Sap30調節小鼠造血干細胞的損傷恢復和體外擴增*
2022-11-01 03:25:14黃璐圓張錦華鞠振宇陳陟陽
中國病理生理雜志 2022年10期
關鍵詞:小鼠
詹 薔, 黃璐圓, 張錦華, 劉 進, 鞠振宇, 陳陟陽△
(1暨南大學教育部再生醫學重點實驗室,衰老與再生醫學研究院,廣東 廣州 510632;2廣州實驗室,廣東 廣州 510005;3暨南大學附屬第一醫院心內科,廣東 廣州 510630)
在生理條件下,造血干細胞(hematopoietic stem cells,HSCs)在自我更新和多能分化之間保持平衡[1],確保血液系統中成熟細胞的數量穩定且保證HSC池不會耗盡。作為造血系統中最上游的細胞,HSCs在骨髓中占比很小,但在許多造血相關疾病的治療中發揮著不可替代的作用[2]。HSCs移植是臨床治療血液學及免疫相關疾病最有效的方法,然而HSCs來源的稀缺性極大限制了其臨床應用,如何有效解決HSCs體外擴增是領域內亟待解決的問題[3]。化療是廣泛應用于多種腫瘤治療的重要手段,然而化療極大地影響了患者HSCs的功能,進而抑制了化療患者的自身免疫力。若能有效促進化療后HSCs數目的恢復,將提高化療患者的免疫力并促進血液系統的重建。雖然有許多研究從多種角度探究如何實現HSCs的有效功能性擴增[4-7],但其具體機制仍不明確。
組蛋白乙酰化作為蛋白質翻譯后修飾的重要調控機制,在調控HSCs的功能中起到重要作用[8]。組蛋白乙酰化修飾主要通過乙酰轉移酶和脫乙酰酶的協同作用,調節乙酰基在賴氨酸上的轉移進而調節染色質結構和動態,從而上調基因轉錄[9];相反,組蛋白脫乙酰酶通過去除乙酰賴氨酸殘基上的乙酰化標記而抑制基因表達。Sap30作為Sin3A和NCoR共抑制因子復合物的配體,可以招募組蛋白脫乙酰酶實現組蛋白去乙酰化。有報道顯示,Sap30基因的缺失通過調控染色質可及性的改變抑制了成體細胞基因的表達,進而促進了成纖維細胞重編程為多潛能干細胞[10]。然而,Sap30介導的乙酰化修飾在HSCs損傷恢復及體外擴增中的作用尚未見報道。本研究應用Sap30基因缺失(Sap30-/-)小鼠模型,開展針對Sap30在小鼠HSCs中功能的研究,以動物模型開展的體內、外實驗數據為基礎,為以后HSCs的應用提供參考資料。
材料和方法
1 動物
