線粒體相關內質網膜調節神經元自噬的研究進展*
2022-11-01 03:25:34王一聃丁旭東陳俊文
中國病理生理雜志 2022年10期
關鍵詞:功能
王一聃, 曾 煉, 丁旭東, 桑 明, 陳俊文
(湖北醫藥學院附屬襄陽市第一人民醫院呼吸與危重癥醫學科,湖北 襄陽 441000)
神經元作為不可分裂的細胞,相比其他細胞壽命更長,在發育與維持過程中,需去除錯誤折疊的蛋白以及損傷衰老的細胞器,以保證生命活動的正常進行[1]。細胞自噬是細胞清除代謝物的主要途徑,其目的在于保證細胞對能量的需求以及內環境的穩定。自噬是維持正常神經功能所必須的,參與神經元的軸突運輸,突觸可塑性以及包括線粒體在內的細胞器的質量控制等過程[2-3]。當神經元自噬障礙時,隨著蛋白質與損傷細胞器的積累,會誘發神經退行性變[4];當神經元自噬過度激活時,也會繼發神經元的壞死,提示適度的自噬水平是神經元發揮正常功能的基礎。然而,目前對于神經元自噬發生的分子機制尚不清楚。越來越多的證據表明線粒體相關內質網膜(mitochondria-associated endoplasmic reticulum membranes,MAMs)在其中發揮重要作用。作為線粒體與內質網之間信號與物質交換的橋梁,MAMs具有廣泛的生物學功能,參與了Ca2+信號轉導,脂質代謝,線粒體動力學調控以及細胞自噬等[5]。通過對MAMs 中的蛋白構成進行蛋白質組學分析,檢測到大量調控細胞自噬的蛋白質(表1)[6-7]。同時MAMs 結構與功能的異常被證實參與神經退行性疾病的發病過程,而目前對于MAMs 在神經元自噬中的作用尚不清楚,本文匯總了MAMs 參與調節神經元自噬和神經退行性疾病的研究進展。
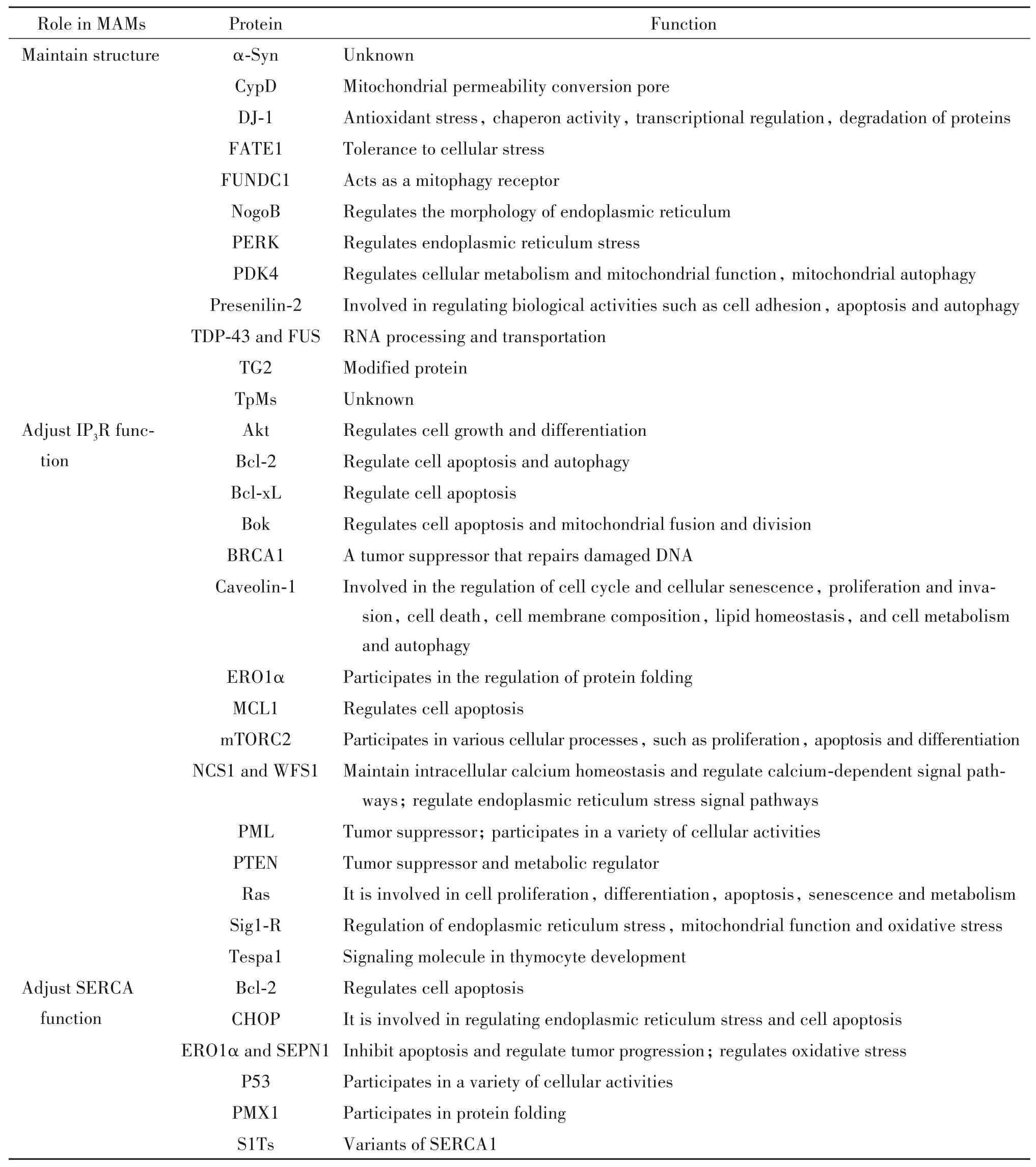
表1 MAMs中維持結構及發揮調節功能的蛋白[7]Table 1. Proteins in MAMs that maintaining structure and regulating functions
1 MAMs的構成與功能
真核細胞具有多種功能各異的膜結構細胞器,它們之間存在信號與物質的交換,而這種交換依賴于細胞器之間的特殊連接,其中MAMs 是典型的位于線粒體與內質網之間接觸的膜結構。MAMs 于上個世紀50 年代首次被觀察到,后續的研究表明大約5%~20%的線粒體外膜與內質網相互關聯[8]。從結構上來看,MAMs的厚度即線粒體外膜與內質網間的距離約為10~25 nm,其中粗面內質網與線粒體間的MAMs 厚度更大[9]。除厚度外,MAMs 形態也各具差異,根據內質網小管包繞線粒體的比例將MAMs 分為3型,其中1型包繞比例約占10%,2型則近乎完全包繞,3型僅包繞50%的線粒體,大多數MAMs屬于1型[10]。