黃海秋季大型底棲甲殼類多樣性及群落結構的初步研究
2022-10-29 06:27:48李少月張蒙生滕安康邵聰聰于子山
海洋科學 2022年9期
關鍵詞:研究
祝 琳, 李少月, 王 智, 張蒙生, 陳 晨, 滕安康, 邵聰聰, 于子山
黃海秋季大型底棲甲殼類多樣性及群落結構的初步研究
祝 琳1, 李少月2, 王 智3, 張蒙生4, 陳 晨1, 滕安康1, 邵聰聰1, 于子山1
(1. 中國海洋大學 海洋生命學院, 山東 青島 266003; 2. 國家海洋局北海環境監測中心, 山東 青島 266003; 3.廈門大學 近海海洋環境科學國家重點實驗室, 福建 廈門 361102; 4. 上海勘測設計研究院有限公司, 上海 200050)
基于2014年秋季(11月)在黃海海域所獲取的大型底棲動物及環境數據, 研究了黃海海域大型底棲甲殼類動物的種類組成、豐度、生物量、多樣性以及群落結構特征等。調查海域共發現71種大型底棲甲殼類動物, 隸屬于32科42屬, 其平均豐度和生物量分別為278 inds./m2和2.33 g/m2。相對重要性指數最高的物種為太平洋方甲漣蟲(Hart, 1930)。在23%的相似度水平上, 可以將大型底棲甲殼類劃分為6個群落。Pearson相關性結果顯示, 太平洋方甲漣蟲豐度與水深呈極顯著正相關, 與底層鹽度呈顯著正相關, 與底層溫度呈顯著負相關。大型底棲甲殼類動物的物種數占比在黃海大型底棲動物類群組成中變化不大, 都在30%左右。BOPA指數表明, 調查海域底棲生態環境整體良好, 僅黃海中部和南部部分站位可能存在中度擾動; BOPA指數能較好地評價研究海域的環境質量狀況。
黃海; 大型底棲甲殼類; 群落; BOPA指數; 環境因子
大型底棲甲殼類(以下簡稱底棲甲殼類)在海洋生態系統的能量流動和物質循環中具有重要作用, 它們大多生活在有氧和有機質豐富的沉積物表層, 通過攝食、掘穴和建管等擾動活動, 直接或間接地影響其所在的生態系統[1]。底棲甲殼類種類多, 數量大, 是優勢類群, 在海洋底棲生物中占有重要的地位[2]。例如生活于沉積物表層的端足類、等足類和漣蟲類, 個體雖小, 但因為數量大, 構成大型無脊椎動物及魚類幼體的重要餌料。底棲甲殼類生活習性相對穩定, 區域性強, 遷移能力弱[3], 對環境污染等人類活動有著持久的綜合的響應。因此, 底棲甲殼類可作為評價環境質量的指示生物。
從20世紀90年代始, 我國海洋甲殼動物的生態學研究陸續展開, 主要包括甲殼類動物資源的生物量評估、群落結構特征的分析、群落的劃分、主要經濟種的生物學特性和時空分布特征等。進入21世紀以來, 我國深入開發利用海洋資源, 海洋甲殼動物調查規模、頻次、所涉海域明顯增加, 這使得對蝦蟹類生態學的研究更為深入[4]。例如黃梓榮等[5]對南海北部陸架區甲殼類動物資源密度分布與種類組成的研究; 盧占暉等[6]對東海中部蝦類季節群落的研究; 宋海棠[7]、俞存根等[8]、陳小慶等[9]、李惠玉[10]對東海區蝦蟹類生態類群和區系特征劃分研究等等。關于黃海的甲殼類, 于海燕等[11]報道了膠州灣大型底棲甲殼動物數量的動態變化, 吳強等[12-13]研究了黃渤海甲殼類動物群落結構。上述這些研究的對象都是來自底拖網的個體較大的經濟甲殼類動物, 而對于采泥樣品中的甲殼類動物的系統研究仍然很缺乏。本文根據2014年秋季黃海大型底棲動物的調查資料, 研究了該海域底棲甲殼類的種類組成、豐度、生物量、群落結構以及與環境因子的相關性等, 通過與歷史資料的對比, 分析了底棲甲殼類物種類組成變化, 并首次利用BOPA 指數(BOPA)對黃海沉積環境質量狀況進行了評價, 以期為黃海底棲甲殼類生態學的深入研究提供參考。
1 材料與方法
1.1 研究海域
于2014年秋季(11月)搭乘“東方紅2號”海洋綜合調查船, 對黃海海域進行了大型底棲動物調查, 共設置29個站位, 站位見圖1。
1.2 樣品采集及處理