Sap30-/-及野生型(Sap30+/+)對照小鼠(均為白細胞表面抗原CD45.2陽性小鼠)由中國科學院廣州生物與健康研究院提供,經由廣東省實驗動物檢測所凈化,生產許可證號為SCXK(粵)2018-0044,凈化后引進并飼養繁殖于暨南大學實驗動物中心屏障環境動物房[SYXK(粵)2017-0174],雌雄及數量不限,2~20月齡。HSCs競爭性移植的供體:CD45.2+小鼠;競爭者:CD45.1+小鼠;受體:CD45.1+CD45.2+雙陽性小鼠飼養于暨南大學實驗動物中心屏障環境動物房[SYXK(粵)2017-0174],各亞型之間沒有功能性的區別。本實驗已獲得暨南大學動物實驗中心倫理委員會批準,并嚴格按照動物使用原則執行。
2 主要儀器和試劑
LSRFortessa流式細胞分析儀和FACSAria III流式細胞分選儀(BD);RS 2000 Pro生物輻射系統(Rad Source);XN-1000V(B1)全自動五分類動物血液分析儀(Sysmex);5% Baytril購 自Bayer;Biotin-CD4(100508) 、Biotin-CD8(100704) 、Biotin-B220(103204)、Biotin-CD11b(101204)、Biotin-Ter-119(116204) 、PerCP-Cy5.5-IL-7R(135022) 、PECD45.2(109808)、PE-CD150(115927)、PE-Cy7-Sca-1(122514)、APC-Cy7-SA(405208)、FITC-CD4(100510)、FITC-CD8(100706)、APC-B220(103212)和FITC-CD48(103443)購自Invitrogen;PE-Flt3(12-1351-83)、APC-c-Kit(17-1171-83)、AF700-CD16/32(56-0161-82)和PerCP-Cy5.5-CD45.1(45-0453-82)購自Thermo Fisher;紅細胞裂解液(555899)和FITCCD34(553733)購自BD Pharmingen;penicillin/streptomycin購自Invitrogen;DAPI(D9542-10MG)購自Sigma;TPO(78072.1)、SCF(78064.1)和SFEM(serumfree expansion medium;09655)購 自STEMCELL Technologies;70 μm濾 膜(22363548)購 自Fisherbrand;異氟烷(R510-22-2)購自RWD;Anti-APC Microbeads(130-090-855)和LS Separation Columns(130-042-401)購自Miltenyi。
3 方法