從功能上來看,MAMs早期主要作為脂質合成的場所,其中首次檢測到的蛋白是參與脂質代謝的蛋白質,包括磷脂酰乙醇胺甲基轉移酶2(phosphatidylethanolamineN-methyltransferase 2,PEMT2)、磷脂酰絲氨酸合酶 1/2(phosphatidylserine synthase 1,PSS1/2)等,而這些蛋白也成為鑒定MAMs 可靠的標志性蛋白[11]。除此之外,MAMs主要的功能是參與調節鈣穩態,進一步調控三磷酸腺苷(adenosine triphosphate,ATP)合成,線粒體分裂和細胞死亡等生命活動。而這種作用主要依賴于IP3Rs/GRP75/VDAC1 信號途徑實現[5]。作為包繞線粒體的膜結構,MAMs 參與調節線粒體的運動與分裂。線粒體具有沿微管運動的特性,受Ca2+調節,故MAMs 通過調控鈣轉運來影響線粒體運動[12]。同時,MAMs 也調控線粒體分裂。其中發動蛋白相關蛋白1(dynamin-related protein,DRP1)由胞質轉運至線粒體作為驅動線粒體分裂的關鍵環節[13]。研究證實 DRP1 在 MAMs 中表達,同時還包括兩種驅動DRP1線粒體轉運的其他蛋白,分別是線粒體分裂因子(mitochondrial fission factor)和分裂蛋白 1(fission protein 1)[14],因此 MAMs 可通過調節它們的表達與轉運進而調控線粒體分裂。
2 神經元自噬的概述
細胞自噬從形成活化的Unc-51樣自噬激活激酶1 復合體開始,在級聯反應作用下,激活beclin-1,參與合成Ⅲ類磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)復合體,驅動自體吞噬。形成的自噬小體經由自噬相關蛋白ATG12-ATG5 以及LC3-Ⅱ(ATG8-Ⅱ)構成的復合體調控,進而驅動自噬體的成熟[15]。成熟的自噬體與溶酶體連接,降解細胞中多余的成分。依據識別的特異性,自噬被分為選擇性自噬和非選擇性自噬。其中非選擇性自噬以饑餓誘導的自噬為代表,目的在于細胞組分的再利用[16],而選擇性自噬具有一個或多個受體介導的識別過程,該受體通過與LC3 相互識別將待降解物與自噬體連接,促進自噬體包繞[15]。依據自噬發生部位的不同,神經元自噬分為軸突自噬,樹突自噬和胞體自噬[3,17]。其中軸突中的自噬較為多見,神經元自噬體由軸突中產生后往胞體運輸,而這種運輸是在驅動蛋白作用下沿微管雙向進行的,最終在胞體中代謝清除[18]。除軸突自噬外,樹突中也檢測到了細胞自噬,其作用主要是調節神經元的興奮性,當使用低劑量N-亞硝基二甲胺刺激原代大鼠海馬神經元后,可觀察到樹突棘中自噬體數量的增多[19]。相比軸突和樹突,胞體中的自噬相對較少,且自噬體的成熟程度也較低[17]。胞體自噬主要目的是去除損傷及衰老的線粒體即線粒體自噬(mitophagy),進一步維持神經元線粒體結構與功能的完整[20]。依據發生時間的不同,神經元自噬又可分為發育期的自噬和成熟期的自噬。自噬是神經元發育所必須的,抑制胚胎期神經元的自噬可以導致皮層中神經元數量的減少[21]。同時自噬還可以調節神經元軸突生長,以及突觸的形成等發育過程[22-23]。對于成熟的神經元,自噬的主要作用是維持神經元的內穩態,而自噬功能異常則是大多數神經退行性疾病發生的重要基礎[24]。
