喀斯特退化天坑不同坡向植物群落系統發育與功能性狀結構
2022-10-27 03:14:40劉櫞錳張永永孫曉瑞
生態學報 2022年19期
稅 偉,馮 潔,李 慧,江 聰,孫 祥,劉櫞錳,張永永,孫曉瑞
1 福州大學環境與安全工程學院, 福州 350116 2 福州大學空間數據挖掘與信息共享教育部重點實驗室, 福州 350116 3 北京大學城市與環境學院, 北京 100871
喀斯特天坑是一種具有垂直峭壁的圈閉化“負地形”,坑內形成異于地表的局域小生境,為某些植物的生長繁殖提供了“避難所”,坑內形成的獨特地下森林,是一座天然的物種保護庫[1—2]。由于天坑不同區域受垂直峭壁影響的程度不同,內部局域環境差異大,植物群落分異明顯。在原生天坑坑壁梯度上存在明顯的生態交錯帶[3],在退化天坑底部的草地群落呈圈層分布,群落物種豐富度和多樣性呈“逆洼地效應”[4],其倒石坡喬灌植物物種多樣性高于坑外[5],不同坡向植物群落類型分異明顯,陽坡以殼斗科等耐旱型植物群落為主要建群種,而陰坡隨著演替的進行,殼斗科等耐旱型植物優勢逐漸減弱[6]。目前天坑植物群落的研究多集中在物種多樣性特征、植物群落的空間特征、不同環境梯度下植物功能性狀特征等,而結合系統發育和功能性狀研究天坑植物群落構建的則鮮有。
群落構建機制受多種條件影響[7],不同群落受地方生物因素和非生物因素影響,導致群落類型和演替速度的不同,其系統發育結構也發生改變[8]。新幾內亞低地雨林的次生林系統發育結構聚集,原始林系統發育結構發散,表明隨著演替的進行群落構建的主導因素由生境過濾轉向生物間的相互作用[9],然而哥斯達黎加東北部演替早期的熱帶低地潮濕雨林群落的系統發育結構發散,競爭排斥影響較大[10]。可見群落的系統發育結構受研究區域的環境因素影響而不同。植物群落構建在小尺度上的研究,地形和土壤因子是非常重要的影響因素,坡向是小尺度研究中的一個典型梯度,對植物群落構建有重要意義[11—12]。不同坡向其光照強度、土壤溫濕度、土壤營養物質含量差異大[13—14],會影響植物群落的物種組成、物種多樣性、功能性狀多樣性等[15],導致其群落構建機制可能會發生變化。隨著Blomberg等[16]提出系統發育信號檢驗的發展,系統發育在群落構建中的研究日漸增多。系統發育的方法可以很好地構建微域的物種庫,揭示某地區的物種進化歷史、過去物種的生態位進化以及當代物種的共存機制[17]。在天坑圈閉化生境中,發育了多個坡向的倒石坡,具有獨特生境的天坑內部又形成了異質性的坡向小生境,因此對坡向植物多樣性維持機制的研究,探討環境作用和生物作用對不同坡向植物群落構建的影響,有助于揭示天坑生物多樣性保護庫價值,為喀斯特地區植被恢復提供依據。
功能性狀是植物與環境之間長期相互影響的結果[6],能夠指示環境變化,反映植物生態策略,對揭示植物群落構建機制有重要作用[18—19],本文以云南沾益深陷塘天坑為例,結合植物功能性狀和系統發育的方法研究植物群落構建,分析天坑坡向植物群落的功能性狀是否具有顯著的系統發育信號,通過不同坡向的植物群落系統發育結構和功能性狀結構,量化環境過濾、競爭排斥和隨機過程在天坑坡向植物群落構建中的作用,為天坑植物多樣性的保護提供科學的參考。