3.1 流式細胞術分析骨髓中造血干/祖細胞 以野生型小鼠作為對照組,Sap30基因敲除小鼠作為實驗組,每組5~8 只小鼠。對野生型和基因敲除小鼠采取頸椎脫臼犧牲[11],取出股骨、脛骨、髂骨及脊柱后,放入研缽中用10 mL 緩沖液(1× PBS+2%胎牛血清FBS)研磨擠壓出骨髓細胞,經過200 目濾膜過濾后取10 μL 細胞懸液計數。將細胞懸液300×g離心5 min 后重懸至細胞濃度為1×1011/L,從中取出100 μL細胞懸液加入 10 μL Lin+抗體混合物[CD4、CD8、B220、Ter119、CD11b 和Gr1 與生物素(biotin)偶聯的抗體],在冰上避光孵育30 min;緩沖液洗去抗體后再次300×g離心5 min,棄上清后每個樣本加入30 μL 含其他表面標記抗體的混合物(表1)重懸[12],在冰上避光孵育30 min;緩沖液洗去抗體后再次300×g離心5 min,棄上清后每個樣本加入500 μL緩沖液重懸細胞并轉移至流式管中,使用LSRFortessa 流式細胞儀收集樣本,收集前加入DAPI(1 g/L),使用Flow-Jo 10對流式細胞儀記錄的數據進行處理。

表1 造血干/祖細胞流式分析染色方案Table 1. Antibody combination for flow cytometric staining of hematopoietic stem/progenitor cells
3.2 流式細胞術分析小鼠脾臟、胸腺和外周血中各類成熟細胞 以野生型小鼠作為對照組,Sap30基因敲除小鼠作為實驗組,每組5~8 只小鼠。對野生型和基因敲除小鼠進行深度麻醉后取外周血并采取頸椎脫臼犧牲,取出脾臟和胸腺后,經過200 目濾膜過濾后用5 mL 緩沖液(1×PBS+2%胎牛血清FBS)研磨成細胞懸液并計數,將細胞懸液300×g離心5 min 后重懸至細胞濃度為1×1011/L,從中取出20 μL 細胞懸液加入表面標記抗體混合物(表2),在冰上避光孵育30 min;緩沖液洗去抗體后再次300×g離心5 min,棄上清后每個樣本加入200 μL緩沖液重懸細胞并轉移至流式管中,使用LSRFortessa 流式細胞儀收集樣本,收集前加入DAPI(1 g/L),使用FlowJo 10 對流式細胞儀記錄的數據進行處理。
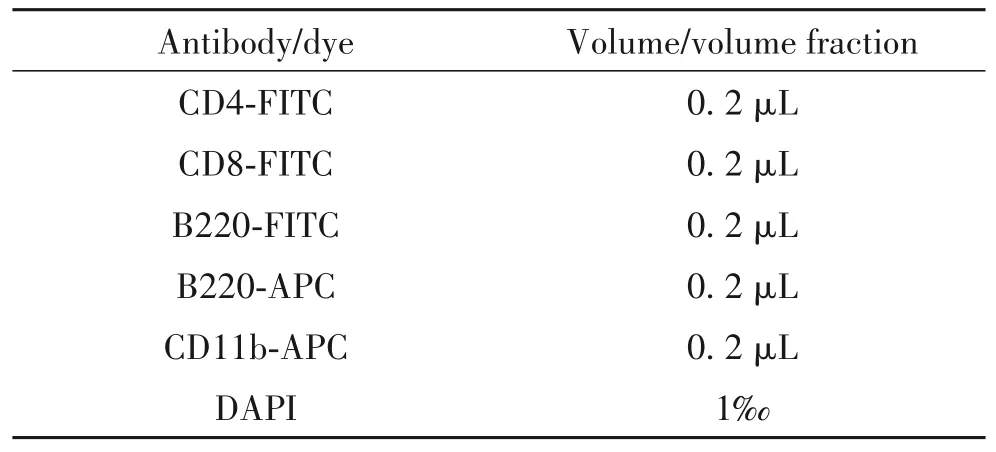
表2 B細胞、T細胞和髓系細胞的流式分析染色方案Table 2. Antibody combination for flow cytometric staining of B/myeloid/T cells
3.3 小鼠造血干/祖細胞流式分選 以野生型小鼠作為對照組,Sap30基因敲除小鼠作為實驗組,每組3~5 只小鼠。參考3.1 方法制備骨髓細胞懸液后,按照干細胞富集試劑盒步驟進行c-kit+富集后再進行表面抗體標記染色(參考3.1 抗體染色步驟),使用FACSAria III 流式分選儀分選造血干/祖細胞于染色緩沖液中。
