馬尾松人工林土壤有機層和礦質土壤層酶活性隨雨旱季的變化
2022-10-27 02:42:26楊萬勤
生態學報 2022年19期
曹 瑞, 楊萬勤,*,袁 吉, 李 晗, 譚 波
1 臺州學院生命科學學院,臺州 318000 2 四川農業大學生態林業研究所,成都 611130
土壤酶是土壤生物化學過程的積極參與者,既能將大分子物質降解為小分子物質,又能參與腐殖質的形成,在植物殘體分解、土壤有機質形成和養分轉化等方面發揮著重要的作用[1],是土壤生產力維持與提高的關鍵。同時,土壤酶能敏感地響應生物與非生物環境因子的輕微變化,其活性變化能綜合反映土壤生物化學過程對氣候變化、土壤理化性質和土壤生物群落的響應[2—3]。然而,已有的相關研究更多地集中在礦質土壤層,而土壤有機層酶活性動態的研究相對較少[4—5]。土壤有機層是指累積在土壤表面未分解到完全分解的有機殘余物質,是土壤與植被之間進行物質轉換和能量交換最為活躍的生態界面之一,在土壤有機碳固定、物質循環和養分有效性等方面具有十分重要的作用[6]。有研究表明,土壤有機層酶活性顯著高于礦質土壤層[7—9],由于土壤有機層直接暴露在地表,其受到環境因子波動的直接影響更強烈。因此,土壤有機層酶活性對周期性環境變化的響應可能更敏感,其響應機制也可能不同于礦質土壤層,但相關機制尚待深入研究。
迄今,土壤酶活性的季節性變化已有大量研究報道[10—13],并認為土壤溫度和濕度是驅動土壤酶活性季節性變化的關鍵因子[12,14—15]。例如,Puissant等[16]研究表明,降雨減少和增溫降低了土壤含水量,抑制土壤微生物生物量,導致夏季土壤酶活性的降低。Kang等[17]研究發現,春季較高的土壤脫氫酶和脲酶活性以及秋季較高的磷酸酶和芳基硫酸酯酶活性是由林地土壤溫度和養分有效性決定的。但近期的一些研究則表明,驅動土壤酶活性變化的關鍵因素是土壤基質質量和pH[18—21]。這些不確定性可能是因為性質迥異的土層(例如土壤有機層和礦質土壤層)對環境因子的響應存在較大差異,也可能是土壤酶活性對降水等環境因子的響應很敏感,僅僅關注土壤酶活性的季節或者月動態不能反映酶活性的環境敏感性。理論上,土壤有機層和礦質土壤層可能具有明顯不同的微生物、水熱和養分有效性動態,從而使土壤酶活性對季節性環境因子變化的響應存在分異,進而影響土壤生物地球化學循環。因此,深入研究土壤有機層和礦質土壤層酶活性隨關鍵時期的變化,有助于識別主導酶活性動態的關鍵因子,有利于理解土壤生物化學循環對環境變化的潛在響應。