1 材料和方法
1.1 區域概況和樣方設置
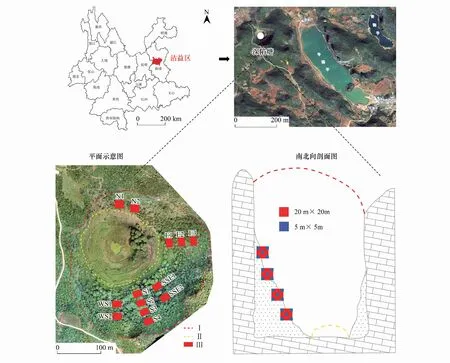
圖1 天坑區位圖及樣方布設示意圖Fig.1 Location map of Tiankeng and schematic diagram of plot layoutN: 北 North; E: 東 East; SSE: 東南偏南 south-southeast.; S: 南 South; WS: 西南 southwest; Ⅰ: 天坑上部邊界Upper boundary of tiankeng; Ⅱ: 坑坡下部邊界Lower boundary of tiankeng slope; Ⅲ: 天坑喬木樣方Arbor quadrat in tiankeng
沾益天坑群位于云南省曲靖市(圖1),地理坐標在25°35′—25°57′N, 103°29′—103°39′E之間,其中包含原生天坑和退化天坑,具有豐富的物種多樣性[20]。該地區屬于亞熱帶高原季風氣候,晝夜溫差較大,年均溫在13.8—14℃,太陽輻射能年總量為518.2 kJ/cm2,天坑群內土壤主要有紅壤、黃棕壤、紫色土及石灰土等。經過對云南喀斯特天坑群的調查,本研究以典型的中度退化天坑——深陷塘天坑為研究對象。深陷塘坑底海拔2028—2031 m,坑口長徑421.9 m,短徑348.7 m,坑深148.7 m[5]。西側為垂直峭壁,北側為陡峭退化坑壁,是天坑的入口,植被以稀疏耐旱的灌木為主;東側上部為垂直峭壁,下部為倒石坡,倒石坡上分布著喬灌相間的植被;南側是坑內最大的倒石坡,植被以喬木為主。南側倒石坡是坑內陰坡,生境條件較好,東坡次之,北坡作為陽坡,生境條件最差。根據坑內坡長和植物生長分布的實際情況,在南側、西南側、南南東側、東側和北側共布設13個20 m×20 m的喬木樣方,在每個喬木樣方內按照“梅花型”設置5個5 m×5 m的灌木樣方,共計65個灌木樣方,分別調查記錄每個樣方內植物的物種名、株高、胸徑、多度,利用GPS記錄經緯度和海拔數據。物種多樣性對植物群落有重要影響[21],通過計算分析可得不同坡向植物群落α多樣性,其中Margalef豐富度為南坡>東坡>北坡,且差異顯著,而不同坡向植物群落的物種多樣性和均勻度差異不顯著。
1.2 功能性狀的采集和測定
根據全球植物功能性狀標準化測量手冊[22],在樣方內使用高枝剪采摘每個物種不同植株不同方向的長勢良好的3—5片葉子。使用精度為0.02 mm的電子數顯卡尺測葉子主脈一側0.25 cm處,記錄三組數據求平均值;使用葉面積儀記錄每一片葉子的葉長(LL)、葉寬(LW)、長寬比(LWR)和葉面積(LA);鮮葉采用精度為0.001 g電子天平稱量鮮重,葉片帶回實驗室后放入設置為80℃的烘干箱48小時至恒重,然后稱量其干重。比葉面積(SLA)為葉面積與葉干重的比值;葉干物質含量(LDMC)為葉干重與葉鮮重的比值。