3.4 HSCs 競爭性移植模型建立及移植小鼠嵌合率分析 以野生型小鼠作為供體對照組,Sap30基因敲除小鼠作為供體實驗組,每組5~8 只小鼠(表面抗原CD45.2+)。參考3.3 造血干/祖細胞流式分選步驟,分選出野生型及Sap30基因缺失的供體小鼠LSK 細胞;同時制備競爭者小鼠(表面抗原CD45.1+)的骨髓單細胞懸液,將 1 000 個供體 LSK 與 1×106個競爭者小鼠骨髓細胞混合(200 μL/受體),一同經尾靜脈移植到經過致死劑量(8 Gy,X-ray)照射的受體小鼠(表面抗原CD45.1/2 雙陽性,輻照后殘留<5%)中,移植之后對受體小鼠進行一個月抗生素(含1%拜有利的飲用水)處理[13-14]。移植后每4 周對小鼠采取麻醉后眼眶后靜脈取外周血后進行表面抗體染色(表3),使用LSRFortessa流式細胞儀收集樣本統計嵌合率。
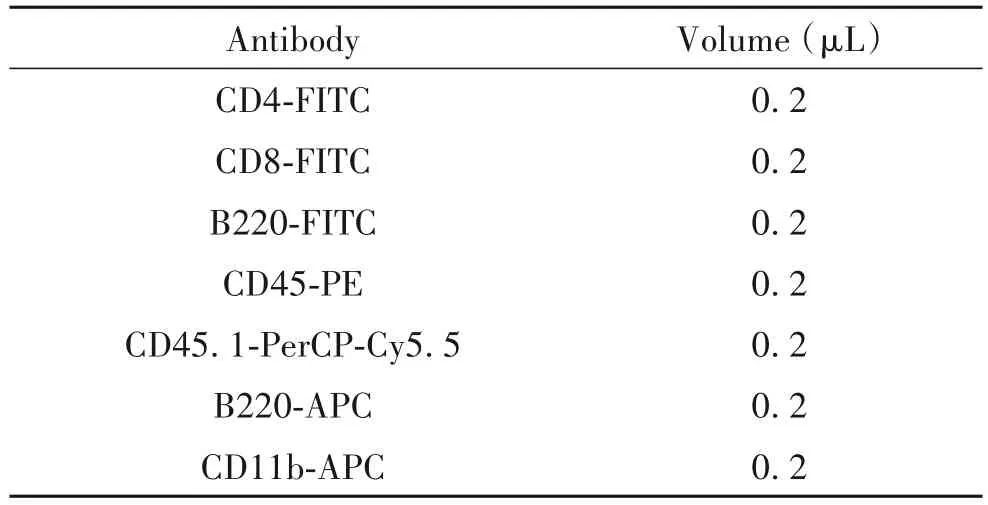
表3 造血干細胞移植受體小鼠外周血流式分析染色方案Table 3. Antibody combination for flow cytometric staining of peripheral blood cells
3.5 5-氟尿嘧啶(5-fluorouracil,5-FU)處理模型建立及分析 以野生型小鼠作為對照組,Sap30基因敲除小鼠作為實驗組,每組4~5 只小鼠。對野生型和Sap30基因敲除小鼠分別注射 5-FU(150 mg/kg)[15],并在6 d 和12 d 后犧牲小鼠并取出骨髓使用流式細胞儀進行分析,統計各組小鼠造血干/組細胞在全骨髓細胞中所占比例及絕對數。
3.6 HSCs 體外培養及分析 參考3.3 分選出野生型和基因敲除小鼠的造血干/祖細胞進行體外培養[16],每孔種入 10 000 個細胞,使用 48 孔細胞培養板培養,培養條件為SFEM 培養液+50 μg/L TPO+50 μg/L SCF+1% penicillin/streptomycin,5% CO2,37 ℃培養箱培養6、7 d 后,收集細胞進行表面抗體染色,并使用流式細胞儀分析。
4 統計學處理
實驗數據均采用均值±標準差表示(mean±SD),并利用GraphPad Prism 軟件進行后續的統計分析。數據在滿足正態分布并通過方差齊性檢驗的條件下,兩組間比較采用Student'st檢驗進行顯著性分析,當P<0.05時認為差異有統計學意義。