使用0.1 m2的箱式采泥器采樣, 每站成功采2次合為1個樣品; 現場利用渦旋器懸洗泥沙, 經孔徑為0.5 mm的網篩分選, 分選后留在網篩上的樣品殘渣全部收集裝瓶, 并用等體積10%福爾馬林溶液固定。每站取一定量的表層沉積物, ?20 ℃冷凍保存, 用于環境因子測定。野外及實驗室內樣品的采集和處理均按照《海洋調查規范: 第6部分海洋生物調查》[14]進行。
本研究所測環境因子包括水深、底層溫度、底層鹽度和沉積物粒度等, 其中水深、底層溫度、底層鹽度是由CTD現場測定, 粒度則使用Mastersizer3000型激光粒度儀進行測定。
1.3 數據處理分析
1.3.1 優勢種
采用相對重要性指數(Index of Relative Importance, 簡稱RI)來確定優勢種[15], 其計算公式如下:
RI(), (1)
式中,為每種生物量占總生物量的百分比,為每種的豐度占總豐度的百分比,為該種出現的頻率。
1.3.2 多樣性指數
采用香農維納多樣性指數(Shannon-Wiener diversity index,′)、均勻度指數(Pielou’s index,)、豐富度指數(Margalef’s index,)進行多樣性分析。計算公式如下:
=–∑(n/)*log2(n/), (2)
′=′/log2, (3)
=(–1)/log2, (4)
式中為樣品的總種數,為樣品的總個體數,n為第種的個體數。
1.3.3 BOPA指數
BOPA指數(benthic opportunistic polychaetes amphipods index, 簡稱BOPA)是指利用底棲多毛類機會種和端足類的比值來評價環境質量狀況[16]。計算公式:

f是多毛類機會種的個體總數與樣本個體總數的比值,f是端足類的個體總數與樣本個體總數的比值。根據BOPA的范圍, 將環境生態質量狀況分為5個等級:優(0.0~0.045 76), 未受污染; 良(0.045 76~ 0.139 66), 輕度污染; 中(0.139 66~0.193 82), 中度污染; 差(0.193 82~0.267 61), 重度污染; 劣(0.267 61~ 0.301 30), 嚴重污染。
1.3.4 數據分析
使用PRIMER6.0軟件, 進行群落結構分析, 對各站位的底棲甲殼類豐度進行平方根轉化使之標準化, 構建Bray-Curtis相似性矩陣進行聚類分析; 利用SPSS22.0軟件進行相關性等分析。
2 結果
2.1 沉積環境
本研究環境因子數據見表1。
由表1可以看出, 本研究的平均水深為53.64 m, 最深處高達82.2 m, 在H10站位; 最淺處是B26和B36站位, 為22 m。平均底溫12.77 ℃, 溫度范圍在8.93~20.23 ℃, 近岸的溫度高于中央冷水團, 近岸區溫度變化較大。平均底層鹽度31.82, 鹽度最高的是H26站位, 為33.165; 最低為30.954, 在H03站位。沉積物粒徑呈現出黃海西部近岸比黃海中部沉積物粒徑粗的特點。

表1 本研究各站位環境因子數據
注: “?”表示未測得該數據
2.2 底棲甲殼類種類組成與優勢種
本研究共采到底棲甲殼類71種, 隸屬于32科42屬。出現種數較多的科分別為蜾蠃蜚科Coro-phii-dae (9種)、馬耳他鉤蝦科Melitidae (6種)和雙眼鉤蝦科Ampeliscidae (6種)。出現種數較多的屬分別為擬鉤蝦屬(5種)、雙眼鉤蝦屬(5種)和馬耳他鉤蝦屬(4種)。物種數較多的站位集中在山東半島近岸附近, 黃海中部區域相對較少(圖2)。
表2列出了本研究中RI指數大小列前五位的種, 分別為太平洋方甲漣蟲(Hart, 1930)、口蝦蛄(De Haan, 1844)、灘擬猛鉤蝦(Hirayama, 1987)、日本沙鉤蝦(Dahl, 1945)和美原雙眼鉤蝦(Nagata, 1959)。其中, 太平洋方甲漣蟲的RI值最高, 其分布于23個站位, 出現率(分布站位數/總站位數)為79.31%; 集中分布在冷水團范圍, 包括山東半島的外圍海域和南黃海中部(圖3), 其中, 在B14站位達到豐度的最高值560 inds./m2。