1.3 土壤因子的測定
土壤理化性質指標依托于福建省農科院土壤肥料研究所測定。土壤含水量(SWC, %)采用烘干法,用環刀采集深度為0—20 cm土壤樣品,稱量其濕重M,在溫度設置為105℃的烘箱中6—8 h,烘干至恒重Ms,計算土壤含水量SWC=(M-Ms)/M;用土壤酸度計測定pH,用重鉻鉀氧化法測定土壤有機質(SOM),堿解擴散法測定土壤堿解氮(AN),土壤全氮(Total nitrogen, TN)的測定采用半微量開氏法,用鉬銻抗比色法測定全磷(TP)和有效磷(AP),用火焰光度計法測定全鉀(TK)和速效鉀(AK)。
1.4 功能性狀的系統發育信號檢測
本文根據Blomberg等[16]提出的K值檢驗法分析系統發育信號。K=1定義為Brownian motion的進化模型;K>1,表明功能性狀具有比Brownian motion模型進化更強的系統發育信號;反之,K<1,表明功能性狀的系統發育信號較弱。通過實際K值與零模型的比較,進而判斷功能性狀系統發育信號的顯著性。當系統發育樹分支末端物種隨機置換999次生成的零模型K值小于實際K值(P<0.05),功能性狀具有顯著的系統發育信號,否則說明系統發育信號不顯著[15]。
1.5 群落系統發育和功能性狀結構
系統發育樹的構建是采用物種的科屬信息經過R語言的“plantlist”程序包處理,輸入網頁版phylomatic軟件中,得到物種的系統發育的拓撲結構,該結構是以被子植物分類系統Ⅲ(APGⅢ, Angiosperm Phylogeny Grup Ⅲ)為基礎構建。然后經過phylocom軟件的BLADJ算法,結合分子及化石定年數據擬合系統發育拓撲結構的分支結構,采用R語言“picante”程序包計算得到系統發育樹。本文研究中的凈親緣關系指數(Net relatedness index, NRI)是群落中所有物種的平均系統發育距離(Mean pairwise distance, MPD)的標準效應值,最近親緣關系指數(Nearest taxon index, NTI)是平均最近相鄰譜系距離(Mean nearest taxon distance, MNTD)的標準效應值。根據調查樣方群落得到實際的MPD或MNTD,把物種隨機置換999次構建零模型,得到隨機狀態下的MPD或MNTD,經過標準化得到NRI或NTI[23—24]。
采用平均配對性狀距離(Mean pairwise trait distance, traitMPD)和平均最近相鄰性狀距離(Mean nearest taxon trait distance, traitMNTD)計算功能性狀結構。TraitSESMPD(Standardized mean pairwise trait distance)和traitSESMNT(Standardized mean nearest taxon trait distance)是基于功能性狀聚類樹計算求得。