結 果
1 Sap30在髓系細胞中高表達
應用Sap30基因報告小鼠(圖1A),使用流式細胞術檢測各類血細胞中mCherry+細胞比例(圖1B)。骨髓分析結果顯示,Sap30基因在造血干/祖細胞中高表達,隨著下游向淋系發育表達逐漸降低(圖1C)。mCherry+細胞在外周血、骨髓、脾臟和胸腺中的髓系細胞(CD11b+細胞)中的占比顯著高于淋系細胞(B、T細胞;P<0.05;圖1D~G)。
2 Sap30基因缺失不影響穩態造血
與野生型小鼠相比,Sap30-/-小鼠骨髓、外周血、胸腺和脾臟中B 細胞、T細胞和髓系細胞的比例無顯著差異(P>0.05;圖2A~D)。流式細胞術結果顯示,穩態下與野生型小鼠相比,Sap30-/-小鼠骨髓中HSCs(CD34+CD150+LSK)、共同淋系祖細胞(common lymphoid progenitors,CLPs)和共同髓系祖細胞(common myeloid progenitors,CMPs)的百分比無顯著差異(P>0.05;圖2E~G)。HSCs 單克隆集落形成實驗結果顯示,與野生型相比,Sap30基因缺失的HSCs集落的大小沒有顯著差異(P>0.05;圖2H)。移植后分別在第1、2 和3 個月檢測受體小鼠外周血中供體LSK 來源的血細胞比例,結果顯示與野生型相比,Sap30基因缺失的LSK 對受體小鼠外周血重建能力無顯著差異(P>0.05;圖 2I~J)。HSC 移植后的歸巢檢測顯示,與野生型相比,Sap30基因的缺失并不影響HSC的歸巢(P>0.05;圖2K)。
3 Sap30 基因缺失促進5-FU 處理后造血干/祖細胞再生
通過對Sap30-/-組和對照組小鼠進行腹腔5-FU(150 mg/kg)注射,建立5-FU 誘導的化療模型。在5-FU 注射后第6 天和第12 天犧牲小鼠,分析骨髓中造血干/祖細胞的數目(圖3A),分析結果顯示,Sap30基因缺失可顯著增加5-FU 處理后HSC 和MPP 的恢復(P<0.05;圖3B~3D)。本實驗所使用的基因缺失小鼠為全身敲除模型,為了排除其它組織器官中Sap30缺失對骨髓HSC 的影響,我們通過全骨髓移植構建了嵌合小鼠模型(骨髓為WT 或者KO,環境均為WT),對嵌合小鼠進行5-FU 處理(圖3E)。結果顯示,Sap30缺失仍可以增加骨髓中造血干/祖細胞的數目和在全骨髓細胞中所占比例(P<0.05;圖3F~3G),也可以顯著增加血液中白細胞的數量(P<0.05;圖3H),促進5-FU處理后血液體統重建。
4 Sap30基因缺失增強HSC體外擴增能力
