slfn5基因在卵巢癌上皮-間質轉化中的作用及機制
2022-11-07 04:50:20徐巧萍鄧魁林青青張珍
浙江醫學 2022年19期
徐巧萍 鄧魁 林青青 張珍
卵巢癌是女性生殖道最常見惡性腫瘤之一,其發病率和病死率逐年升高。據統計,2015年中國約有52 100例新發卵巢癌病例,病死率高達50%[1-2]。上皮-間質轉化(epithelial-mesenchymal transition,EMT)指上皮到間質細胞的轉化,它賦予細胞轉移和入侵的能力,在促進癌癥獲得轉移能力的過程中起重要作用。EMT的分子機制相當復雜,轉化生長因子(transforming growth factor,TGF)-β/母體抗生物皮膚生長因子同源蛋白(drosophila mothers against decapentaplegic protein,SMAD)、非受體酪氨酸激酶(Janus kinase,JAK)2/信號轉導及轉錄激活因子(signal transducer and activator of transcription,STAT)3、Notch、Ras-促分裂素原活化蛋白激酶(Ras-mitogen-activated protein kinase,Ras-MAPK)、NF-κB和p38等多條分子信號通路均被報道參與調控卵巢癌的EMT過程[3-4]。因此,尋找阻斷EMT的信號通路對抑制卵巢癌的侵襲、轉移具有重要意義。人類slfn(Schlafen)基因家族有6個成員,其中slfn5基因在肺癌、大腸癌、乳腺癌、胰腺癌等多種腫瘤組織中表達升高,通過激活蛋白激酶(protein kinase,PK)B/糖原合成酶激酶(glycogen synthase kinase,GSK)-3β/β-鏈環蛋白(β-catenin)通路、抑制基質金屬蛋白酶(matrix metalloproteinase,MMP)14表達來抑制細胞遷移和侵襲[5-6]。Fischietti等[7]推斷人類胰腺導管腺癌(pancreatic ductal adenocarcinoma,PDAC)患者的預后較差與slfn5基因高表達有關,這暗示slfn5基因還參與了PDAC的病理生理,并導致預后不良。對人肺癌細胞系A549的研究發現,slfn5基因過表達導致EMT相關轉錄因子Snail的水平升高,伴隨E-鈣黏蛋白(E-cadherin)表達降低,β-catenin從膜向細胞質或細胞核易位[8],可見slfn5基因是EMT過程中的關鍵調控分子。深入剖析slfn5基因上下游調控信號通路,探究癌細胞轉移機制,可為抑制轉移提供新的潛在治療靶點。本研究以人卵巢癌細胞系為材料,通過沉默slfn5基因,研究slfn5基因在卵巢癌EMT中的作用及其對腫瘤侵襲、遷移能力的影響,探討EMT標記蛋白和典型轉錄因子在卵巢癌發生、發展中的作用及機制,以期為卵巢癌治療提供新線索。
1 材料和方法
1.1 材料
1.1.1 組織材料 收集2019年8月至2022年4月在杭州市第一人民醫院行手術治療的21例卵巢癌患者的癌組織標本作為卵巢癌組。納入標準:手術病理檢查確診為卵巢癌且為首次確診;術前未進行放化療、靶向治療。排除標準:合并其他惡性腫瘤;合并子宮肌瘤、子宮內膜異位癥、子宮腺肌病、盆腔炎性疾病;合并重要臟器功能障礙;合并凝血功能障礙、傳染性疾病和感染性疾病。另收集同期行良性卵巢囊腫切除的14例患者的正常卵巢組織(距腫瘤組織邊緣>5 cm,經病理檢查確認)作為正常組。臨床樣本正常組年齡38~76(53.22±10.11)歲,卵巢癌組年齡39~75(52.73±12.04)歲。兩組年齡比較差異無統計學意義(P>0.05)。本研究經杭州市第一人民醫院醫學倫理委員會審核批準,所有患者均知情同意并簽署知情同意書。
1.1.2 細胞培養與分組 人卵巢表面上皮細胞IOSE80和卵巢癌細胞系 A2780、SKOV3、OVCAR3、HO8910均購于浙江美森細胞科技有限公司。所有細胞均使用專用培養基,置于37℃、5%二氧化碳培養箱中培養,隔天換液,待培養瓶底部鋪滿細胞時傳代,取生長狀態良好的對數生長期細胞進行后續實驗。