系統發育指數(NRI和NTI)和功能性狀結構指數(traitSESMPD和traitSESMNTD)等于0時,表示譜系結構和功能性狀結構自由擴散;指數大于0時,表示譜系結構和功能性狀結構聚集;指數小于0時,表示譜系結構和功能性狀結構發散。
式中,MPDobs、MNTDobs、traitMPDobs、traitMNTDobs是實際觀測值;MPDrand、MNTDrand、traitMPDrand、traitMNTDrand是零模型下隨機零群落MPD、MNTD、traitMPD、traitMNTD的值;mean(MPDrand)、mean(MNTDrand)、mean(traitMPDrand)、mean(traitMNTDrand)是通過零模型計算的平均值;sd(MPDrand)、sd(MNTDrand)、sd(traitMPDrand)、sd(traitMNTDrand)是零模型下隨機零群落的各指數的標準差。
1.6 數據處理
植物功能性狀的種內變異是物種性狀的標準差與平均值的比值。重要值是用綜合性數量指標來反映植物物種在群落中的相對重要性[25],本文以重要值的大小作為群落主要優勢物種的測度指標[26—27]。喬木層重要值=(相對密度+相對頻度+相對顯著度)/3,灌木層重要值=(相對密度+相對頻度+相對顯著度)/3。根據單因素方差分析判斷不同坡向的系統發育指數、功能性狀結構指數的顯著性差異。數據整理和多樣性指數計算在Excel 2016中進行,系統發育和功能性狀結構指數的計算在R 4.0.2中實現,相關制圖均在R 4.0.2和Origin 2018中繪制。
2 結果與分析
2.1 不同坡向土壤環境差異
深陷塘天坑的土壤有機質、有效磷和土壤含水量從北坡到南坡逐漸增大,土壤pH從北坡到南坡逐漸減小。全氮含量在北坡(2.03 g/kg)最低,而東坡和南坡差異不大;土壤堿解氮含量東坡(185.2 mg/kg)最高,北坡(105.3 mg/kg)最低;土壤全磷在北坡含量(0.67 g/kg)最高,東坡和南坡相差不大;土壤全鉀在南坡含量(14.33 g/kg)最高,東坡含量(5.10 g/kg)最低;土壤速效鉀在北坡含量(139.50 mg/kg)最高,東坡含量(87.93 mg/kg)最低(表1)。從北坡—東坡—南坡,土壤含水量及土壤營養物質含量有逐漸增大的趨勢,東坡土壤含水量和土壤營養物質介于北坡和南坡之間,且變異系數較大,可能是因為東坡處于陰坡和陽坡的交界處,造成不同坡位樣方的土壤環境差異較大。
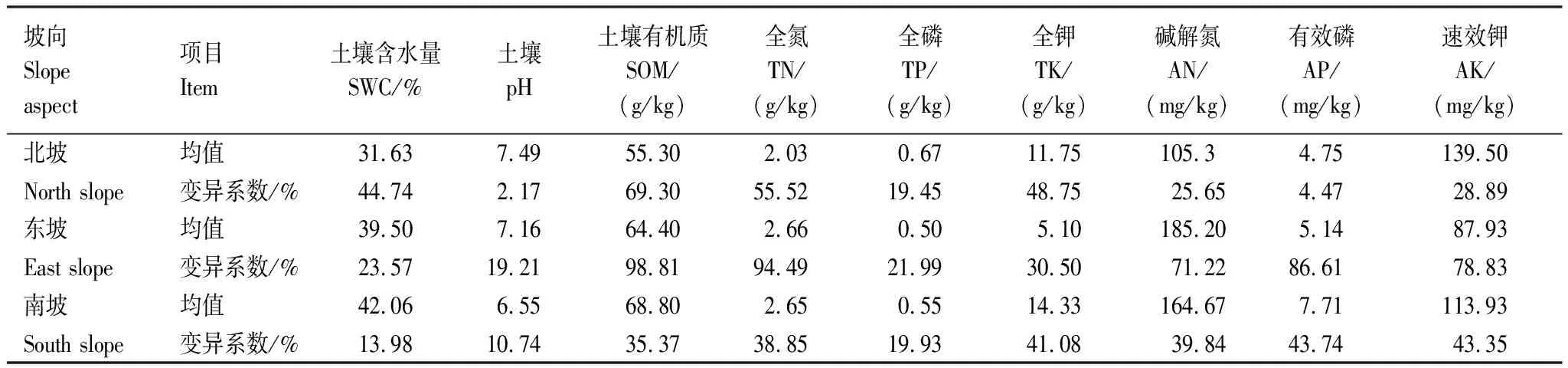
表1 深陷塘不同坡向土壤環境因子
2.2 不同坡向植物群落組成
本研究共調查26科40屬47種物種(圖2),其中北坡15種,東坡20種,南坡45種。本文取各個坡向重要值最大的前三個物種來表征該坡向的優勢種,因此北坡優勢種有黃背櫟、長圓葉梾木(Swidaoblonga)、球核莢蒾(Viburnumpropinquum),東坡優勢種有黃背櫟、栓皮櫟(Quercusvariabilis)、清香木 (Pistaciaweinmannifolia),南坡優勢種有頭狀四照花 (Dendrobenthamiacapitata)、青岡、構樹(Broussonetiapapyrifera)(表2)。