表2 相對重要性指數排名前5位的底棲甲殼類物種
2.3 底棲甲殼類的豐度和生物量
底棲甲殼類的平均豐度為278 inds./m2, 豐度最高的站位在B14, 為850 inds./m2, 原因是該站位出現了大量的太平洋方甲漣蟲, 其豐度占大型底棲動物總豐度的65.88%; 豐度最低的站位在B26, 為60 inds./m2。本研究底棲甲殼類豐度高值區大體都位于北黃海遠岸海域(圖4a)。
底棲甲殼類的平均生物量為2.33 g/m2, 生物量最高的站位在H09, 為22.98 g/m2, 其次為H40站位, 原因是這兩個站位采集到了口蝦蛄; 生物量最低的站位在H18, 為0.014 g/m2。本研究底棲甲殼類生物量高值區大體都位于黃海中部(圖4b)。
2.4 底棲甲殼類多樣性指數
由表3看出, 各個站位底棲甲殼類種數()平均值為9.2, 種數最多的站位位于山東半島近岸的B05, 為21種, 最少的站位位于黃海中部的H14, 為3種。香農維納多樣性指數()平均值為2.22, 最高出現在山東半島近岸的B05站位, 達3.66, 最低出現在黃海中部的H14站位, 為0.48。
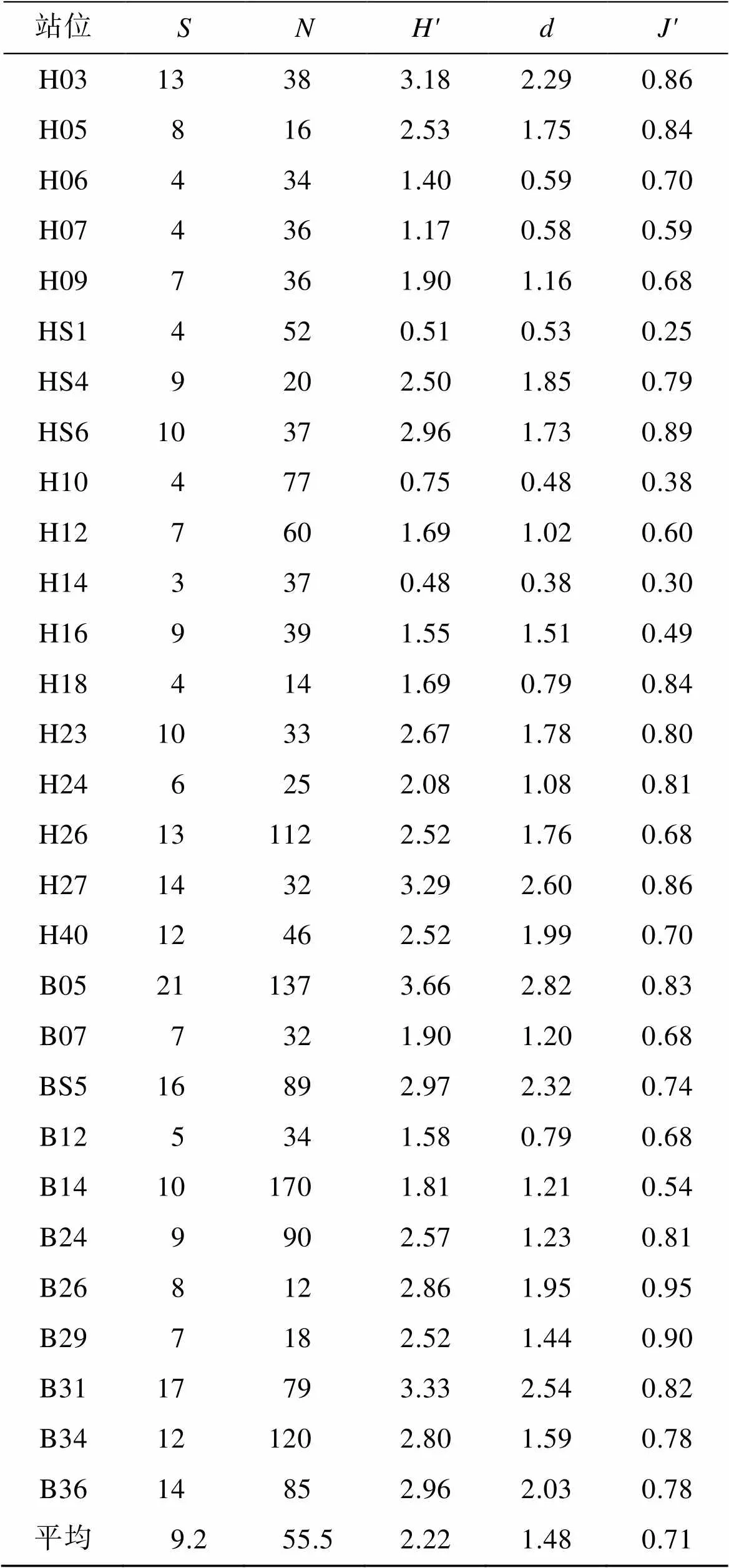
表3 研究海域底棲甲殼類多樣性指數
2.5 底棲甲殼類群落
在23%的相似度水平上可以劃分為6個群落(圖5)。群落Ⅰ和Ⅱ均由1個站位組成, 群落Ⅰ僅包含黃海西南部的H16站位, 其中豐度較高的物種為日本沙鉤蝦, 為145 inds./m2。群落Ⅱ僅包含H40站位, 位于黃海南部, 其中豐度較高的物種為鐮形葉鉤蝦(Montagu, 1808), 為120 inds./m2。群落Ⅲ: 塞切爾泥鉤蝦(Chevreux, 1901)-美原雙眼鉤蝦群落, 由南黃海西部的H03和H18組成, SIMPER分析表明該群落的平均相似性為38.46%, 其中塞切爾泥鉤蝦的貢獻率達到了70%。群落Ⅳ: 太平洋方甲漣蟲-灘擬猛鉤蝦-日本沙鉤蝦群落, 由18個站位組成, 這些站位與黃海冷水團范圍相當, 群落的平均相似性為30.78%, 其中太平洋方甲漣蟲的貢獻率達到了81.10%。群落Ⅴ: 日本長尾蟲(Shiino, 1937)-日本大螯蜚(Step-he-n-sen, 1938)-內海擬鉤蝦(Nagata, 1961)群落, 由北黃海西部的B29、B31和B34站位組成, 該群落的平均相似性為16.62%。群落Ⅵ: 灘擬猛鉤蝦-日本擬背尾水虱(Richardson, 1909)-潮間海鉤蝦(Ren, 1992)群落, 由山東半島近岸4個站位組成, 該群落的平均相似性為15.63%。ANOSIM分析表明各群落間物種組成差異顯著(Global=0.789,=0.001)。
2.6 底棲甲殼類豐度與環境因子的關系
為探討環境因子對底棲甲殼類豐度的影響, 本研究將底棲甲殼類總豐度和優勢種太平洋方甲漣蟲的豐度與水深、底溫、底鹽、平均粒徑和中值粒徑進行了Pearson相關性分析。結果(表4)顯示, 底棲甲殼類總豐度與各環境因子的相關性均不顯著, 而太平洋方甲漣蟲豐度與水深呈極顯著正相關, 與底鹽呈顯著正相關, 與底溫呈顯著負相關。
2.7 BOPA指數
圖6為本研究各站位的BOPA。除個別站位外, 大部分站位的BOPA都為0~0.13 966, 指示沉積環境狀況優良。位于黃海南部的H27站位的BOPA為0.139 66~ 0.193 82, 指示沉積環境受到中度擾動, 位于黃海冷水團中部的H05、HS4、H14站位的BOPA為0.193 82~ 0.267 61, 指示沉積環境受到重度擾動。根據BOPA, 除部分站位外, 黃海海域沉積環境整體良好。

表4 研究海域底棲甲殼類豐度與環境因子的相關性
注: *表示相關性顯著,0.05(雙尾); **表示相關性極顯著,<0.01(雙尾)。