長江上游低山丘陵區屬于典型的亞熱帶季風氣候,降雨季節和干旱季節明顯,全年降水主要集中在夏秋兩季,具有極為明顯的雨熱同期現象[22]。馬尾松(Pinusmassoniana)是本區的主要造林樹種,在林業生產和生態建設中具有舉足輕重的地位[23]。因此,深入研究馬尾松人工林土壤酶活性隨關鍵時期的變化特征及其主要驅動因子,可為馬尾松人工林可持續經營與管理提供科學依據。然而,已有的土壤酶活性研究更多關注礦質土壤層,有關土壤有機層和礦質土壤層的酶活性隨不同關鍵時期環境因子變化的變化尚缺乏深入研究。為此,本文以長江上游低山丘陵區的宜賓高縣馬尾松人工林為研究對象,采用原位土柱培養法,同步研究了土壤有機層和礦質土壤層中與碳氮磷循環密切相關的轉化酶、脲酶和酸性磷酸酶活性隨關鍵時期的變化特征,揭示驅動土壤有機層和礦質土壤層酶活性動態的關鍵因子。
1 材料與方法
1.1 樣地設置
研究區域位于宜賓市高縣境內(104°32′—104°33′E,28°34′—28°35′N),地處長江上游低山丘陵區,地貌以低山丘陵為主,海拔高度400—500 m。氣候類型屬亞熱帶濕潤季風氣候,雨熱同期特征明顯,年平均溫度約為18℃,最高月均溫(7月)為36.8℃,最低月均溫(1月)為7.8℃,年降水量850—1200 mm,相對集中于5—10月[24]。
研究樣地布設于來復鎮毛巔坳林區的馬尾松人工林,其主要伴生樹種為香樟(Cinnamomumcamphora),林下灌木主要是梨葉懸鉤子(Rubuspirifolius)、枹櫟(Quercusserrate)、油樟(Cinnamomumlongepaniculatum)等。林下草本植物主要包括金星蕨(Parathelypterisglanduligera)、芒(Miscanthussinensis)、芒萁(Dicranopterisdichotoma)等。土壤為黃壤,基本特性詳見表1。
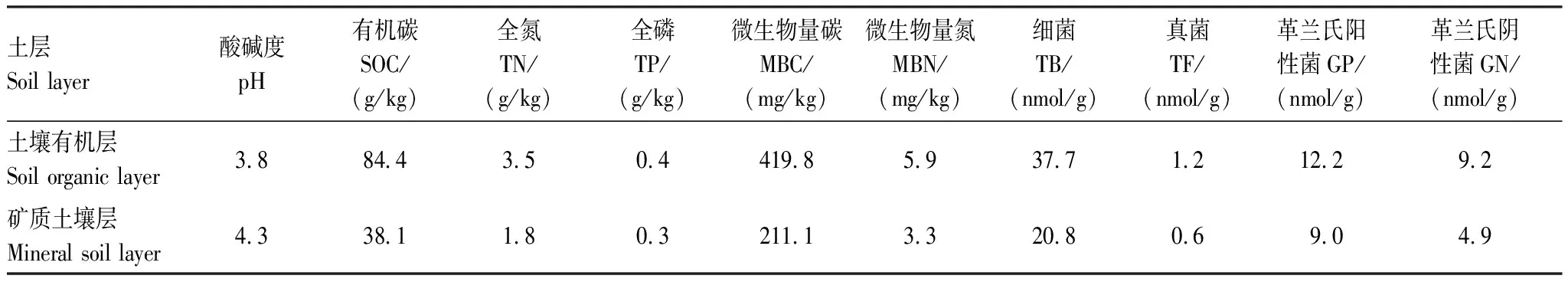
表1 土壤有機層和礦質土壤層土壤基本特性
1.2 實驗設計