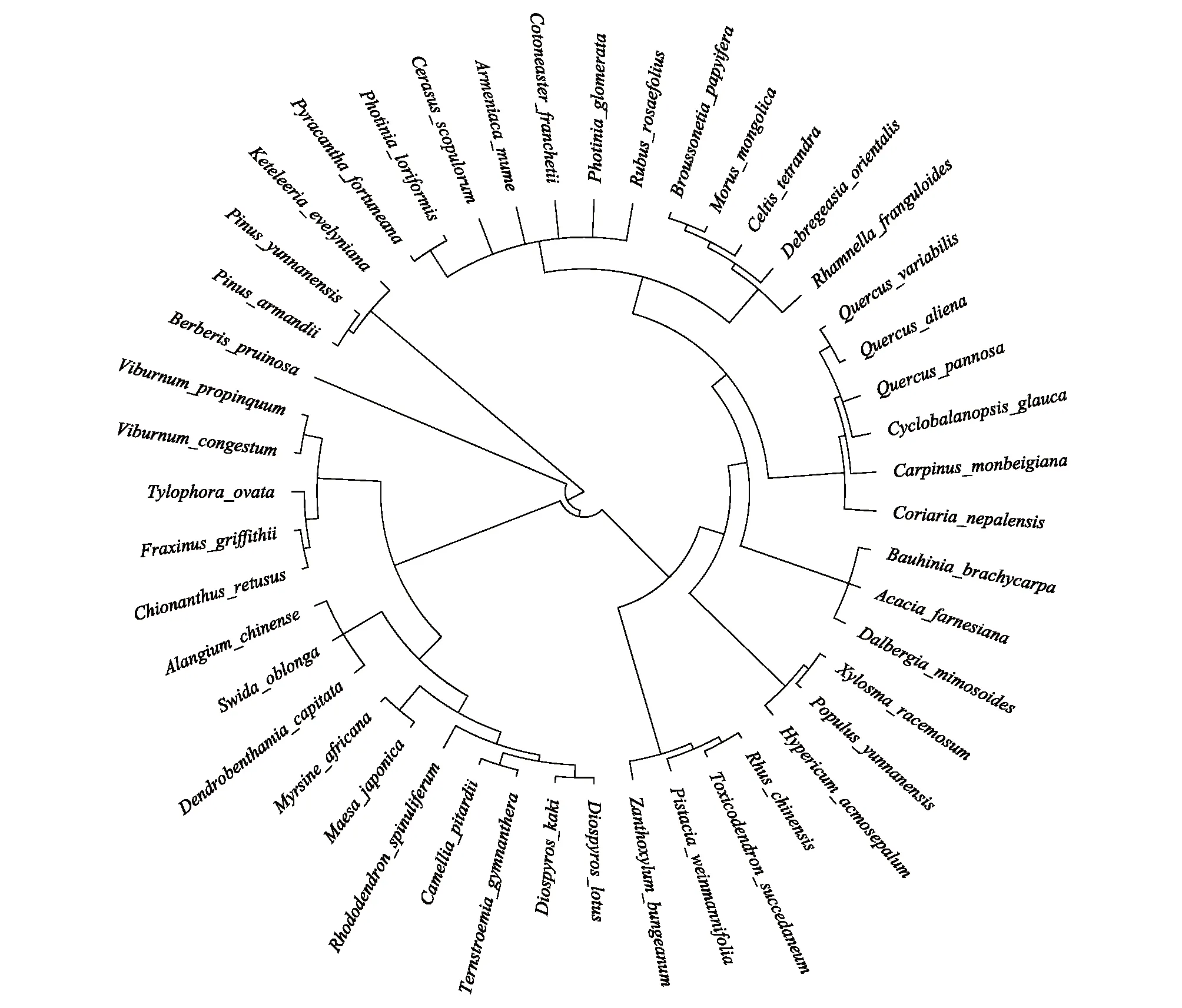
圖2 系統發育樹Fig.2 Phylogenetic treeBerberis pruinosa: 粉葉小檗; Toxicodendron succedaneum: 野漆; Rhus chinensis: 鹽膚木; Pistacia weinmanniifolia: 清香木; Zanthoxylum bungeanum: 花椒; Hypericum acmosepalum: 尖萼金絲桃; Populus yunnanensis: 滇楊; Xylosma racemosum: 柞木; Dalbergia mimosoides: 象鼻藤; Acacia farnesiana: 金合歡; Bauhinia brachycarpa: 鞍葉羊蹄甲; Coriaria nepalensis: 馬桑; Carpinus monbeigiana: 云南鵝耳櫪; Quercus aliena: 槲櫟; Quercus variabilis: 栓皮櫟; Cyclobalanopsis glauca: 青岡; Quercus pannosa: 黃背櫟; Rhamnella franguloides: 貓乳; Celtis tetrandra: 四蕊樸; Morus mongolica: 蒙桑; Broussonetia papyrifera: 構樹; Debregeasia orientalis: 水麻; Photinia loriformis: 帶葉石楠; Pyracantha fortuneana: 火棘; Cotoneaster franchetii: 西南栒子; Armeniaca mume: 梅; Photinia glomerata: 球花石楠; Cerasus scopulorum: 崖櫻桃; Fraxinus griffithii: 光蠟樹; Chionanthus retusus: 流蘇樹; Tylophora ovata: 娃兒藤;Rubus rosaefolius: 空心泡; Viburnum propinquum: 球核莢蒾; Viburnum congestum: 密花莢蒾; Rhododendron spinuliferum: 爆杖花; Camellia pitardii: 西南紅山茶; Ternstroemia gymnanthera: 厚皮香; Diospyros kaki: 野柿; Diospyros lotus: 君遷子; Myrsine africana: 鐵仔; Maesa japonica: 杜莖山; Alangium chinense: 八角楓; Swida oblonga: 長圓葉梾木; Dendrobenthamia capitata: 頭狀四照花; Pinus yunnanensis: 云南松; Pinus armandii: 華山松; Keteleeria evelyniana: 云南油杉