3 討論
3.1 本研究結果與歷史資料的對比
本研究共采到底棲甲殼類71種, 物種數較多的站位分布在山東半島近岸附近, 一方面可能由于近岸有機質含量較高, 吸引了更多的底棲甲殼類; 另一方面是由于秋季近岸水域溫度比冬春季高, 可能導致較多的種類分布在近岸; 而黃海中部由于冷水團的存在, 限制了一些屬暖溫帶和亞熱帶混合類型的底棲甲殼類的分布[17], 導致了黃海中部底棲甲殼類的物種數較低。
本研究結果與歷史資料的對比見表5。從表5可以看出, 本研究底棲甲殼類的種數和豐度, 比之前年份的研究資料都略高。彭松耀等[18]報道過, 一些游泳性的底棲甲殼類會由于索餌和越冬導致其豐度出現季節變動, 在本文所列出的歷史資料中(表5), 采樣季節并不相同, 可能導致了不同研究之間底棲甲殼類物種數的差異。設置的站位的多少與采樣區域的不同等, 也會對底棲甲殼類的豐度和物種數產生影響[19]。具體原因還有待進一步研究。

表5 本研究結果與歷史資料的對比
注: “-”表示文獻中未有該數據
此外, 在歷年的研究結果中, 底棲甲殼類種數在大型底棲動物中的占比都在30%左右, 說明底棲甲殼類在黃海大型底棲動物類群組成中變化不大。
3.2 太平洋方甲漣蟲的豐度與環境因子的關系探討
太平洋方甲漣蟲隸屬于軟甲綱Malacostraca、漣蟲目Cumacea、尖額漣蟲科Leuconidae、方甲漣蟲屬, 在渤海、黃海、日本沿海、顎霍次克海、白令海和北美洲太平洋沿岸都有分布[24]。張均龍[22]和ZHANG等[25]在1992年南黃海定量采泥中, 首次報道小個體的太平洋方甲漣蟲為南黃海冷水團的優勢種之一, 此后, 劉衛霞等[23]和楊傳平[20]也均報道過此現象。
太平洋方甲漣蟲在本研究中具有極高的優勢, 在近80%的站位樣品中出現, 集中分布在黃海冷水團范圍。黃海冷水團在冬季時形成, 經過春季的發展, 至7—8月份達到鼎盛期。本研究取樣時間處于冷水團逐漸消退階段, 太平洋方甲漣蟲為冷水性種[24], 能夠適應低溫高鹽環境, 因此可以在此大量繁殖, 形成較高的優勢。
就像20世紀60年代出現的引人注意的魚類資源小型化一樣[26], 大型底棲動物也同樣存在此現象。如徐勇報道了從20世紀50年代到21世紀, 生命周期較短的小型多毛類動物, 取代了生命周期較長的大型軟體動物、棘皮動物和多毛類動物等, 成為南黃海西部海域大型底棲動物的優勢種[27], 并且提出群落變化可能會向離岸較遠的南黃海東部區域擴展[28]。早在20世紀中葉, 劉瑞玉先生等報道的黃海冷水團優勢種[28]并不包括太平洋方甲漣蟲, 而現在該種卻成為研究海域的最占優勢的物種, 這極有可能是底棲甲殼類動物應對環境變遷, 出現的優勢種更替現象。太平洋方甲漣蟲個體很小, 平均體長僅4.38 mm[24], 這可能是黃海底棲甲殼類響應環境變化而小型化的一種表現, 值得進一步深入研究。
3.3 利用IBOPA來評價海域沉積環境質量狀況
2000年Gómez-Gesteira和Dauvin首次提出了利用多毛類機會種和端足類指數, 來確定溢油對軟底質底棲動物群落的影響。多毛類和端足類底棲生物是環境擾動的指示生物, 當環境擾動增加時, 生命周期短、成熟快、繁殖效率高的多毛類機會種占據優勢地位, 個體數量大量增加, 是環境污染或人為擾動后的先鋒底棲生物[29]; 而端足類動物在缺氧的沉積物中建管, 促進海底的氧化過程和硝化過程, 可作為環境修復的指示生物[29-30]。因此, 利用多毛類機會種和端足類的比值(BOPA)來研究沉積環境的擾動狀況具有一定的意義。目前, 國內對于BOPA的研究較少, 僅見張瑩等[29, 31-32]、任中華[33]和李少文等[34]利用BOPA對萊州灣和黃河口的環境質量狀況進行的評價, 余驥[35]和劉志權[36]利用BOPA對崇明東灘潮間帶的研究, Xu等[37]利用BOPA對蘇北淺灘生態狀況進行評價。在這些研究中,BOPA均能比較準確地評價研究區域的環境質量狀況。
