379.3%,0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體含量比耕地分別下降了20.0%、36.7%、51.8%和52.8%、81.9%、83.0%。10—20 cm土層的團聚體分布的變化規律與0—10 cm土層相同,但相較于0—10 cm土層,各團聚體分布的變化幅度相對較小。生態修復后,>2 mm團聚體含量的變化最大,其次為<0.053 mm和0.053—0.25 mm團聚體,0.25—2 mm團聚體的變化最小。
生態恢復顯著增加MWD和MGD,表現為次生林>花椒林>耕地(圖3)。0—10 cm土層,花椒林和次生林的土壤MWD、MGD分別增加了75.2%、82.6%和172.0%、265.2%。10—20 cm土層,花椒林和次生林的土壤MWD、MGD分別增加了77.2%、94.5%和92.4%、125.6%。
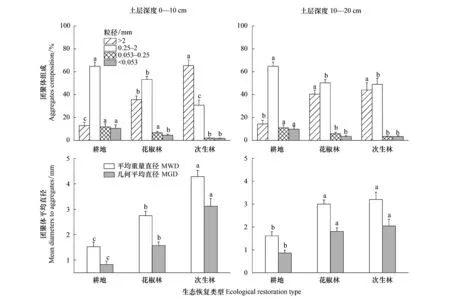
圖3 生態恢復后土壤團聚體組成和穩定性的變化Fig.3 Changes of soil aggregates composition and stability after ecological restorationMWD:平均重量直徑;MGD:幾何平均直徑
2.3 團聚體有機碳含量及團聚體有機碳貢獻率
在0—20 cm土層,耕地轉為花椒林和次生林不同程度增加了>2 mm、0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體有機碳含量(圖4)。在0—10 cm土層,花椒林和次生林的>2 mm、0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體有機碳含量依次增加了12.9%、12.2%、19.0%、16.9%和51.4%、52.5%、84.2%、62.2%,其中次生林的土壤團聚體有機碳含量顯著高于花椒林和耕地。在10—20 cm土層,不同土地利用的各粒徑團聚體有機碳含量差異不顯著(P>0.05)。生態恢復后,0.053—0.25 mm和<0.053 mm團聚體有機碳含量的增加幅度較大,>2 mm團聚體有機碳含量的增加幅度最小。