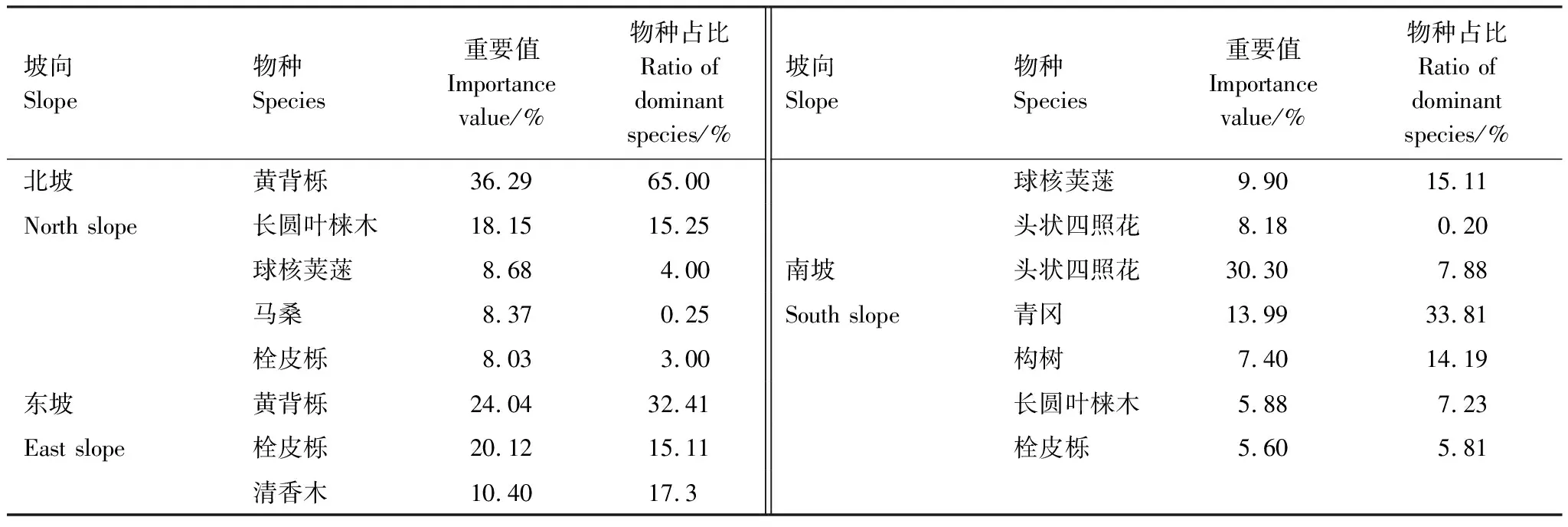
表2 不同坡向重要值前5的物種
2.3 退化天坑不同坡向優勢種功能性狀變異特征
深陷塘各坡向優勢種性狀總體變異程度中等,不同性狀之間存在差異。從表3來看,在北坡,黃背櫟功能性狀變異程度最大的是葉面積,最小是長寬比,而長圓葉梾木葉厚度變異程度最大,比葉面積變異程度最小,球核莢蒾也是葉面積變異程度最大,但葉寬變異程度最小;東坡黃背櫟和栓皮櫟都是葉面積變異程度最大,葉長最小,清香木比葉面積變異程度最大,葉寬最小;南坡頭照四狀花和構樹都是葉干重變異程度最大,葉長最小,而青岡葉干物質含量變異程度最大,葉寬最小。三個坡向的優勢種中,東坡清香木葉面積變異程度最大(54.26%),而變異程度最小(8.66%)的則是南坡青岡的葉寬。整體上,南坡優勢物種變異程度最大,東坡次之,北坡優勢物種變異程度最小。
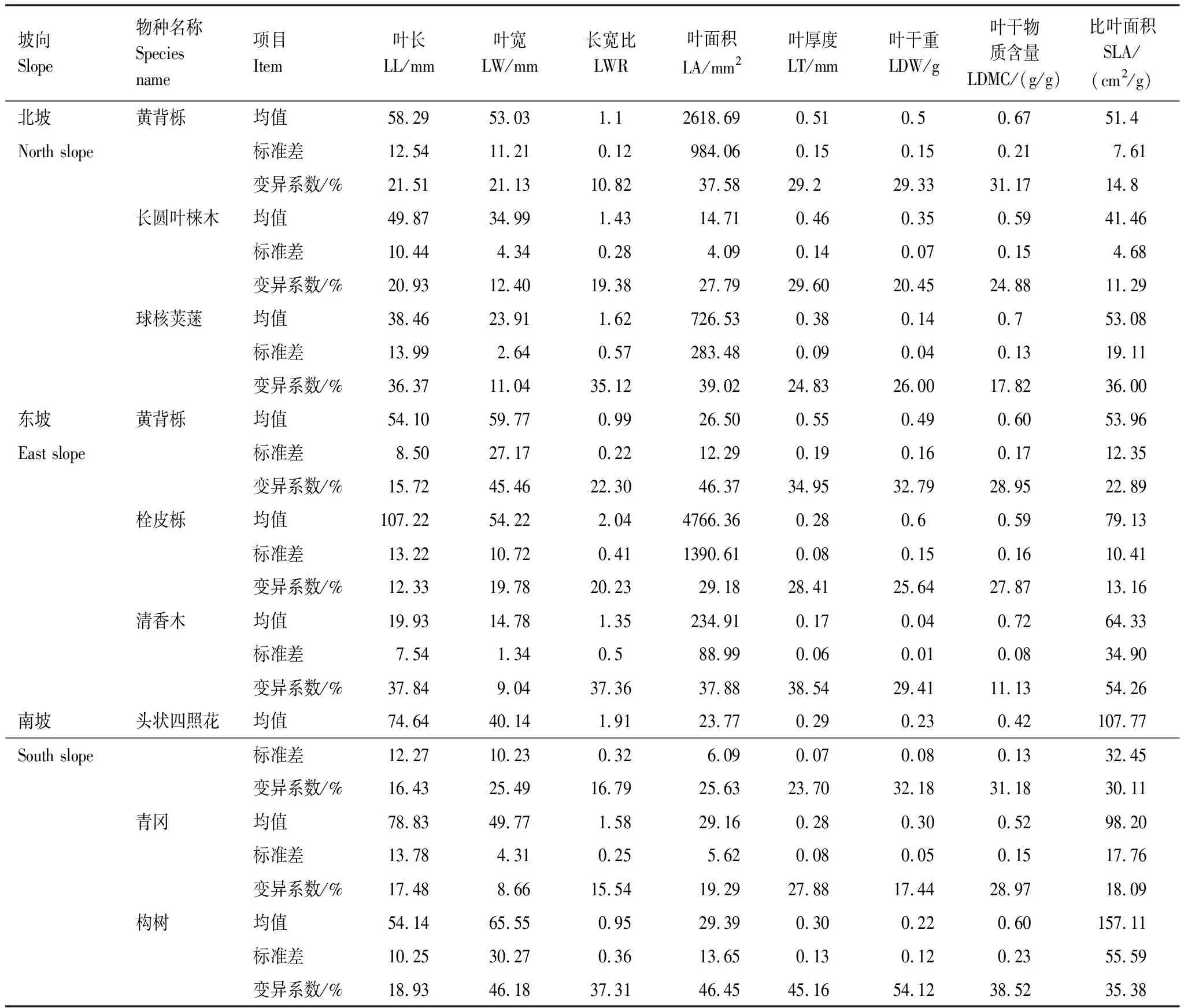
表3 深陷塘不同坡向優勢物種功能性狀變異特征
2.4 退化天坑不同坡向植物功能性狀的系統發育信號檢驗
植物群落構建過程中聚集是環境過濾占主導作用,而發散主要由競爭排斥作用主導。喀斯特天坑深陷塘坡向植物群落的葉長、葉面積、葉干重、葉干物質含量、最大樹高的K值小于1,說明功能性狀表現出的系統發育信號強度弱,這5種功能性狀都沒有較強的系統發育保守性,性狀在進化過程中受生境影響更大;而葉寬、比葉面積、長寬比、葉厚度的K值大于1,說明功能性狀表現出的系統發育信號強,受遺傳因素影響顯著(P<0.05)(表4),表明譜系距離近的物種,功能性狀的相似度高。
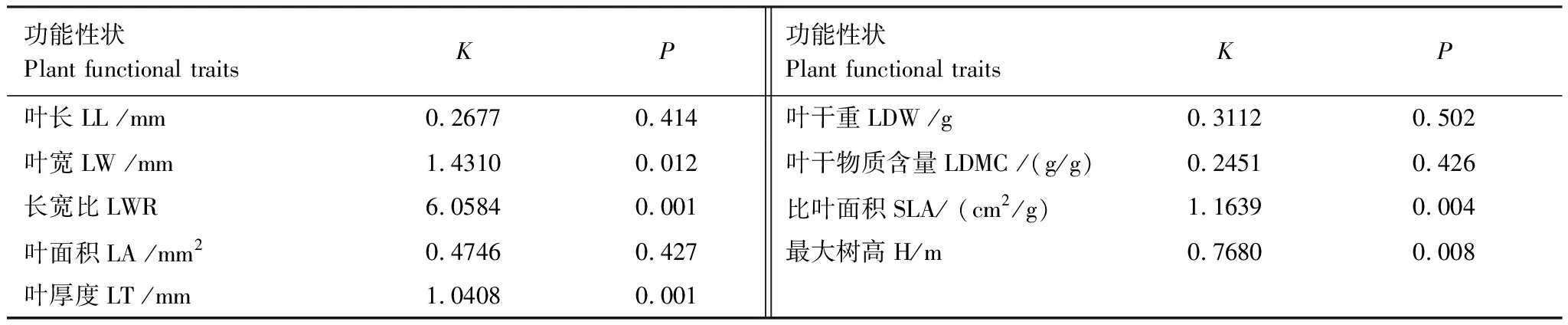
表4 深陷塘坡向植物功能性狀系統發育信號檢驗
2.5 退化天坑不同坡向群落的系統發育和功能性狀結構的差異
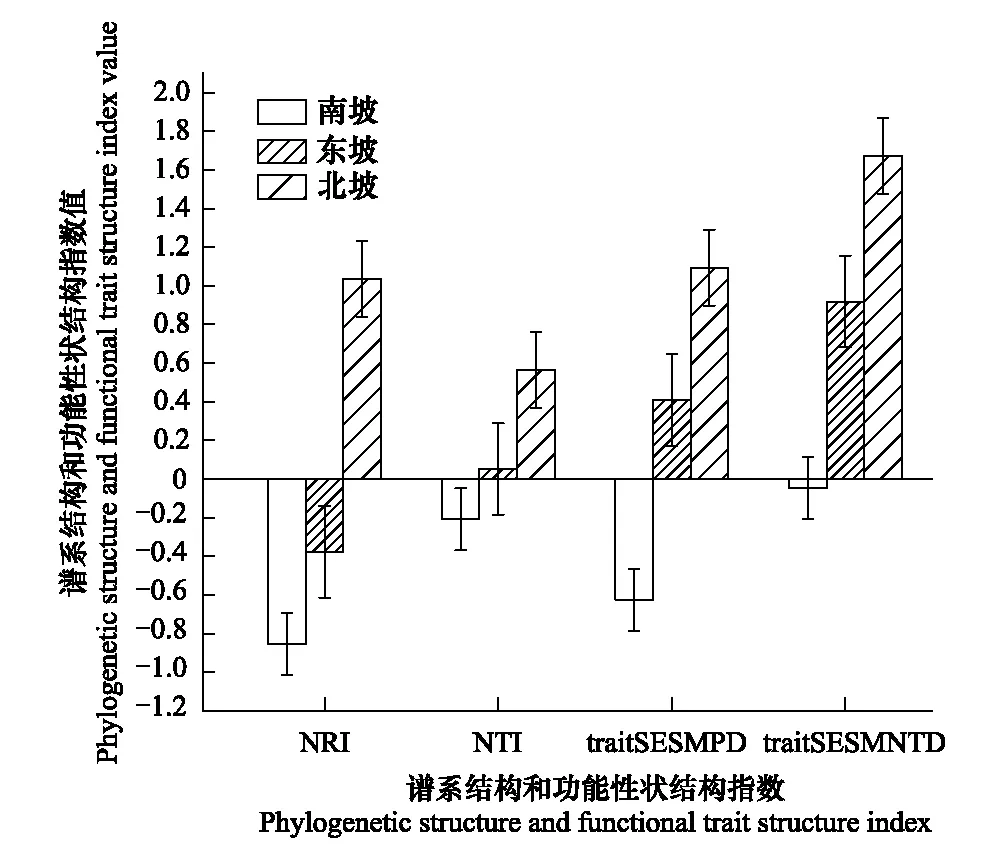
圖3 不同坡向的譜系結構和功能性狀結構 Fig.3 Phylogenetic structures and functional structures of different slope NRI: 凈種間親緣關系指數 Net relatedness index; NTI: 最近種間親緣關系指數 Nearest taxon index; trait SESMPD: 標準化種間平均配對性狀距離 Standardized mean pairwise trait distance; trait SESMNTD: 標準化種間平均最近相鄰性狀距離 Standardized mean nearest taxon trait distance