3 MAMs調節神經元自噬
MAMs 對于神經元自噬的調節作用主要體現在以下幾方面:(1)對來源神經元的MAMs 中所含蛋白質進行蛋白質組學分析,檢測到大量與自噬相關或調節自噬的蛋白[6-7]。其中包括線粒體融合蛋白2(mitofusin 2,MFN2),MFN2 表達上調時,可削弱神經元中線粒體與內質網間的相互作用,干擾線粒體的正常功能,并促進MAMs 中的鈣轉運,轉運至線粒體的Ca2+通過激活DRP1 促進線粒體分裂以及PTEN誘導的假定激酶1(PTEN-induced putative kinase 1,PINK1)介導線粒體自噬,從而誘發神經退行性疾病[25]。除MFN2 外,磷酸呋喃酯酸性簇分選蛋白2(phosphofurin acidic cluster sorting protein-2,PACS-2)也被證實表達于MAMs 中,其功能主要是參與調節MAMs 的結構和細胞自噬。當下調神經元中PACS-2的表達時,除了影響MAMs 結構的完整外,還會導致脂質合成功能障礙,從而阻止LC3 與磷脂酰乙醇胺(phosphatidylethanolamine,PE)的偶聯抑制細胞自噬[26]。(2)MAMs參與自噬體的形成,被認為是神經元自噬的潛在起源。自噬體的發生受自噬相關蛋白的調控,神經元MAMs 中被證實含有大量自噬相關蛋白,包括ATG14 和ATG5 等,當自噬被激活時,MAMs中表達的ATG14L,ATG5 和含雙FYVE 結構的蛋白1(double FYVE-containing protein 1,DFCP1)的含量增多,在它們協同作用下,驅動細胞自噬[27]。相反的,當抑制MAMs 中的蛋白表達時,可以檢測到神經元中自噬體數量的減少[28]。(3)MAMs 通過干預脂質合成調控神經元自噬,LC3 與PE 偶聯被認為是自噬體成熟的標志,而PE 主要是通過內質網合成后轉運至MAMs 進行加工分泌,在此過程中受MAMs 的調控,故MAMs 可以通過影響PE 的合成來參與調控神經元自噬[29]。(4)MAMs 通過調節 Ca2+轉運來調控神經元自噬,Ca2+作為調控神經元生命活動的重要信號分子,也參與了介導神經元自噬的發生尤其是線粒體自噬,當神經元線粒體功能障礙時,由內質網經MAMs 轉運至線粒體的Ca2+誘發線粒體鈣超載,導致線粒體膜電位的降低,進一步激活線粒體自噬。而當破壞MAMs 介導的Ca2+轉運時,會導致AMPK 易位至MAMs 中進一步誘導神經元發生自噬性壞死[30]。總之,MAMs對于神經元自噬的調節主要基于其物質與功能的共同作用。