構建體外再生模型,分別對野生型和Sap30-/-小鼠骨髓中分選出來的HSC(圖4A)進行了體外培養。體外培養7 d 后分析結果顯示,Sap30基因缺失促進HSC數目維持(P<0.05;圖4B)。
討 論
化療作為目前殺傷惡性腫瘤細胞的重要手段,在殺傷腫瘤細胞的同時也會抑制正常HSCs 的功能,在進行化療后HSC需要迅速向下游分化來恢復造血系統的穩態。在這一過程中如改善HSCs 的功能使造血系統盡快恢復重建,對于化療后病人健康狀態的恢復及免疫水平的提高存在重要的臨床意義[17]。改善小鼠HSCs 功能的手段眾多,如運動[18]和卡路里限制[19]等都能有效改善小鼠HSCs 的功能。在本研究中,我們應用基因缺失小鼠模型,以5-FU 模擬臨床的化療手段,開展了針對小鼠HSC 損傷后修復的研究。我們的研究顯示,Sap30基因的缺失在5-FU處理模擬的化療模型中能夠顯著提高HSCs 的再生能力,為提高化療后造血系統功能的恢復提供了干預靶點。HSCs 移植對眾多疾病的治療具有廣泛的應用價值,然而HSCs 有限的細胞來源及不成熟的體擴增/外再生的手段,極大地限制了供體來源的HSCs的數目[20]。近年來,越來越多的研究從組蛋白乙酰化修飾的角度開展了對HSC 體外擴增/再生的表觀遺傳調控機制研究,并證明許多脫乙酰酶抑制劑可以有效地促進HSCs 的體外擴增/再生。本研究中,應用小鼠模型及小鼠HSCs 體外培養實驗,結果顯示提示抑制脫乙酰酶輔酶的活性同樣能夠有效的促進HSC的體外擴增的作用。
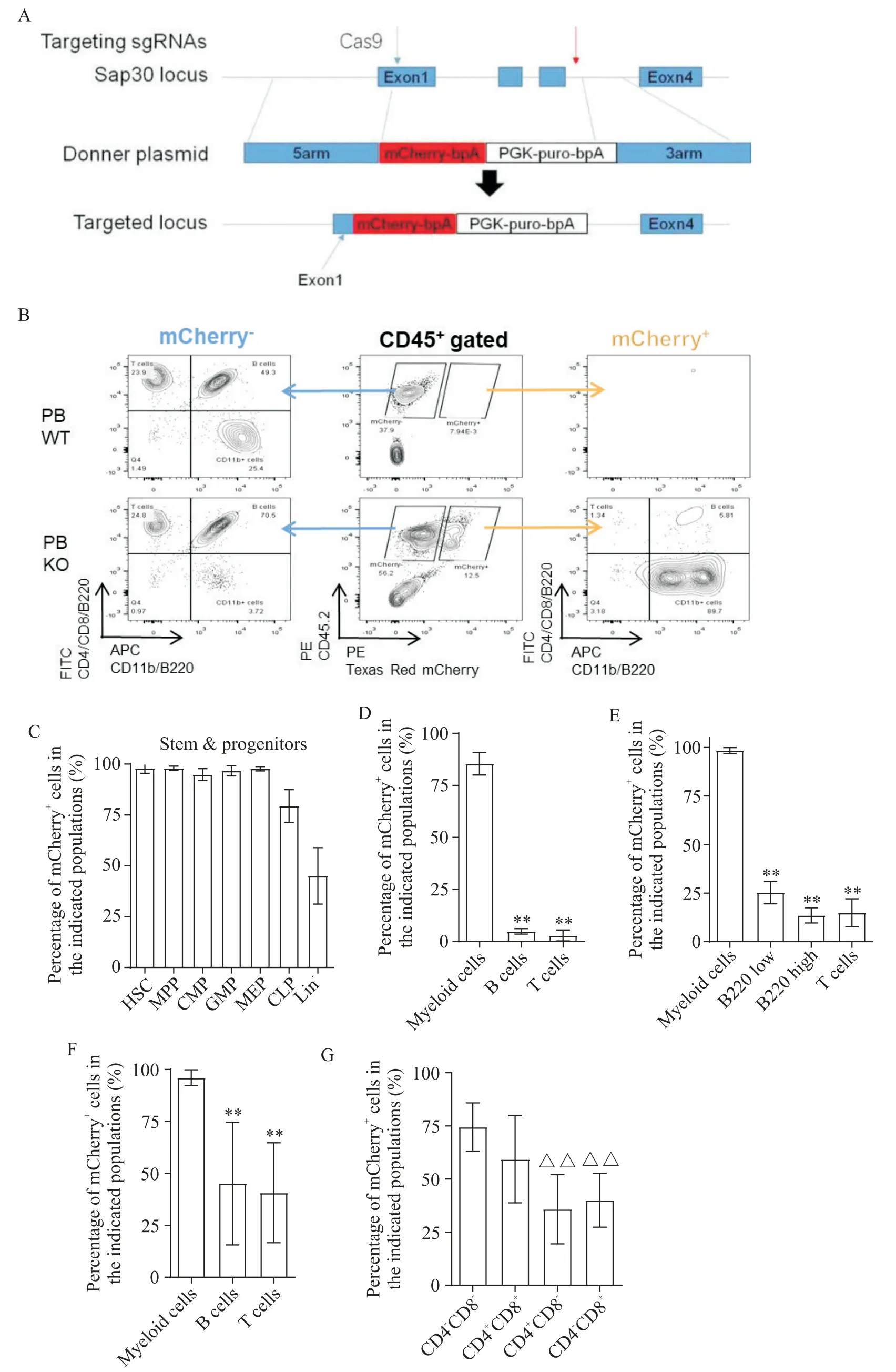
Figure 1. Sap30 was highly expressed in myeloid cells. A:strategies for constructing a Sap30-/- mouse model;B:FACS analysis showing the expression of Sap30 gene in peripheral blood(PB);C:the percentages of mCherry+ cells in stem/progenitor cells;D to F:the percentages of mCherry+ cells in B cells,T cells and myeloid cells in PB(D),bone marrow(E)and spleen(F);G:the percentages of mCherry+ cells in different T cells in thymus. Mean±SD. n=5 to 8.**P<0.01 vs myeloid cells;△△P<0.01 vs CD4-CD8-group.圖1 Sap30在髓系細胞中高表達