喀斯特天坑深陷塘北坡植物群落NRI、NTI和traitSESMPD、traitSESMNTD均大于0,表明系統發育結構和功能性狀結構均聚集。而南坡植物群落的NRI、NTI指數和traitSESMPD、traitSESMNTD指數均小于0,表明功能性狀結構和功能性狀結構均發散。東坡植物群落系統發育兩指數結果不一致,NRI指數小于0,說明系統發育結構發散;NTI指數大于0,表明系統發育結構聚集;功能性狀結構指數traitSESMPD和traitSESMNTD均是大于0,說明功能性狀結構聚集。NRI、NTI和traitSESMPD、traitSESMNTD沿著坡向變化(北坡-東坡-南坡)存在相同的變化趨勢,即隨著坡向的變化系統發育結構和功能性狀結構都是從集聚逐漸轉變為發散(圖3)。
3 討論
3.1 天坑植物群落系統發育與功能性狀結構指數隨坡向的變化特征
深陷塘天坑受西面垂直峭壁的遮擋影響,不同方位日輻射量不同,坑內濕熱環境異于地表,坑內不同坡向生境中植物群落類型不同,其群落構建機制過程存在差異。江聰等[28]在研究紫莖澤蘭(Eupatoriumadenophorum)入侵巴家陷塘天坑時發現,紫莖澤蘭在坑坡灌草地較為集中,主要分布在西南坡,而東北坡基本沒有分布,從西南坡到東北坡,植物群落由陽生植物演替為陰生植物。本研究表明,深陷塘各坡向系統發育指數和功能性狀結構指數有相同的變化趨勢,即從北坡到南坡,系統發育指數減小,功能性狀結構指數也減小。由圖3可知,北坡的植物群落的系統發育結構和功能性狀結構均為聚集,而南坡植物群落的系統發育結構和功能性狀結構均為發散,且從北坡到南坡發散程度逐漸上升。在東坡,兩種功能性狀結構指數均為聚集,而兩種系統發育指數結果不同,NRI指數表明東坡系統發育結構發散,NTI指數表明東坡系統發育結構聚集。車應弟等[29]在分析亞高寒草甸群落構建時發現西坡兩種系統發育指數也不一致。東坡兩種系統發育指數結果不同,可能有兩種解釋:一是兩種系統發育指數側重點不同,NRI親緣指數是基于群落整體水平的,反映的是整個系統發育樹的發育模式,而NTI親緣關系是基于系統進化樹末端的最近的系統發育關系[30];二是由于深陷塘西面是垂直峭壁,東坡屬于北坡和南坡的坡向過渡地帶,環境條件和種間競爭共同影響了該坡的群落構建。受垂直峭壁的遮擋影響,陰坡和陽坡生境分異明顯,深陷塘北坡為陽坡,土壤含水量低,土壤養分含量少,植被主要以耐旱的黃背櫟等灌木為主,環境對物種的生長影響較大,這與劉旻霞等[31]的研究結果一致,即環境過濾是陽坡群落構建的主要作用;而深陷塘南坡為陰坡,土層深,土壤含水量高,養分豐富,總生態位寬,植被以頭狀四照花和構樹等闊葉喬木為主,物種多樣性高,加劇了一些功能性狀相似的物種因生態位重疊范圍較大而引起的競爭,這與朱云云等[32]的研究結果一致,即競爭排斥是陰坡群落構建的主要作用;東坡作為北坡和南坡的過渡地帶,喬灌混雜,受環境過濾和競爭排斥的共同作用。從北坡到東坡,再到南坡的植物群落構建過程中,環境過濾作用逐漸減小,競爭排斥作用逐漸增大,植被群落由耐旱性的灌木,逐漸演替為高大的喬木,這三個坡向植物組成構成了一個群落演替系列。