4 MAMs調控神經元線粒體自噬
線粒體自噬是損傷線粒體通過選擇性自噬途徑轉運至溶酶體中進行代謝清除的過程,其目的在于維持線粒體質量的穩定。線粒體自噬需要受體介導,其中主要的信號途徑即PINK1/parkin 途徑,當線粒體損傷時,聚積于線粒體中的PINK1 招募parkin并磷酸化激活它,parkin 活化后泛素化修飾p62/SQSTM1等受體蛋白,介導線粒體自噬[31]。線粒體自噬對于維持神經元正常的生理功能具有重要意義,同時也參與了眾多神經病理性過程。無論是腦損傷還是神經退行性疾病,線粒體自噬功能障礙被認為是疾病發生的重要基礎[32-33],尤其在神經退行性疾病中,由于神經元中積累著大量錯誤折疊的蛋白質,他們一方面抑制線粒體自噬來逃脫降解,另一方面過度激活線粒體自噬損害功能正常的線粒體,最終造成神經元的死亡[33]。MAMs作為線粒體外膜的延續,也參與調控神經元線粒體自噬。其中PINK1 被證實在神經元MAMs中有表達,同時兼具募集parkin的功能,當線粒體功能障礙時,儲存于MAMs 中的PINK1可以轉運至線粒體中招募并激活parkin,驅動線粒體自噬[34]。除 PINK1/parkin 途徑外,MAMs 還可通過影響含FUN14 結構域蛋白1(FUN14 domain-containing 1,FUNDC1)的表達調控線粒體自噬。FUNDC1主要參與缺氧條件下誘導的線粒體自噬,與PINK1/parkin途徑不同的是,FUNDC1 可以直接與LC3 結合,但這個過程受到氧氣的調控[35]。除直接激活FUNDC1外,MAMs 通過促進 FUNDC1 與 IP3Rs 的相互作用來進一步增加MAMs 中Ca2+轉運影響線粒體自噬[14]。除了調節 Ca2+轉運外,MAMs 中的 FUNDC1 對 MAMs本身也有作用,當抑制FUNDC1 表達時,會導致MAMs 的蛋白豐度減少包括參與調節細胞自噬的組分,從而干擾其對于神經元自噬的調控[36]。
5 MAMs相關的神經元自噬與神經退行性疾病
MAMs 結構與功能的異常被證實參與神經退行性疾病的發病過程。MAMs 主要通過調控β-淀粉樣蛋白(amyloid β-protein,Aβ)和α-突觸核蛋白(αsynuclein,α-Syn)等錯誤折疊蛋白合成、神經元鈣穩態、神經元線粒體功能、脂質代謝、神經元自噬等參與神經退行性疾病的發生過程[37]。由于MAMs 中富含乙酰輔酶A 乙酰轉移酶1(acetyl-CoA acetyltransferase 1,ACAT1)和膽固醇,兩者都是調節自噬的重要分子,因此MAMs 被認為可通過增強細胞自噬,調控阿爾茨海默病(Alzheimer disease,AD)患者和動物模型中 Aβ 和 tau 蛋白的清除[38]。此外,神經元線粒體功能障礙是AD 發生的重要基礎,其通過增加MAMs 介導的活性氧的合成,誘導內質網應激,進而減少自噬體形成,促進tau蛋白的沉積[39]。