在研究樣地內選擇3個間距約為100 m、長寬為10 m、面積為100 m2,且坡度、坡向和地表植被類型相似的平行樣方。在去除地表新鮮凋落物和地表植被后,采用直徑11 cm、高30 cm的PVC管垂直打入土壤采集具有完整土壤有機層和礦質土壤層的土柱。在不破壞柱內土壤結構的情況下取出土柱,立即使用0.2 mm的尼龍網布封住土柱上下兩端,隨后將保持相對完好的土柱集中培養在同一樣地中環境條件一致的地方,以避免森林環境異質性對土柱內土壤酶活性的影響。2013年9月樣地布設完畢,待實驗干擾降低后,于2014年6月開始采集樣品,持續三年。實驗期間,根據高縣多年降水資料和研究團隊前期研究結果為參考[22,25],并結合我國農歷24節氣,將采樣時間確定為雨季早期(Early rainy season,ERS,6月6日)、雨季中期(Mid rainy season,MRS,10月8日)、雨季末期(Late rainy season,LRS,12月6日)、旱季早期(Early dry season,EDS,3月5日)和旱季末期(Late dry season,LDS,4 月5日)等5個關鍵時期。
1.3 土壤采集與測定
依照實驗設計,確保采樣時間誤差不超過正負一周的前提下,在每個樣方隨機采集3根土柱并按土壤有機層和礦質土壤層分別混合,每次采集6份土壤樣品(2土層×3樣方)。土壤樣品通過低溫保存箱迅速帶回實驗室,過2 mm分樣篩后,一部分風干密封保存,一部分用自封袋密封保存于4℃冰箱測定土壤酶活性。
土壤酶活性測定參照關松蔭的研究方法進行改進[26]。轉化酶活性采用3,5-二硝基水楊酸比色法測定,用1g干土在37℃下24h水解產生葡萄糖的毫克數表示;脲酶活性采用靛酚藍比色法測定,用1g干土在37℃下24h水解產生銨氮的毫克數表示;酸性磷酸酶活性采用比色法測定,用1g干土在37℃恒溫培養條件下,24h釋放的苯酚量來計算。
土壤微生物生物量碳和氮采用改進的氯仿熏蒸-K2SO4浸提方法測定[27];土壤微生物磷脂脂肪酸含量通過對總脂質進行單相萃取后測定[28]。土壤pH(土水比為1∶2. 5)采用pH計測定。土壤有機碳含量采用重鉻酸鉀加熱法測定,全氮含量采用凱氏定氮法測定,全磷含量采用鉬銻抗比色法測定[29]。此外,土壤含水量通過稱取10g新鮮土樣烘干至恒重,用水分重量占1g干土的百分比來表示;土壤溫度采用埋設紐扣式溫度記錄器(DS1923-F5 iButton,Maxim Integrated Products, Inc., Sunnyvale, CA, USA)每2h記錄一次。
1.4 數據處理與統計分析