在0—20 cm土層,生態恢復后各粒徑團聚體有機碳貢獻率的變化趨勢與土壤團聚體組成一致(圖4)。生態恢復顯著增加了>2 mm團聚體有機碳貢獻率,顯著減少了0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體有機碳貢獻率。在0—10 cm土層,花椒林和次生林的>2 mm團聚體有機碳貢獻率分別增加157.9%和363.7%,0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體有機碳貢獻率分別下降了22.5%、37.3%、51.0%和53.7%、78.7%、82.1%。>2 mm團聚體有機碳貢獻率增加的幅度大于<2 mm團聚體有機碳貢獻率減小的幅度。耕地中以0.25—2 mm團聚體有機碳貢獻為主體,花椒林以0.25—2 mm和>2 mm團聚體有機碳貢獻為主,次生林以>2 mm團聚體有機碳貢獻為主導。
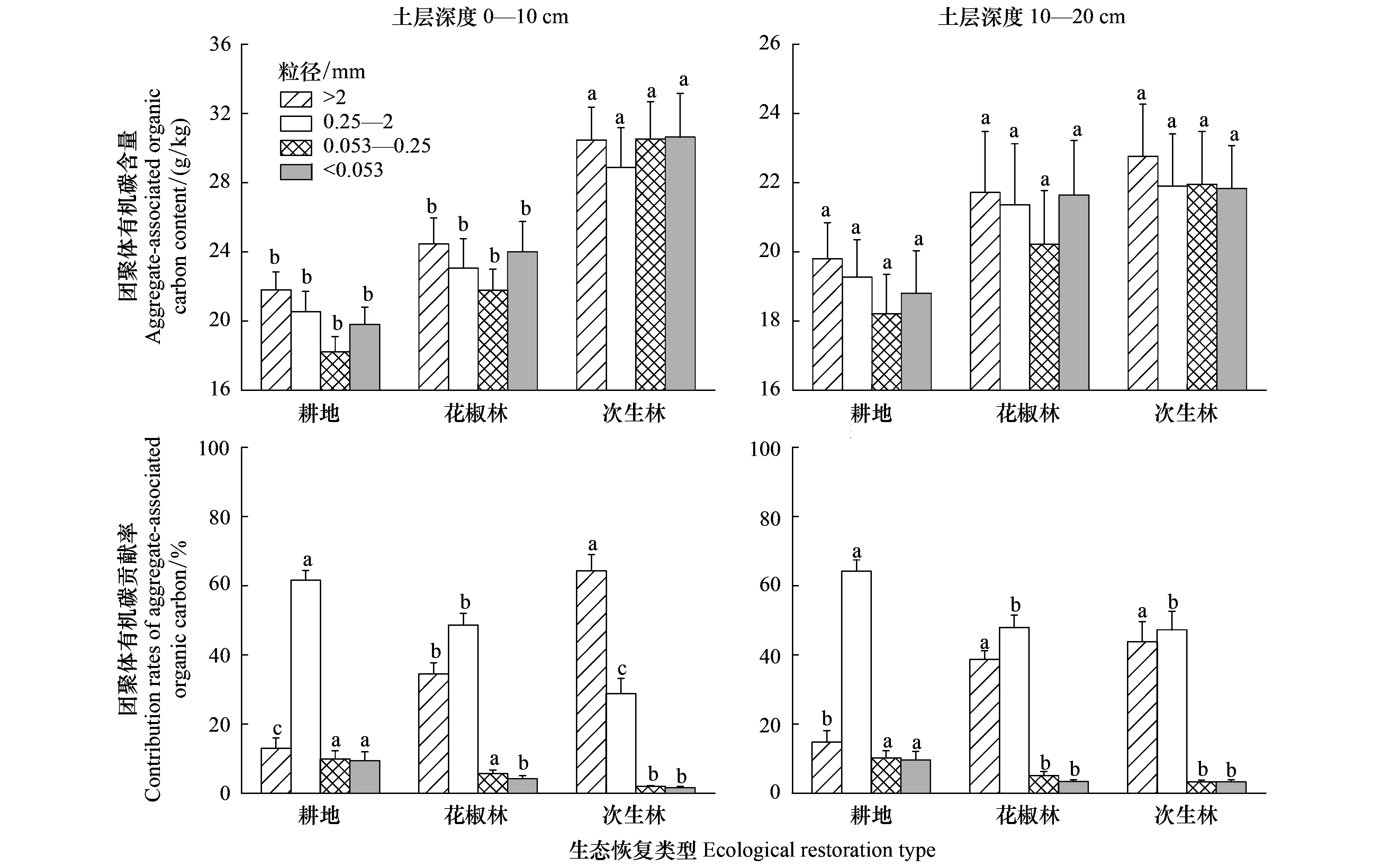
圖4 生態恢復后土壤團聚體有機碳含量及貢獻率的變化Fig.4 Variation of content and contribution rates of soil aggregate-associated organic carbon after ecological restoration
2.4 土壤碳組分間關系及碳組分與團聚體的關系
由表1可知,土壤SOC分別與POC、EOC、MOC和NLOC顯著正相關(P<0.01);與MOC(r=0.515) 相比,POC、EOC與SOC的相關系數較高(r=0.876,r=0.871);除了MOC外,土壤碳組分之間均顯著正相關(P<0.01)。
由表2可知,>2 mm團聚體有機碳貢獻率與土壤SOC、POC、EOC顯著正相關(P<0.01),與土壤NLOC顯著正相關(P<0.05),0.25—2 mm和0.053—0.25 mm團聚體有機碳貢獻率與土壤SOC、POC、EOC、NLOC顯著負相關(P<0.05),< 0.053 mm團聚體有機碳貢獻率與土壤碳組分無顯著相關(P>0.05);各粒徑團聚體有機碳貢獻率與土壤MOC均無顯著相關(P>0.05)。