在本研究中, 根據BOPA, 除黃海中部和南部部分站位受到中度和重度擾動外, 大部分站位處于優和良范圍, 即黃海沉積環境整體良好。李少月[38]利用ABC曲線方法對黃海沉積物環境質量狀況進行的評價, 也顯示黃海多數站位未受擾動, 僅南黃海中部的部分站位受到擾動, 與本研究結果基本一致。王教凱[39]等在2011年對南黃海沉積物中有機氯農藥(OCPs)的研究發現, OCPs濃度高值出現在南黃海中部泥質區, 其范圍與本文黃海中部相當。山東半島附近海域是現代黃河泥沙的主要沉積區, 沿岸流帶來大量泥砂的同時, 也攜帶了大量黃河流域的污染物進入南黃海, 這可能是造成擾動的原因之一[40]。
綜上所述, 本文初步認為調查海域沉積環境良好, 僅黃海中部和南部部分站位受到了中度擾動,BOPA能較好地評價研究區域的環境質量狀況。
4 結論
1) 底棲甲殼類在歷年的黃海大型底棲動物類群組成中變化不大, 占比都在30%左右。
2) 太平洋方甲漣蟲具有極高的優勢, 底溫、底鹽和水深是影響其豐度的主要環境因子。
3) 研究海域沉積環境整體良好, 僅黃海中部和南部部分站位受到了中度擾動。
4)BOPA能較好地評價海域的沉積環境質量狀況。
[1] BOIX D, GASCóN S, SALA J, et al. Patterns of composition and species richness of crustaceans and aquatic insects along environmental gradients in Mediterranean water bodies[J]. Hydrobiologia, 2008, 597(1): 53-69.
[2] DANA J D. Conspectus of the Crustacea of the exploring expedition under Capt. Wilkes, U. S. N.[J]. Proceedings of the Academy of Natural Sciences of Philadelphia, 1852, 6: 73-86.
[3] 何明海. 利用底棲生物監測與評價海洋環境質量[J]. 海洋環境科學, 1989, 8(4): 49-54.
HE Minghai. Monitoring and evaluation of marine environmental quality by benthos[J]. Marine Environmental Science, 1989, 8(4): 49-54.
[4] Abelló P, Valladares F J, Castellón A. Analysis of the structure of decapod crustacean assemblages off the Catalan coast (North-West Mediterranean)[J]. Marine Biology, 1988, 98(1): 39-49.
[5] 黃梓榮, 陳作志, 鐘智輝, 等. 南海北部陸架區甲殼類的種類組成和資源密度分布[J]. 上海海洋大學學報, 2009, 18(1): 59-65.
HUANG Zirong, CHEN Zuozhi, ZHONG Zhihui, et al. Species composition and resource density of crustaceans in the continental shelf of northern South China Sea[J]. Journal of Shanghai Ocean University, 2009, 18(1): 59-65.
[6] 盧占暉, 徐漢祥, 薛利建, 等. 東海中部蝦類群落的季節變化研究[J]. 浙江海洋學院學報(自然科學版), 2015, 34(6): 497-504.