3.2 天坑坡向植物群落構建中的生物過濾機制
群落構建在生物過濾和環境過濾的長期影響下,物種逐漸趨于局域環境物種庫的過程[33],其中種間競爭和種內性狀變異是生物過濾的主要生態學過程[34]。種間競爭是群落結構和植物形態的重要影響因素之一,導致物種間形成生態排序,競爭能力弱的物種逐漸從群落中淘汰,最后形成的共存物種有均等的適合度[35—36],植物群落逐漸趨于穩定。種內變異多是因為植物功能性狀的生態適應性策略,植物功能性狀的變異受環境因子(氣候、地形、土壤因子等)的影響,但功能性狀之間的相互作用也是重要因素[34]。Ackerly等[37]的研究表明由于南北坡光照、濕度等存在差異,導致不同坡向植物的種內功能性狀出現變異。本研究中,北坡是陽坡,光照強度大,土壤水分含量低,營養物質相對貧瘠,惡劣的環境使得北坡植物群落的譜系結構聚集,促進了植物功能性狀趨同進化。而南坡是陰坡,土壤水分含量高,營養物質豐富,植物群落復雜,屬于針闊混交林。林下灌叢茂密,植物群落間競爭排斥占主導作用,導致植物功能性狀趨異進化。南坡優勢種通過增大種內變異程度來緩沖物種間的競爭作用,提高南坡植物群落的穩定性。這與劉旻霞等[38]和盤遠方等[39]的研究結果一致,表明由于陰坡和陽坡的生境差異,導致坡向變化是植物功能性狀結構存在差異的主要原因。東坡是連接北坡與南坡的生態交錯帶[40],生境條件次于南坡,卻高于北坡;東坡植物的葉長、葉寬、長寬比、葉面積和葉干重有較小的種內變異,可能是因為東坡以黃背櫟和栓皮櫟為主要優勢建群種,其多度較大,種內變異程度與物種的多度有負相關關系[34],因為常見種在資源獲取的能力方面差異不大,所以種內變異程度較小。
4 結論
本文通過對深陷塘退化天坑不同坡向植物功能性狀和系統發育的研究發現:深陷塘天坑不同坡向的系統發育和功能性狀結構具有不一致性,但總體是受生態位過程主導。環境過濾是北坡物種共存的主要機制,競爭排斥是南坡物種共存的主要機制,對于東坡這種生境過渡區域,受到環境過濾和競爭排斥的共同作用。坑內植物功能性狀受遺傳因子和生境的共同作用,在不同的坡向梯度生境中,采取不同的生存策略,揭示了坡向生境對植物群落功能性狀的影響。未來可以結合土壤、光照、水分等環境因子,進一步量化環境因子在不同坡向植物群落構建中的作用,為云南喀斯特天坑植物資源保護和繁育、天坑群生態保護和恢復工作提供理論支撐。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中華詩詞(2022年6期)2022-12-31 06:41:24
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國科技論壇(2017年7期)2017-07-25 08:49:53
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
現代企業(2015年9期)2015-02-28 18:56:50