由表2可知,除MOC外,土壤SOC、POC、EOC、NLOC均與>2 mm團聚體組成、MWD、MGD顯著正相關(P<0.05),與0.25—2 mm、0.053—0.25 mm團聚體組成顯著負相關(P<0.05),且POC和EOC對團聚體組成和穩定性的影響更為顯著(P<0.01);土壤MOC與團聚體組成和穩定性無顯著相關(P>0.05),土壤各碳組分均與<0.053 mm團聚體組成無顯著相關(P>0.05)。土壤碳組分與團聚體有機碳含量顯著正相關(P<0.01),相對于MOC,土壤POC、EOC與團聚體有機碳含量的相關性較強(r=0.884,P<0.01),土壤NLOC與團聚體有機碳含量相關系數也較高。土壤碳組分與團聚體有機碳貢獻率的關系與團聚體組成一致,土壤POC、EOC對團聚體有機碳貢獻率的影響更為顯著(P<0.01)。
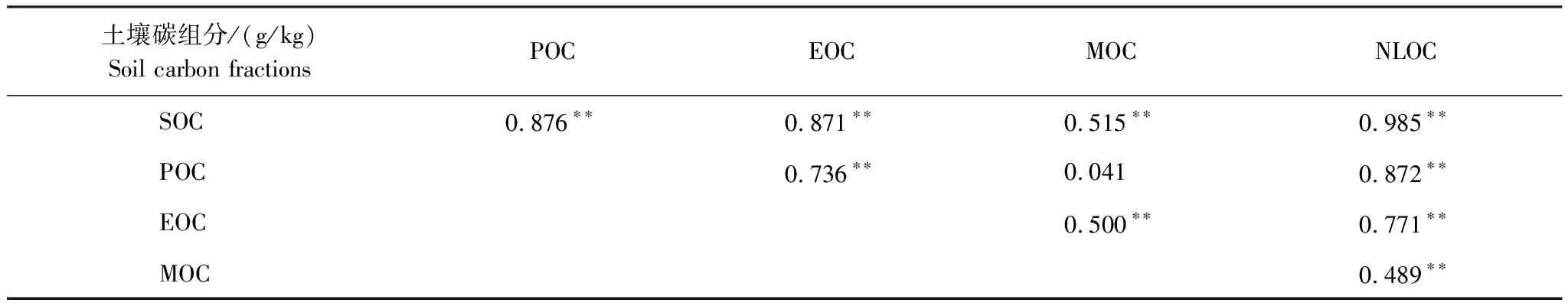
表1 土壤碳組分之間的相關性分析
2.5 土壤團聚體組成、有機碳含量與團聚體有機碳貢獻率關系
由表3可知,各粒徑團聚體有機碳含量與>2 mm團聚體有機碳貢獻率顯著正相關(P<0.05),與0.053—0.25 mm團聚體有機碳貢獻率顯著負相關(P<0.05),與<0.053 mm團聚體有機碳貢獻率無顯著相關(P>0.05);0.053—0.25 mm和<0.053 mm團聚體有機碳含量與有機碳貢獻率相關性較強。土壤各粒徑團聚體含量與其有機碳貢獻率均顯著正相關(P<0.01)。
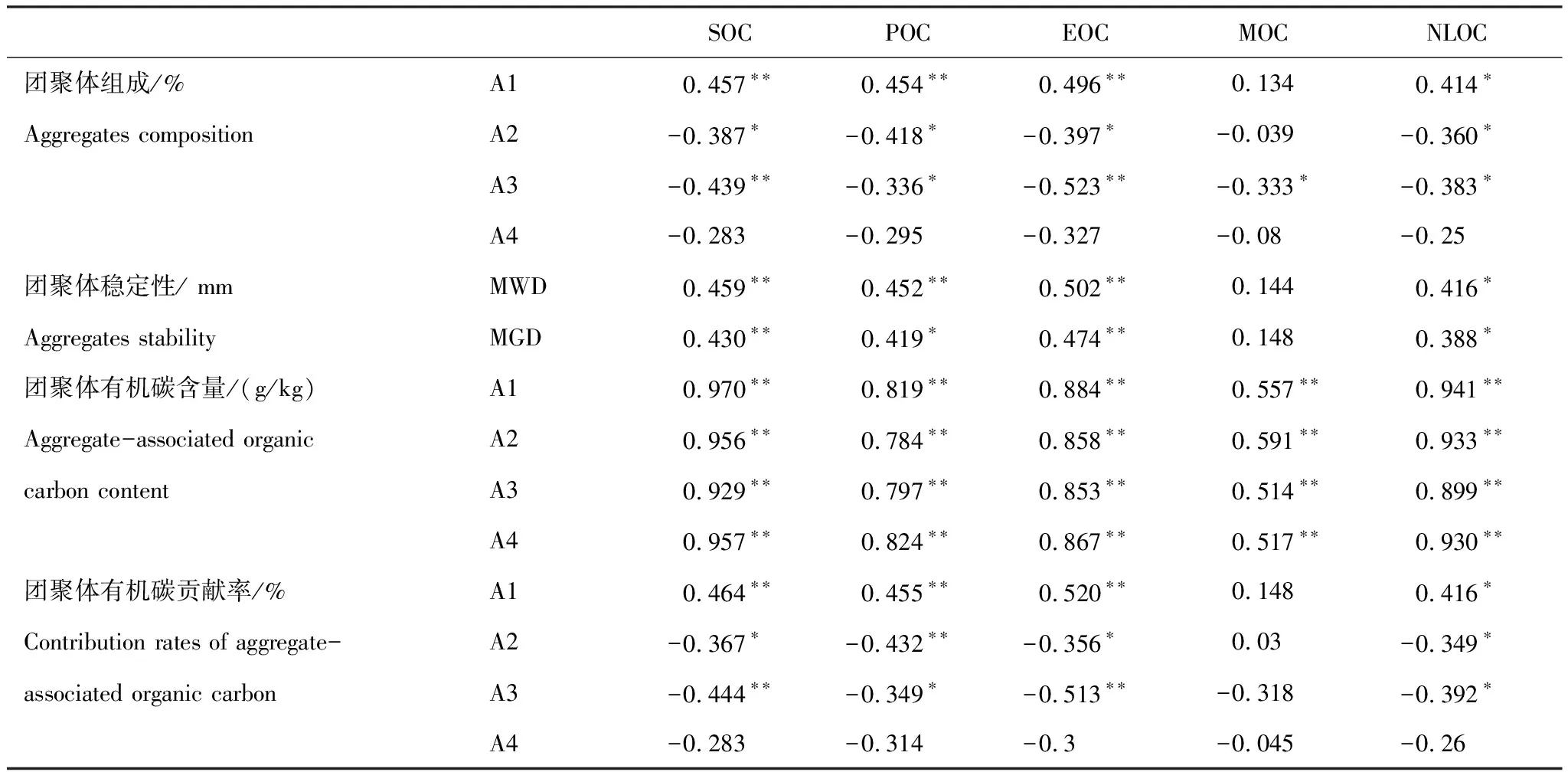
表2 土壤碳組分與團聚體組成、穩定性及團聚體有機碳的相關系數
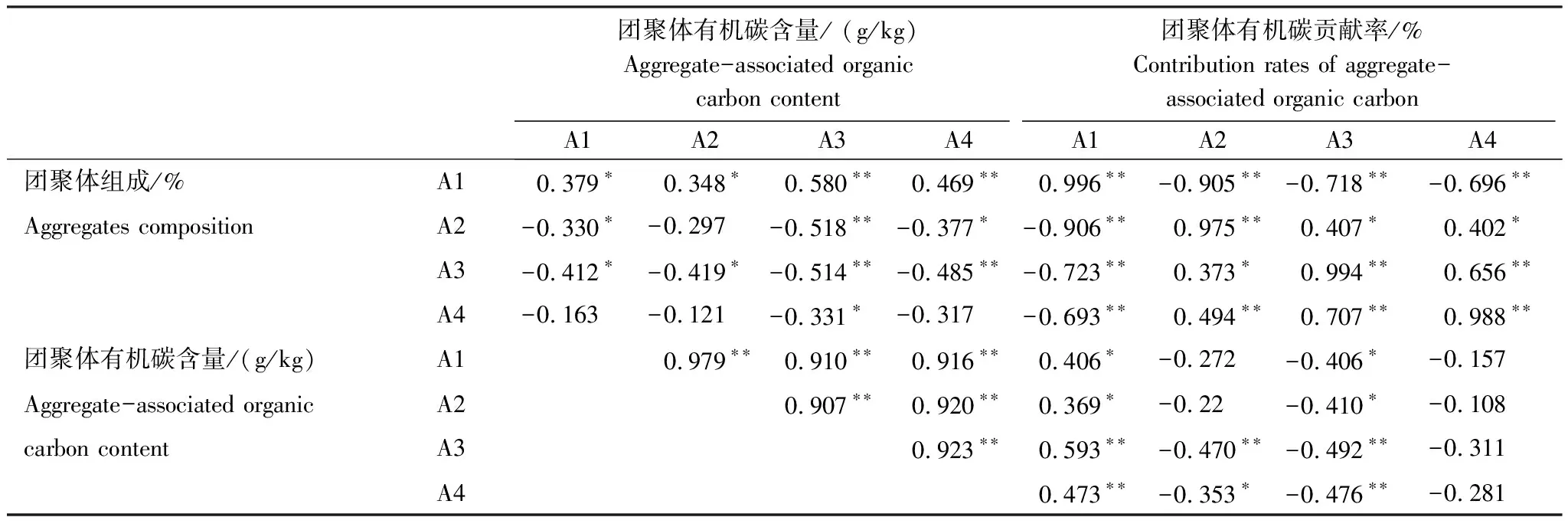
表3 土壤團聚體組成、有機碳含量與貢獻率的相關系數
3 討論
3.1 生態恢復對土壤有機碳及組分影響