通過SPSS 20.0軟件,采用重復測量方差分析(Repeated Measures ANOVA)檢驗關鍵時期、年際、土層及其相互作用對土壤酶活性影響的顯著性。基于R(4.0.2),通過ggpubr軟件包,采用單因素方差分析(One-way ANOVA)對土壤酶活性不同關鍵時期的差異顯著性進行檢驗(P=0.05)。通過 SIMCA 14.1軟件,采用偏最小二乘法(PLS)評估環境因子對土壤有機層和礦質土壤層酶活性的影響程度和相對重要性。采用ggpplot 2 軟件包進行作圖。
2 結果與分析
2.1 環境因子隨關鍵時期和年份的變化特征
研究期間,三年的平均降雨量為944.36 mm,降雨主要集中在5—10月,最低的月降水量為7.6 mm,最高的月降水量為282.7 mm;年際間的降雨量無顯著差異,但月份之間和季節之間的變化明顯。三年的大氣相對濕度平均值為81.57%,并無明顯的季節差異(圖1)。觀測樣地的土壤溫度與大氣溫度變化相一致,夏季較高,冬季較低。同時,土壤有機層溫度波動幅度明顯高于礦質土壤層(圖1)。土壤含水量均呈下降趨勢,且土壤有機層下降幅度顯著大于礦質土壤層(圖1)。
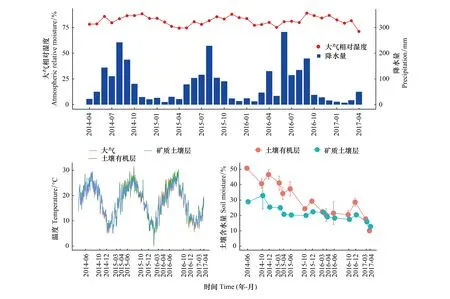
圖1 馬尾松人工林觀測樣地的土壤溫濕度和大氣溫濕度動態Fig.1 Dynamics of temperature and moisture in both air and soil in the observational plots of Masson pine plantation
2.2 土壤轉化酶活性動態
關鍵時期、年際、土層及其相互作用均對土壤轉化酶活性產生了顯著影響(表2)。除第一年外,土壤有機層和礦質土壤層的轉化酶活性隨關鍵時期發生了顯著變化,其差異顯著性取決于培養時間和土層(圖2)。土壤有機層轉化酶活性在雨季初期(ERS)顯著最高,在旱季末期(LDS)顯著最低;而礦質土壤層中,第二個培養年的轉化酶活性在雨季初期(ERS)和旱季末期(LDS)顯著高于其他關鍵時期,第三個培養年的轉化酶活性在雨季初期(ERS)顯著最低(圖2)。土壤有機層的轉化酶活性顯著高于礦質土壤層,且隨培養年際的增加表現出明顯的降低趨勢(圖2)。
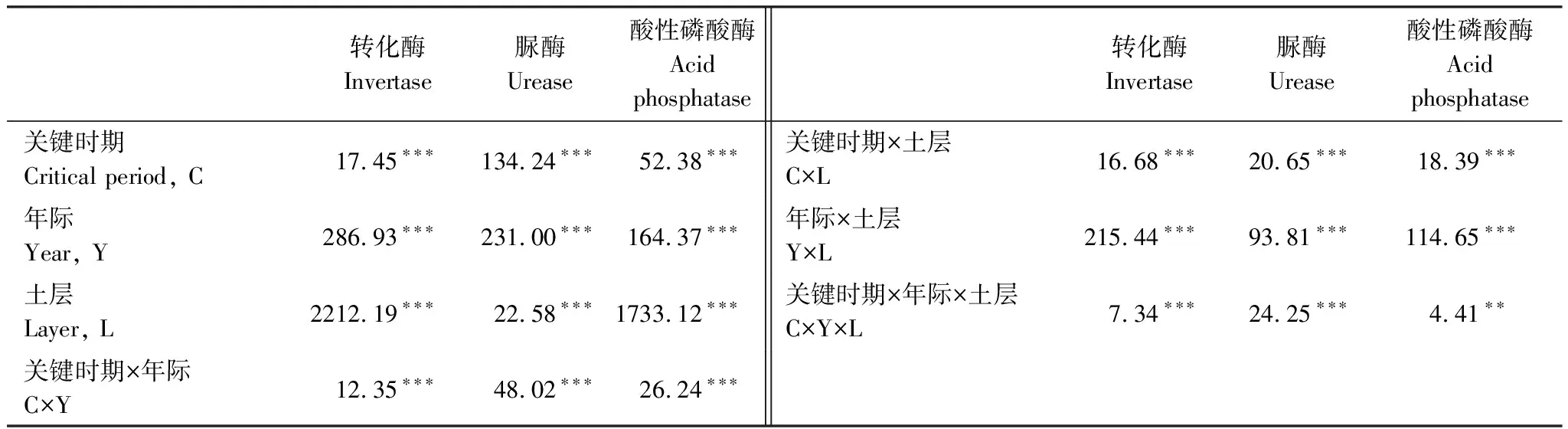
表2 關鍵時期、年際、土層及其相互作用對土壤酶活性影響的重復測量方差分析
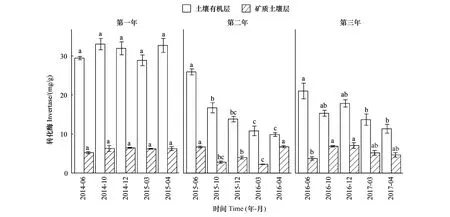