LU Zhanhui, XU Hanxiang, XUE Lijian, et al. A study on seasonal changes of the shrimp communities in the middle East China Sea[J]. Journal of Zhejiang Ocean University (Natural Science), 2015, 34(6): 497-504.
[7] 宋海棠. 東海蝦類的生態群落與區系特征[J]. 海洋科學集刊, 2002, 44: 124-133.
SONG Haitang. The ecological colony and fauna characteristics of East China Sea shrimp[J]. Studia Marina Sinica, 2002, 44: 124-133.
[8] 俞存根, 陳小慶, 宋海棠, 等. 春季東海海域蝦類群落結構及其多樣性[J]. 生態學報, 2009, 29(7): 3593- 3604.
YU Cungen, CHEN Xiaoqing, SONG Haitang, et al. Analysis on community structure and diversity of shri-mps in the East China Sea in spring[J]. Acta Ecologica Sinica, 2009, 29(7): 3593-3604.
[9] 陳小慶, 俞存根, 虞聰達, 等. 東海中南部外海蝦類群落結構特征分析[J]. 水生生物學報, 2009, 33(4): 664-673.
CHEN Xiaoqing, YU Cungen, YU Congda, et al. Com-munity characteristics of shrimps in the offshore waters of the mid-southern East China Sea[J]. Acta Hydrobio-logica Sinica, 2009, 33(4): 664-673.
[10] 李惠玉. 東海區甲殼動物群落結構的研究[C]//中國水產學會. 2007年中國水產學會學術年會暨水產微生態調控技術論壇論文摘要匯編. 北京: 中國水產學會, 2007.
LI Huiyu. Study on crustacean community structure in East China Sea[C]// Chinese Fisheries Society. Abstract compilation of papers of the 2007 academic annual meeting of the Chinese Fisheries Society and the forum on aquatic microecological regulation technology. Beijing: Chinese Fisheries Society, 2007.
[11] 于海燕, 李新正, 李寶泉, 等. 膠州灣大型底棲甲殼動物數量動態變化[J]. 海洋與湖沼, 2005, 36(4): 289-295.
YU Haiyan, LI Xinzheng, LI Baoquan, et al. Distribution of macrobenthic crustacean in Jiaozhou Bay[J]. Ocea-no-logia et Limnologia Sinica, 2005, 36(4): 289-295.
[12] 吳強, 王俊, 李忠義, 等. 黃渤海春季甲殼類群落結構的空間變化[J]. 水產學報, 2012, 36(11): 1685-1693.
WU Qiang, WANG Jun, LI Zhongyi, et al. Spatial varia-tion of crustacean community structure in Yellow Sea and Bohai Sea in spring[J]. Journal of Fisheries of China, 2012, 36(11): 1685-1693.
[13] 吳強, 李忠義, 戴芳群, 等. 黃渤海甲殼類的分類多樣性[J]. 生物多樣性, 2016, 24(11): 1306-1314.
WU Qiang, LI Zhongyi, DAI Fangqun, et al. Taxono-mic diversity of crustaceans in the Yellow Sea and Bohai Sea[J]. Biodiversity Science, 2016, 24(11): 1306-1314.
[14] 國家質量監督局和國家標準化委員會. GB/T 12763. 6-2007海洋調查規范第6部分: 海洋生物調查[S]. 北京: 中國標準出版社, 2007.
The State Bureau of Quality and Technical Supervision and Standardization Administration of the People's Republic of China. Specifications for oceanographic survey-part 6: marine biological survey[S]. Beijing: Standards Press of China Press, 2007.
[15] PIANKA E R. Ecology of the Agamid Lizardin Western Australia[J]. Copeia, 1971, 1971(3): 527-536.
[16] DAUVIN J C, RUELLET T. Polychaete/amphipod ratio revisited[J]. Marine Pollution Bulletin, 2007, 55(1/6): 215-224.
[17] 徐勤增, 李瑞香, 王宗靈, 等. 南黃海夏季大型底棲動物分布現狀[J]. 海洋科學進展, 2009, 27(3): 393-399.
XU Qinzeng, LI Ruixiang, WANG Zongling, et al. Macrobenthos distribution of the South Yellow Sea in summer[J]. Advances in Marine Science, 2009, 27(3): 393- 399.
[18] 彭松耀, 李新正, 徐勇, 等. 黃海大型底棲動物功能攝食類群的空間格局[J]. 海洋與湖沼, 2020, 51(3): 456-466.
PENG Songyao, LI Xinzheng, XU Yong, et al. Spatial distribution pattern of macrobenthos feeding functional groups in the Yellow Sea[J]. Oceanologia et Limnologia Sinica, 2020, 51(3): 456-466.
[19] 彭松耀. 黃東海大型底棲動物群落結構特征[D]. 青島: 中國科學院研究生院(中國科學院海洋研究所), 2013.
PENG Songyao. Characteristics of microbenthic community structure in the Yellow Sea and East China Sea[D]. Qingdao: University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Sciences), 2013.