土壤SOC儲量取決于凋落物、根系分泌物和生物量等碳輸入和土壤呼吸、淋失等碳輸出的平衡[47]。本研究中生態恢復后,0—20 cm土壤碳組分的含量與儲量顯著增加(MOC除外)。許多退化喀斯特生態系統的研究也證實了植被恢復促進了SOC的積累是由于退耕后較高的植物殘體和根系生物量返回到土壤中[18,38,48]。地表凋落物量和根系分泌物量為土壤有機質積累提供豐富的來源[8]。在同一研究區發現,不同種植年限(5年、10年和20年) 花椒林SOC含量高于耕地[49]。次生林和花椒林的POC、EOC含量和儲量、POC/SOC、EOC/SOC顯著高于耕地(圖2) 表明次生林和花椒林有更多的新鮮植物殘體來源碳回歸土壤。這是因為土壤活性有機碳主要來源于植物生物量和根系分泌物[43]。次生林地表生物量(2278.32 g/cm2) 大于花椒林地表生物量(681.39 g/cm2) 使得次生林SOC及組分含量較高。楊龍[50]在本研究區的研究也發現花椒林、灌木林和喬木林的生物量為721.39 g/cm2、1219.38 g/cm2和2716.83 g/cm2,SOC含量高低由地表生物量決定。相關分析(表1) 發現SOC與活性有機碳相關性較強(r=0.876,P<0.01),表明土壤SOC的增加主要依賴于土壤POC、EOC等活性有機碳的增加。另外,喀斯特地區植被恢復促使土壤有機質的積累進而促進了土壤團聚體的形成和穩定,從而增強團聚體內有機碳的物理保護[15,39,48]。植被恢復增加的植物殘體的輸入、根系生物量和分泌物也能夠促進土壤顆粒的團聚[51]。相反,長期的耕作或人為擾動一方面破壞了土壤大團聚體,降低了團聚體的穩定性[52],另一方面較高的分解速率、土壤侵蝕和有機質輸入減少導致SOC及組分含量和儲量[12]。與花椒林和次生林相比,耕地土壤中EOC、POC的含量較低(圖2) 表明耕地SOC可能具有較高的分解速率,使得活性有機碳大量損失。大量研究表明植被恢復或造林能夠顯著增加土壤活性有機碳組分[35,53—54],這與我們的研究結果一致。一方面植被下層凋落物輸入可以直接提供活性有機碳組分[55];另一方面,植被恢復提升了微生物活性,微生物迅速分解促進了植物凋落物來源碳轉化為活性有機碳組分[56]。然而,生態恢復并未顯著提升MOC含量和儲量(圖2),這與其它喀斯特地區的研究結果一致[15]。這可能是因為MOC是與粘粒和礦物質結合的有機碳組分,相對穩定且很難分解,對土地利用變化不敏感[43]。