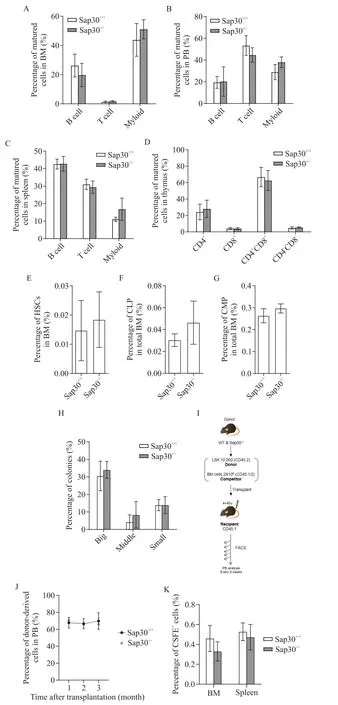
Figure 2. Sap30 was dispensable for maintaining hematopoiesis at steady stage. A to C:the percentages of B cells,T cells and myeloid cells in bone marrow(BM),peripheral blood(PB)and spleen of wild-type and Sap30 gene deletion mice;D:the percentages of different T cells in thymus of wild-type and Sap30 gene deletion mice;E to G:the proportions of hematopoietic stem cells(HSCs;E),common lymphoid progenitor(CLP;F)and common myeloid progenitor(CMP;G)in BM of wild-type and Sap30 gene deletion mice(n=5 to 8);H:the single HSC was sorted,and the size of colonies were measured(n=3 to 5);I:the experimental scheme of LSKs transplantation assay;J:the chimerism of PB of recipient mice was analyzed at 1,2 and 3 months after the transplantation(n=3 to 5);K:the percentages of CFSE-dilution on BM and spleen(n=3 to 5).圖2 Sap30基因缺失對小鼠造血系統的影響
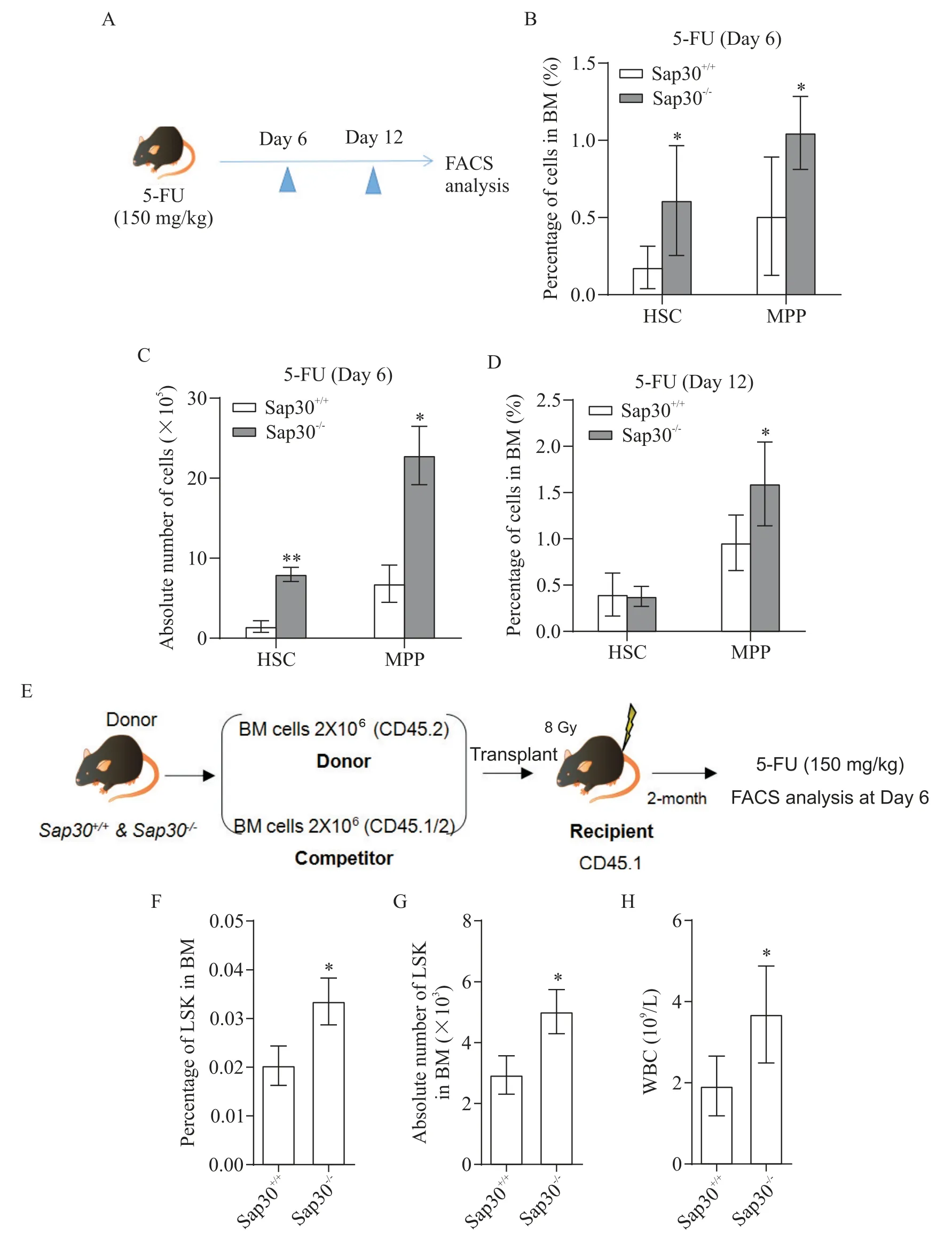