圖2 土壤有機層和礦質土壤層轉化酶活性隨關鍵時期的變化Fig.2 Change of invertase activities in both soil organic layer and mineral soil layer with critical periods不同字母表示各關鍵時期在同一年際和土層的差異顯著(P<0.05)
2.3 土壤脲酶活性動態
關鍵時期、年際、土層及其相互作用均對土壤脲酶活性產生了顯著影響(表2)。土壤有機層和礦質土壤層的脲酶活性隨關鍵時期均發生了顯著的變化,其變化趨勢也基本一致(圖3)。第一個培養年中,土壤有機層和礦質土壤層脲酶活性在雨季初期(MRS)和雨季末期(LRS)顯著高于其他關鍵時期;第二個培養年中,兩層土壤脲酶活性在雨季末期(LRS)顯著最高;第三個培養年中,土壤脲酶活性在旱季初期(EDS)顯著最低(圖3)。土壤有機層和礦質土壤層脲酶活性在第一年顯著最高,隨培養年際增加有顯著降低的變化趨勢(圖3)。
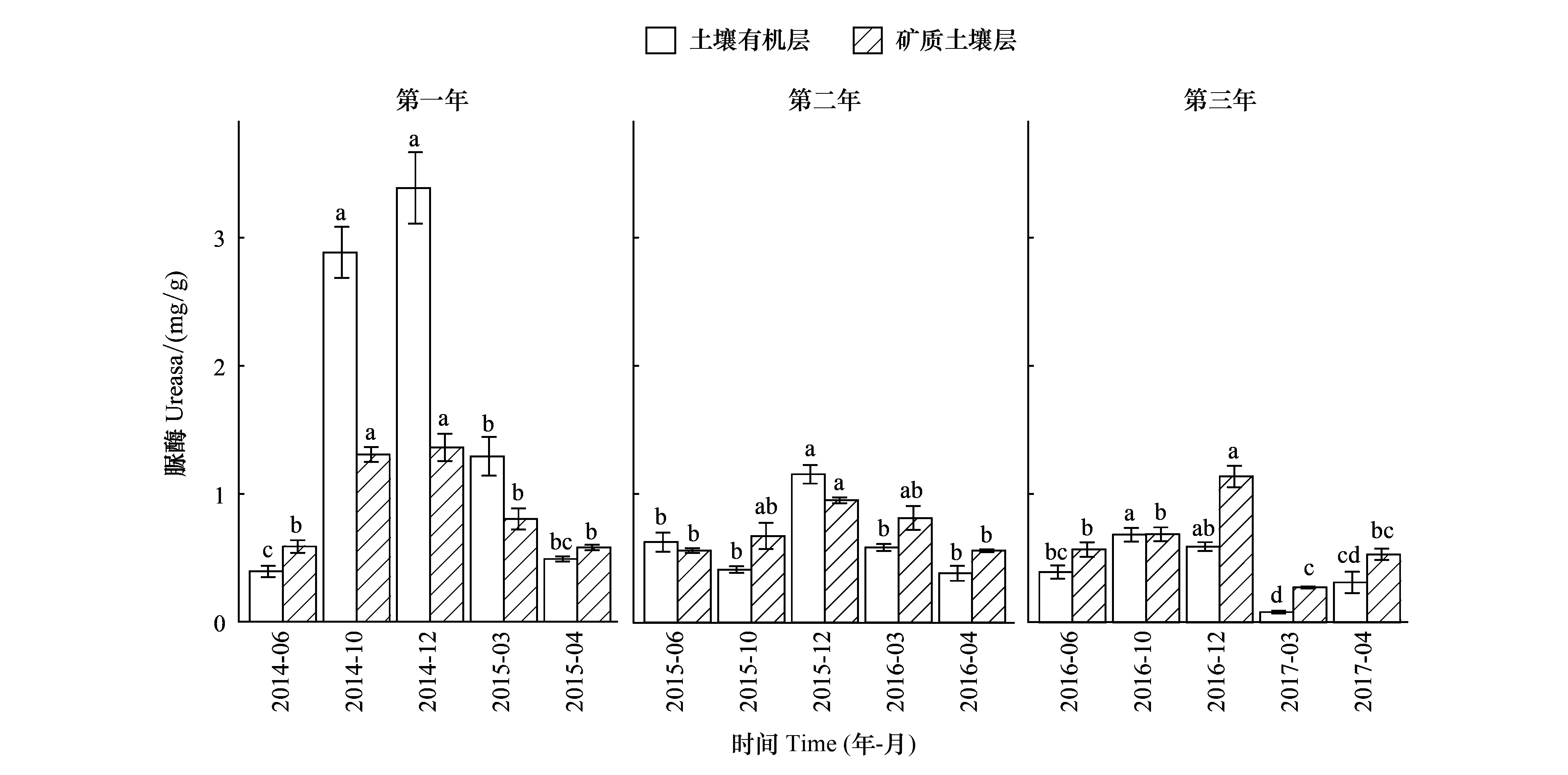
圖3 土壤有機層和礦質土壤層脲酶活性隨關鍵時期的變化Fig.3 Change of urease activities in both soil organic layer and mineral soil layer with critical periods
2.4 土壤酸性磷酸酶動態
關鍵時期、年際、土層及其相互作用均顯著影響了土壤酸性磷酸酶活性(表2)。除第三年的礦質土壤層外,土壤酸性磷酸酶活性隨關鍵時期均有顯著變化(圖4)。第一個培養年中,土壤有機層和礦質土壤層酸性磷酸酶活性均在雨季中期(MRS)顯著最高;第二個培養年中,土壤有機層和礦質土壤層酸性磷酸酶活性的顯著最低值分別出現在旱季末期(LDS)和雨季初期(ERS);第三個培養年中雨季中期(MRS)和雨季末期(LRS)的土壤有機層酸性磷酸酶活性顯著高于其他時期(圖4)。土壤有機層酸性磷酸酶活性隨培養年際增加無顯著變化,而礦質土壤層有顯著增加的趨勢(圖4)。土壤有機層酸性磷酸酶活性總體上顯著高于礦質土壤層,土壤有機層酸性磷酸酶活性的變化幅度也略大于礦質土壤層(圖4)。

圖4 土壤有機層和礦質土壤層酸性磷酸酶活性隨關鍵時期的變化Fig.4 Change of acid phosphatase activities in both soil organic layer and mineral soil layer with critical periods
2.5 土壤酶活性動態的驅動因子