也可能與恢復時間有關,因為只有長期的植被恢復才能使有機物的輸入和輸出之間達到相對平衡,非保護性的POC才能逐步向穩定性MOC轉化,增加MOC含量[57]。次生林0—10 cm土層土壤NLOC含量和儲量顯著高于耕地(圖2)。總之,生態恢復不僅增加了土壤總有機碳含量,產生了更多的活性有機碳組分,而且也增加了非活性碳組分的含量,但SOC的增加主要歸因于土壤活性有機碳的增加。此外,自然恢復措施(次生林) 比人工恢復(種植花椒) 可以更好地促進0—20 cm土層SOC的積累。
3.2 生態恢復下團聚體對土壤有機碳變化的作用
生態恢復后,各粒徑團聚體有機碳含量均不同程度增加,其中0.053—0.25 mm和<0.053 mm團聚體有機碳含量的增加幅度較大,>2 mm團聚體有機碳含量的增加幅度最小(圖4)。這符合有機碳輸入優先向小粒徑團聚體積累而后再膠結組成較大團聚體的層級理論[26],并與其它喀斯特地區研究結果一致[37]。從團聚體對有機碳貢獻率的變化來看,也證實了較大團聚體是由較小團聚體與有機膠結物組成的。耕地土壤以0.25—2 mm團聚體有機碳貢獻為主體,而生態恢復后轉為以>2 mm團聚體對有機碳的貢獻為主體(圖4),>2 mm團聚體有機碳貢獻率的顯著增加抵消了<2 mm團聚體有機碳貢獻率的減小,最終增加了SOC含量。這表明>2 mm團聚體有機碳增加成為生態恢復后SOC增加的主要形式。土壤SOC與>2 mm團聚體含量顯著正相關(表2) 也驗證了這一點。在其它喀斯特地區的研究也發現植被自然演替恢復過程中SOC的增加主要歸因于>2 mm團聚體有機碳的積累[48]。退耕地植被恢復過程中土壤團聚體有機碳貢獻率主要受團聚體數量和團聚體有機碳含量的影響[58]。王興等[29]的研究也發現撂荒地植被恢復過程中團聚體對SOC的貢獻率80%來自>2 mm團聚體,>2 mm團聚體數量的顯著增加是該團聚體有機碳貢獻率高的主要原因,其次才是該團聚體有機碳含量的增加。本研究中,>2 mm團聚體數量、>2 mm團聚體有機碳含量分別與>2 mm團聚體有機碳貢獻率顯著正相關(表3),且團聚體數量與團聚體有機碳貢獻率關系更強(r=0.996,P<0.01),進一步證實了>2 mm團聚體數量是影響團聚體有機碳貢獻率的關鍵因子。