[20] 楊傳平. 南黃海大型底棲動物生態學研究[D]. 青島: 中國海洋大學, 2014.
YANG Chuanping. The ecological study on Macrobenthos in the Southern Yellow Sea, China[D]. Qingdao: Ocean University of China, 2014.
[21] 賈海波, 胡顥琰, 唐靜亮, 等. 南黃海大型底棲生物生態調查與研究[J]. 海洋與湖沼, 2010, 41(6): 842-849.
JIA Haibo, HU Haoyan, TANG Jingliang, et al. The macrobenthos ecology of the Southern Yellow Sea[J]. Ocea-nologia et Limnologia Sinica, 2010, 41(6): 842-849.
[22] 張均龍. 黃海大型底棲生物多樣性與群落結構的研究[D]. 青島: 中國科學院研究生院(中國科學院海洋研究所), 2012.
ZHANG Junlong. Biodiversity and community structure of macrobenthos in the Yellow Sea[D]. Qingdao: University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Sciences), 2012.
[23] 劉衛霞, 于子山, 曲方圓, 等. 北黃海冬季大型底棲動物種類組成和數量分布[J]. 中國海洋大學學報(自然科學版), 2009, 39(S1): 115-119.
LIU Weixia, YU Zishan, QU Fangyuan, et al. Species composition and quantitative distribution of abundance and biomass of macrobenthos in the North Yellow Sea in winter[J]. Periodical of Ocean University of China, 2009, 39(S1): 115-119.
[24] 沙忠利, 任先秋, 王永良. 膠州灣及青島鄰近海域底棲甲殼動物(上冊)[M]. 北京: 科學出版社, 2017: 280-282.
SHA Zhongli, REN Xianqiu, WANG Yongliang. Marine benthic Crustracea from Jiaozhou Bay and Qingdao adjacent waters(1)[M]. Beijing: Science Press, 2017: 280-282.
[25] ZHANG J L, XU F S, LIU R Y. Community structure changes of macrobenthos in the South Yellow Sea[J]. Chinese Journal of Oceanology and Limnology, 2012, 30(2): 248-255.
[26] 劉其根, 沈建忠, 陳馬康, 等. 天然經濟魚類小型化問題的研究進展[J]. 上海水產大學學報, 2005, 14(1): 79-83.
LIU Qigen, SHEN Jianzhong, CHEN Makang, et al. Advances of the study on the miniaturization of natural economical fish resources[J]. Journal of Shanghai Fisheries University, 2005, 14(1): 79-83.
[27] 徐勇. 黃東海大型底棲動物多樣性變化及黑潮的影響[D]. 青島: 中國科學院大學(中國科學院海洋研究所), 2017.
XU Yong. Variations of macrofaunal diversity in the Yellow Sea and East China Sea and the influence of Kuroshio Current[D]. Qingdao: University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Sciences), 2017.
[28] 劉瑞玉, 崔玉珩, 徐鳳山, 等. 黃海、東海底棲生物的生態特點[J]. 海洋科學集刊, 1986, 27: 154-173.
LIU Ruiyu, CUI Yuheng, XU Fengshan, et al. Ecological characteristics of macrobenthos of the Yellow Sea and the East China Sea[J]. Studia Marina Sinica, 1986, 27: 154-173.
[29] 張瑩, 李少文, 呂振波, 等. 多毛類底棲動物在萊州灣生態環境評價中的應用[J]. 生態學報, 2013, 33(8): 2522-2530.
ZHANG Ying, LI Shaowen, LV Zhenbo, et al. Appli-cation of polychaete in ecological environment evaluation of Laizhou Bay[J]. Acta Ecologica Sinica, 2013, 33(8): 2522-2530.
[30] GESTEIRA J L G, DAUVIN J C. Amphipods are good bioindicators of the impact of oil spills on soft-bottom macrobenthic communities[J]. Marine Pollution Bulletin, 2000, 40(11): 1017-1027.
[31] 張瑩, 劉元進, 張英, 等. 萊州灣多毛類底棲動物生態特征及其對環境變化的響應[J]. 生態學雜志, 2012, 31(4): 888-895.
ZHANG Ying, LIU Yuanjin, ZHANG Ying, et al. Ecological characteristics of hairy benthos and their response to environmental changes in Laizhou Bay[J]. Chinese Journal of Ecology, 2012, 31(4): 888-895.
[32] 張瑩, 呂振波, 徐宗法, 等. 環境污染對小清河口大型底棲動物多樣性的影響[J]. 生態學雜志, 2012, 31(2): 381-387.
ZHANG Ying, LV Zhenbo, XU Zongfa, et al. Effects of environmental pollution on the diversity of macrobenthos in Xiaoqing Estuary[J]. Chinese Journal of Ecolo-gy, 2012, 31(2): 381-387.
[33] 任中華. 萊州灣及黃河口海域大型底棲動物群落結構多樣性及其生態學研究[D]. 上海: 上海海洋大學, 2016.