土壤含水量、有機碳和全氮對土壤有機層轉化酶活性具有顯著的積極影響,土壤全磷則具有顯著的消極影響(P<0.05)(圖5)。而在礦質土壤層中,土壤微生物量碳、真菌含量、革蘭氏陰性菌含量、有機碳和革蘭氏陽性菌含量對土壤轉化酶具有顯著的積極影響(P<0.01),土壤全磷則具有顯著的消極影響(P<0.05)(圖5)。土壤含水量、微生物量氮、真菌含量、微生物量碳和革蘭氏陽性菌含量對土壤有機層脲酶活性具有顯著的正向調節作用(P<0.05),土壤全磷則具有顯著的負向調節作用(P<0.01)(圖5)。土壤含水量、真菌含量、有機碳和微生物量氮對礦質土壤層脲酶活性具有顯著的積極影響,土壤全磷則具有顯著的負向調節作用(P<0.01)(圖5)。土壤微生物量氮、降水量、土壤溫度、大氣溫度和有機碳對土壤有機層酸性磷酸酶活性具有顯著的正效應(P<0.01),而在礦質土壤層中,土壤全磷、酸堿度和微生物量氮對其具有顯著的正效應(P<0.01)(圖5)。此外,研究結果還表明,土壤含水量和土壤微生物量碳分別是土壤有機層和礦質土壤層轉化酶活性最重要的影響因素;土壤有機層和礦質土壤層脲酶活性最重要的影響因素均為土壤含水量;土壤微生物量氮和全磷分別是土壤有機層和礦質土壤層酸性磷酸酶活性最重要的影響因素。
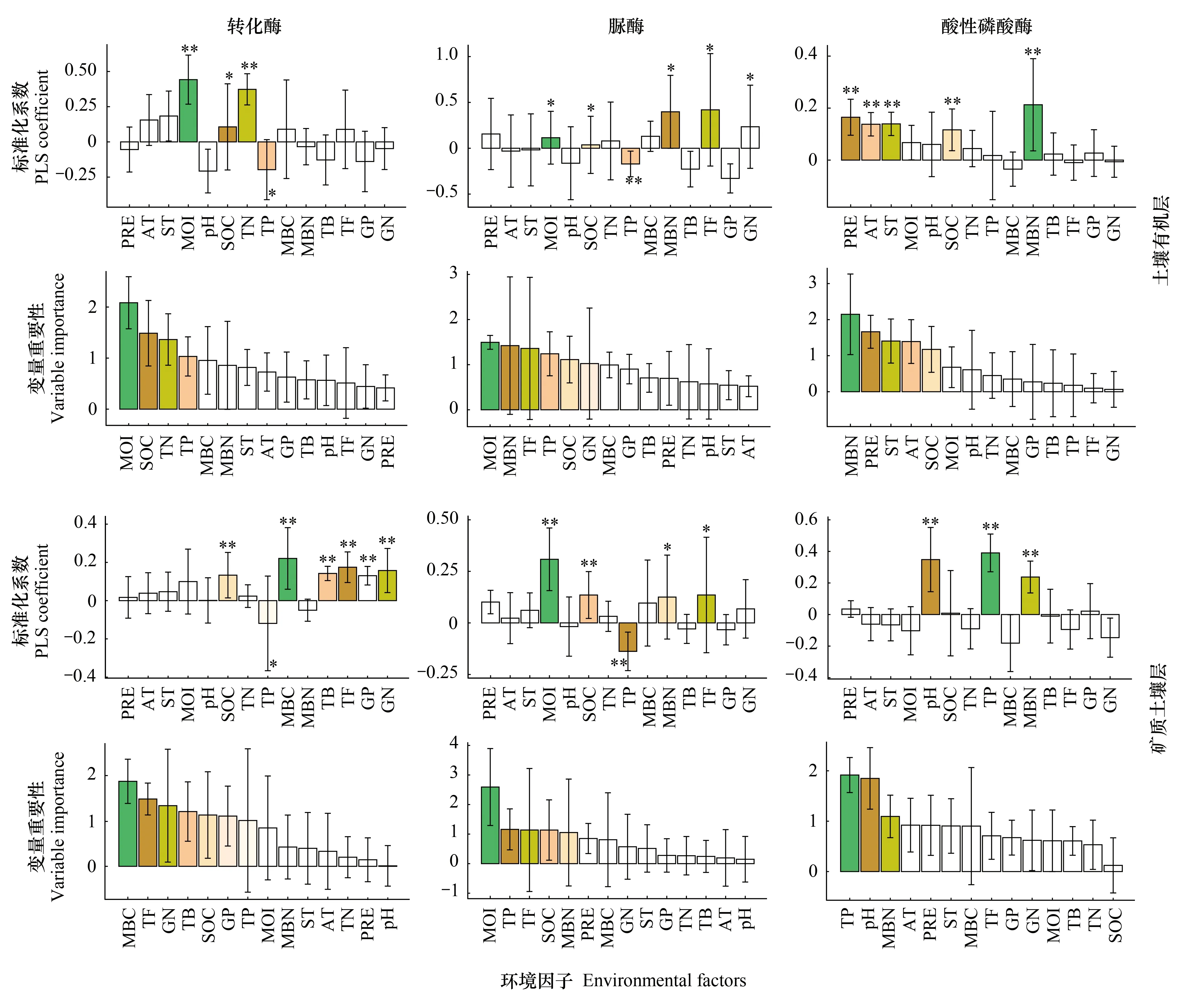
圖5 環境因子對土壤有機層和礦質土壤層酶活性影響的偏最小二乘法(PLS)分析結果Fig.5 Results of partial least squares (PLS) analysis testing for the effects of environmental factors on the soil enzyme activity in soil organic layer and mineral soil layer偏最小二乘法標準化系數顯示了每個環境因子對土壤酶活性的影響方向和大小,大于1的變量重要性表示其影響顯著;PRE:降水量 Precipitation; AT:大氣均溫 Mean air temperature;ST:土壤均溫 Mean soil temperature;MOI:土壤含水量 Soil moisture; SOC:土壤有機碳 Soil organic carbon;TN:全氮 Total nitrogen;TP:全磷 Total phosphorus;MBC:土壤微生物量碳 Microbial