此外,0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體有機碳貢獻率顯著減小也主要是因為團聚體數量的顯著減小(圖4,表3),這潛在說明生態恢復使團聚體有機碳含量發生了再分配過程,即生態恢復促進有機碳由<2 mm團聚體向>2 mm團聚體轉化,且這種轉化主要是由于植被恢復促進了較小團聚體膠結形成較大團聚體,從而保護了有機碳不易被微生物利用。
3.3 生態恢復下土壤碳組分對團聚體的影響
生態恢復后,>2 mm團聚體數量、MWD、MGD顯著增加,而0.25—2 mm、0.053—0.25 mm和< 0.053 mm團聚體數量顯著減少(P<0.01,圖3),這表明生態恢復能夠有效改善土壤團聚體結構和團聚體穩定性。這與Hu等[48]在重慶南川巖溶石漠化地區關于植被自然演替恢復對團聚體結構的影響的結果一致。生態恢復后植物殘體和根系生物量不斷輸入土壤,促進SOC的積累,有機碳作為團聚體主要的膠結物質促進團聚體的形成和穩定[59]。SOC及組分與>2 mm團聚體數量、MWD、MGD顯著正相關(表2) 驗證了這一點。土壤團聚體的有機膠結物質可分為瞬變性(微生物和植物來源得多糖類物質)、臨時性(根系和真菌菌絲及分泌的多糖) 和持久性膠結劑三大類[26]。有機碳組分中EOC和POC對團聚體形成和穩定性影響更大,主要是因為EOC主要來源于植物凋落物和根系分泌物的分解[43],主要包含復合多糖等有機化合物[60],POC主要來源于新鮮動植物殘體以及腐殖有機質中的臨時或過渡有機碳[61],因此EOC和POC很可能由各種臨時或暫時的膠結劑組成,它們促進了大團聚體的形成和穩定。廖林超等[62]的研究也驗證了土壤團聚體的分布和穩定性與活性有機碳關系更密切,活性有機碳是土壤團聚化的重要有機質膠結物。然而,SOC及組分含量與0.25—2 mm、0.053—0.25 mm和<0.053 mm團聚體數量顯著負相關,可能是由于初始有機碳最先在微團聚體或細顆粒中積累,隨后整合進入大團聚體[26]。NLOC和MOC含量與團聚體數量和穩定性的相關性較弱(表2),可能與其對生態恢復的響應較不敏感有關。MOC是有機物的最終分解產物,與粘粒、粉粒和礦物質結合的有機碳,相對穩定且很難分解[43],因此其可能很少參與團聚體的形成。