除AD 外,MAMs 中還存在大量調控線粒體自噬的相關蛋白包括PINK1/parkin,其中parkin 的突變被證實是常染色體陰性遺傳帕金森病患者發病的重要原因,其表達異常會誘導線粒體腫脹,引起線粒體損傷,最終導致黑質-紋狀體中多巴胺能神經元進行性減少并伴隨著 α-Syn 的沉積[28]。由 MAMs 蛋白功能障礙導致內質網-線粒體接觸失調通過影響神經元自噬活性從而介導肌萎縮側索硬化癥(amyotrophic lateral sclerosis,ALS)的發生,其中多個參與調節ALS 的關鍵基因 包 括SQSTM1、OPTN、UBQLN2和VCP等 都 在MAMs 中有所表達[40]。總之,MAMs 調控神經元自噬進而調節神經退行性疾病依賴于其內容物成分的變化,主要體現在調節自噬相關蛋白的表達發生改變。
6 總結與展望
自噬維持神經元內穩態,同時也是神經元發育和發揮神經功能的基礎。MAMs 作為內質網與線粒體間的橋梁,通過多種途徑介導神經元自噬(圖1):一方面,它充當了自噬相關蛋白執行功能的平臺,參與自噬體的形成與成熟;另一方面通過調控Ca2+轉運,脂質代謝影響自噬體的結構與功能。總的來說,MAMs 對于神經元自噬的調節是基于其物質與功能的聯合作用。MAMs 被揭示參與神經退行性疾病的發生,同時大多數神經退行性疾病都涉及到神經元自噬功能的異常。雖然目前已有研究揭示了MAMs參與調控細胞自噬,甚至被認為是自噬發生的潛在起源,但仍有許多問題要我們進一步的探索。MAMs究竟是通過表達哪些蛋白參與調節神經元自噬體的形成與成熟?這個過程是否還受到其他因子的調控?它們是否與MAMs的其他功能包括Ca2+轉運,脂質代謝之間存在相互關聯?MAMs 結構與功能的異常,神經元自噬的異常,神經退行性疾病三者之間的聯系?隨著技術的不斷發展,MAMs的成分和功能被深入挖掘,其在神經元自噬過程中的作用會被進一步闡明,有望成為治療神經退行性疾病的有效靶點。
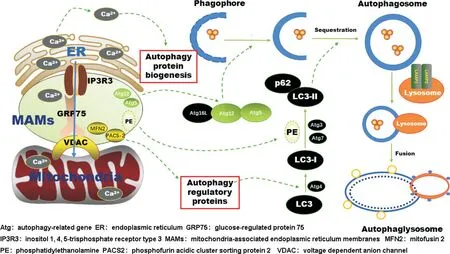
Figure 1. Schematic diagram of MAMs regulating neuronal autophagy. MAMs are involved in the genesis and maturation of autophagosomes by regulating calcium ion transduction,biosynthesis of autophagy-related proteins,and the expression of autophagyregulatory proteins.圖1 MAMs調控神經元自噬示意圖
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