biomass carbon;MBN:土壤微生物量氮 Microbial biomass nitrogen;TB:細菌PLFAs含量 Bacteria PLFAs;TF:真菌PLFAs含量 Fungi PLFAs;GP:革蘭氏陽性菌PLFAs含量 Gram-positive bacteria PLFAs;GN:革蘭氏陰性菌PLFAs含量 Gram-negative bacteria PLFAs
3 討論
土壤酶活性是環境變化的敏感生物學指標,受到環境因子的綜合調控[2];因而環境因子的季節性差異通常會導致土壤酶活性的季節性變化[13,30—31]。本研究表明,土壤有機層和礦質土壤層酶活性均隨關鍵時期顯著變化,總體上表現為雨季大于旱季。這與Fan等[32]的研究結果基本一致,他們的研究發現除脲酶以外,轉化酶、酸性磷酸酶和纖維素酶等五種酶在雨季活性高于旱季,并認為濕度的增加刺激了微生物和植物根部分泌土壤酶。同時,有研究表明,土壤含水量是主導季節性降水明顯區域土壤酶活性動態的關鍵因子[33—34]。本研究中,多種環境因子對土壤酶活性具有顯著影響,但土壤含水量和微生物生物量是驅動土壤酶活性隨關鍵時期發生變化的最關鍵因素。這主要歸因于潮濕的環境有利于微生物的生理活動[35—36],提高微生物同化的效率并延長微生物的活動周期[37],導致較高的微生物生物量分泌更多的酶[38—40];反之,干旱則會顯著抑制土壤酶的活性[41—42]。本研究中,馬尾松人工林位于亞熱帶季風區,具有較為明顯的干濕季交替特征,易引起土壤含水量和微生物生物量的劇烈變化,最終導致不同關鍵時期土壤酶活性的劇烈變化。
與此同時,土壤有機層轉化酶和脲酶活性隨培養年限的增加均有顯著降低的趨勢。可能的原因是:土柱培養期間,去除新鮮凋落物減少了有機質輸入,并改變了土壤水熱條件、底物有效性和微生物量,從而降低了相關土壤酶活性[43—45]。一方面,去除新鮮凋落物增加了土壤有機層水分蒸發,顯著降低了土壤含水量,從而強化了土壤含水量對土壤酶活性的正向調節作用。另一方面,去除新鮮凋落物阻斷了土壤碳氮的主要來源,致使土壤轉化酶和脲酶活性受底物限制而顯著降低[46]。然而,不同于土壤有機層轉化酶的變化特征,礦質土壤層轉化酶活性隨年際的變化幅度相對穩定。這主要是由于礦質土壤層土壤含水量、有機碳及養分含量顯著低于土壤有機層,限制了土壤微生物分泌土壤酶[31,47],使土壤酶活性長期維持在較低水平。