REN Zhonghua. The diversity of community and the ecology of macrobenthos in Laizhou Bay[D]. Shanghai: Shanghai Ocean University, 2016.
[34] 李少文, 張瑩, 李凡, 等. 調水調沙對黃河口海域大型底棲動物群落的影響[J]. 環境科學研究, 2015, 28(2): 259-266.
LI Shaowen, ZHANG Ying, LI Fan, et al. Effects of water and sediment discharge regulation on macrobenthic community in the Yellow River Estuary[J]. Research of Environmental Sciences, 2015, 28(2): 259-266.
[35] 余驥. 崇明東灘潮間帶大型底棲動物群落的生態學研究[D]. 上海: 華東師范大學, 2014.
YU Ji. The ecological study on macrozoobenthic com-mu-nities in the intertidal zone of Chongming east shoal[D]. Shanghai: East China Normal University, 2014.
[36] 劉志權. 崇明東灘大型底棲動物對人類活動的響應及生態修復研究[D]. 上海: 華東師范大學, 2017.
LIU Zhiquan. Response of macrobenthos on human activities and the ecological restoration in east shoal of Chongming island[D]. Shanghai: East China Normal University, 2017.
[37] XU Y, LI X Z, WANG H F, et al. Status of macrobenthic diversity and distribution in Subei Shoal, China[J]. Aquatic Ecosystem Health & Management, 2016, 19(4): 411-419.
[38] 李少月. 黃海春秋季大型底棲動物生態學研究及功能群初探[D]. 青島: 中國海洋大學, 2016.
LI Shaoyue. Preliminary study on the ecology and functional groups of macrobenthos in the spring and autumn of the Yellow Sea[D]. Qingdao: Ocean university of China, 2016.
[39] 王教凱. 生物標志物指示南黃海沉積物中有機氯農藥的分布及來源的可行性研究[D]. 青島: 中國海洋大學, 2014.
WANG Jiaokai. Feasibility study on biomarkers indicating the distribution and source of organochlorine pesticides (OCPs) in sediment from Southern Yellow Sea (SYS)[D]. Qingdao: Ocean University of China, 2014.
[40] 李孟芳, 張大海, 段曉勇, 等. 南黃海表層沉積物中鄰苯二甲酸酯的分布特征[J]. 海洋環境科學, 2014, 33(5): 682-687.
LI Mengfang, ZHANG Dahai, DUAN Xiaoyong, et al. Distribution of phthalic acid esters (PAEs) in surface sediments of the Southern Yellow Sea[J]. Marine Environmental Science, 2014, 33(5): 682-687.
Primary study on the diversity and community structure of macrobenthic crustaceans in the Yellow Sea during autumn
ZHU Lin1, LI Shao-yue2, WANG Zhi3, ZHANG Meng-sheng4, CHEN Chen1, TENG An-kang1, SHAO Cong-cong1, YU Zi-shan1
(1. College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China; 2. North Sea Environmental Monitoring Center, State Oceanic Administration, Qingdao 266003, China; 3. State Key Laboratory of Marine Environmental Science, College of Ocean and Earth Sciences, Xiamen University, Xiamen 361102, China; 4. Survey and Design Institute of Shanghai, Shanghai 200050, China)
The species composition, abundance, biomass, diversity, and community structure of the macrobenthic crustaceans were studied based on Yellow Sea macrobenthos data from November 2014. A total of 71species of macrobenthic crustaceans were identified from 32 families and 45 genera, with an average abundance of 278 inds./m2and biomass of 2.33 g/m2.Hart, 1930 had the highest Index of Relative Importance value. The cluster analysis showed that the macrobenthic crustaceans in the surveyed area were divided into six communities at a similarity level of 23%. The result of Pearson’s correlation analysis showed that the abundance ofHart, 1930 was strongly positively correlated with water depth and bottom salt and negatively correlated with bottom temperature. In previous studies, the proportion of macrobenthos among benthic crustacean species did not change much, accounting for about 30% of the total number of macrobenthic species. The benthic ecological environment in the surveyed area was undisturbed according to the benthic opportunistic Polychaeta Amphipoda (BOPA) index results, and only the central and southern parts of the Yellow Sea were moderately disturbed. The environmental quality of the surveyed area was well evaluated by the BOPA index.
Yellow Sea; macrobenthic crustaceans; community; BOPA index; environmental factors
Mar. 4, 2022
Q958.8
A
1000-3096(2022)09-0098-11
10.11759/hykx20220304001
2022-03-04;
2022-04-06
科技部項目(2012FY112200); 國家自然科學基金項目(41976100)
[Ministry of Science and Technology Projects, No. 2012FY112200; National Natural Science Foundation of China, No. 41976100]
祝琳(1997—), 女, 山東青島人, 碩士生, 研究方向為海洋底棲生物生態學, E-mail: 805531826@qq.com; 于子山(1963—),通信作者, 男, 山東青島人, 副教授, 研究方向為海洋底棲生物生態學, E-mail: yu_zishan@ouc.edu.cn
(本文編輯: 趙衛紅)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