Figure 3. Sap30 deletion improved hematopoietic stem/progenitor cell regeneration in response to 5-FU treatment. A:the experimental scheme of 5-FU induction chemotherapy model;B and C:the percentages(B)and absolute numbers(C)of HSC and MPP in BM;C:of HSC and MPP after 5-FU injection on day 6;D:the percentages of HSC and MPP in BM after 5-FU injection on day 12;E:the experimental scheme of BM competitive transplantation assay;F and G:the percentages(F)and numbers(G)of LSK after 5-FU injection on day 6;H:the number of WBC. Mean±SD. n=3 to 4.*P<0.05,**P<0.01 vs Sap30+/+group.圖3 Sap30基因缺失促進了造血干/組細胞在5-FU處理后的再生
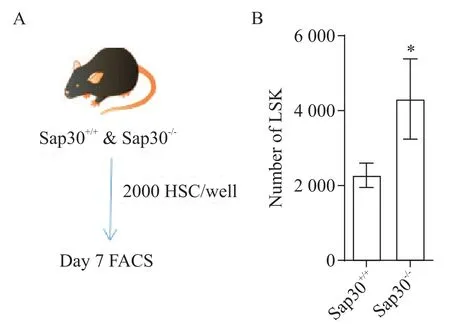
Figure 4. Sap30 deletion enhanced hematopoietic stem cell(HSC)ex vivo expansion. A:the experimental scheme of HSC ex vivo culture;B:the number of LSK was checked by FACS after 7 d. Mean±SD. n=4 to 5.*P<0.05 vs Sap30+/+group.圖4 Sap30基因缺失促進了造血干細胞的體外擴增
Sap30 蛋白作為脫乙酰酶的輔酶,調控組蛋白H3K27 乙酰化位點[11],我們觀察到它在髓系細胞中特異性表達,或可作為造血系統的細胞向髓系分化的表面標記,但其生理意義并不明確。同時我們觀察到穩態造血條件下Sap30的缺失并不會影響骨髓中各類細胞的比例,然而在應激或再生模型中Sap30的缺失增強了HSCs 的功能并促進其造血重建能力,提示Sap30在血液系統中的功能是條件依賴性的,應激環境可能激活了某類轉錄因子的表達從而招募乙酰化蛋白使得染色質中的某些特定區域發生乙酰化從而由關閉狀態到開放狀態進而調控HSC 的功能。由于Sap30調控H3K27 乙酰化位點,進一步明確Sap30在再生模型下的作用機制需要結合轉錄組學測序和H3K27乙酰化的ChIP-Seq來分析。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