同時,礦質土壤層生化特性對環境變化的響應比土壤有機層更遲鈍[6],使其具有較強的穩定性并導致土壤酶活性的變化幅度明顯小于土壤有機層。此外,本研究表明,礦質土壤層酸性磷酸酶活性隨培養年限的增加呈現出增加趨勢,且土壤全磷是其變化最主要調控因素。這可能是因為酸性磷酸酶主要參與土壤磷的生物化學過程,而土壤磷元素主要受母巖和土壤淋溶過程的影響[24,48—49]。
總體而言,土壤酶活性隨關鍵時期和年際的變化受到土壤層次和酶種類的控制,主要與環境因子對土壤酶活性的影響程度的顯著差異有關。偏最小二乘分析結果表明,土壤含水量、微生物量和底物含量對土壤酶活性具有顯著影響。其中,土壤含水量和土壤微生物量碳分別是土壤有機層和礦質土壤層轉化酶活性最重要的影響因素;土壤有機層和礦質土壤層脲酶活性最重要的影響因素均為土壤含水量;土壤微生物量氮和全磷分別是土壤有機層和礦質土壤層酸性磷酸酶活性最重要的影響因素。此外,土壤微生物各類群僅在礦質土壤層對土壤轉化酶活性具有顯著的正效應,這與土壤微生物各類群組成與多數土壤酶具有顯著相關性的研究結果并不完全一致[50—51]。推測土壤微生物類群對土壤酶的調控作用可能被土壤水分或總生物量等因子限制,但相關機制有待進一步研究。同時,本研究沒有發現溫度對土壤酶活性的顯著影響,也與溫度顯著影響土壤酶活性的研究結果不一致[52—53],這可能是因為亞熱帶地區土壤酶活性的溫度敏感性較低[54]。值得注意的是,本研究只發現土壤pH與礦質土壤層酸性磷酸酶活性顯著相關,這與土壤pH是影響土壤酶活性的重要因素的研究結果不一致[55—57]。這可能是因為微環境尺度上土壤pH季節和年際變化小,為土壤酶提供了相對穩定的酸堿環境,對土壤酶活性的影響被削弱;另一方面是因為土壤pH是土壤酶活性空間分異的主要驅動因子[4],其對土壤酶活性空間分布的影響要強于對時間動態的影響。
4 結論
馬尾松人工林土壤酶活性隨關鍵時期和年際的變化顯著,其變化幅度和差異顯著性受土壤層次和酶的種類的限制。土壤有機層轉化酶和酸性磷酸酶活性顯著高于礦質土壤層。土壤有機層三種酶活性隨關鍵時期的變化幅度和隨年際的變化幅度均高于礦質土壤層。土壤含水量、土壤微生物量和底物含量是馬尾松人工林土壤酶活性的主要調控因子。其中,土壤含水量和土壤微生物量碳分別是土壤有機層和礦質土壤層轉化酶活性最重要的影響因素;土壤有機層和礦質土壤層脲酶活性最重要的影響因素均為土壤含水量;土壤微生物量氮和全磷分別是土壤有機層和礦質土壤層酸性磷酸酶活性最重要的影響因素。