然而,Wu等[10]研究發現較不穩定有機碳組分,化學穩定性有機碳組分促進>2 mm團聚體的形成,對團聚體穩定性的影響更顯著。NLOC作為非活性有機碳,也與顯著團聚體數量和穩定性顯著相關,但其相關性小于活性有機碳與團聚體數量和穩定性的關系(表2)。因此,生態恢復使得新鮮有機質不斷輸入,顯著提升了活性有機碳,活性有機碳作為臨時或暫時的有機膠結劑主導了>2 mm團聚體形成和穩定。生態恢復增加的有機碳及組分作為膠結物質促進了團聚體周轉、形成和穩定,反過來有機碳及組分受到團聚體的物理保護,使得SOC在不同粒徑團聚體進行再分配。因此,SOC及組分含量與團聚體有機碳含量顯著相關(表2)。生態恢復導致的地上生物量增加了有機殘體的輸入,可能通過促進大團聚體形成中作為主導膠結劑的EOC和POC的供應,加速大團聚體形成,從而導致大團聚體有機碳儲量的顯著增加。>2 mm團聚體有機碳貢獻率與土壤EOC和POC呈顯著正相關,且其相關系數高與NLOC和MOC驗證了這一點。
4 結論
(1)生態恢復顯著增加土壤有機碳組分含量與儲量(MOC除外),尤其是產生了更多的POC和EOC,土壤SOC含量的增加主要依賴于土壤POC、EOC含量的增加。
(2)生態恢復加速了有機碳從較小團聚體向較大團聚體的重新分配過程,>2 mm團聚體有機碳儲量增加是生態恢復后SOC儲量增加的主要形式,而>2 mm團聚體有機碳儲量增加依賴于其團聚體數量的大幅提升,其次才是團聚體有機碳含量的增加。
(3)生態恢復導致更多的新鮮植物殘體來源碳的輸入,可能通過促進>2 mm團聚體形成中作為主導膠結劑的EOC和POC的供應,從而改善土壤團聚體結構和促進土壤SOC的積累。
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
貴茶(2019年3期)2019-12-02 01:46:32
鄉村地理(2018年3期)2018-11-06 06:51:02
茶葉通訊(2017年2期)2017-07-18 11:38:40
河北城市研究(2015年4期)2015-08-23 11:53:06
浙江工商大學學報(2015年6期)2015-03-01 02:56:40