1.1.3 主要試劑 Real-time反轉錄試劑盒購自日本TaKaRa公司,Trizol試劑、Real-time PCR檢測試劑盒購自美國Promega公司;DEPC水購自北京索萊寶公司;細胞裂解液試劑盒購自美國Thermo公司;十二烷基硫酸鈉(SDS)-聚丙烯酰胺凝膠電泳(polyacrylamide gel electrophoresis,PAGE)試劑盒、二喹啉甲酸(bicinchoninic acid,BCA)蛋白濃度測定試劑盒購自上海碧云天生物技術有限公司;抗SLFN5抗體(批號:6789P)和N-鈣黏蛋白(N-cadherin,批號:14215S)、E-cadherin(批號:3195P)、波形纖維蛋白(Vimentin,批號:9585P)、Snail(批號:3879S)、微管蛋白(Tubulin,批號:sc-47778)抗體均購自美國CST公司,β-catenin抗人多克隆抗體購自武漢三鷹生物技術有限公司;辣根過氧化物酶(horseradish peroxidase,HRP)標記的抗羊IgG、抗兔IgG購自美國CST公司。LipofectamineTM2000轉染試劑盒購自上海賽默飛世爾科技有限公司;丙烯酰胺、雙丙烯酰胺、二甲基亞砜(DMSO)和磺酰羅丹明B購自美國Sigma公司。
1.2 兩組樣本組織中slfn5 mRNA表達的檢測 將卵巢癌組織和癌旁組織經液氮速凍后剪碎研磨成粉末,采用Trizol法抽提總RNA,采用熒光定量PCR(qRTPCR)法檢測mRNA擴增情況。按照逆轉錄試劑說明書配置逆轉錄體系,其中RNA模板1 μl,逆轉錄試劑 14 μl,用 DEPC 水補齊 20 μl。反應條件為 95 ℃預變性 15 min,94 ℃變性30 s,58℃退火 30 s,72℃延伸30 s,40個循環。每個樣品設置3個平行反應孔,實驗重復3次,以甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)為內參,采用 2-ΔΔCt法測定slfn5 mRNA的相對表達量。slfn5引物:正向5'-CATCCGACGCATCACCGATCTG-3',反 向 5'-CATCCG ACGCATCACCGATCTG-3';GAPDH引物:正向5'-TGAACGGGAAGCTCACTGG-3',反向 5'-TCCACCACCCTGTTGCTGTA-3'。引物由上海生工生物工程公司合成。
1.3 細胞實驗及指標測定
1.3.1 5種細胞中SLFN5蛋白表達的檢測 采用Western blot法。將5組細胞分別加入細胞裂解液,冰上裂解30 min,離心收集上清液。采用BCA蛋白檢測試劑盒測定蛋白水平。樣品加入上樣緩沖液,根據SDS-PAGE試劑盒方法分離樣品,將條帶轉移到聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜上,加入5%脫脂奶粉封閉1 h,洗膜后加入抗SLFN5一抗孵育過夜。再次洗膜后加入HRP二抗,孵育1 h。洗膜后加入化學發光增強劑,自顯影。采用圖像分析處理系統(Image J軟件)對蛋白條帶掃描分析,用測得的蛋白光密度值與Tubulin光密度值的比值表示5組細胞中SLFN5蛋白的相對表達量。
1.3.2 細胞分組、轉染及sfln5 mRNA表達的檢測 取上述SLFN5蛋白相對表達水平高的2個細胞系用于后續實驗。由上海吉凱公司設計并合成特異性沉默slfn5的RNA(si-slfn5#1、si-slfn5#2)及陰性對照(siRNANC),采用LipofectamineTM2000轉染試劑盒方法依次轉染,即為si-slfn5#1組、si-slfn5#2組和si-NC組,繼續培養24 h;si-slfn5#1組、si-slfn5#2組引物序列見表1。取出細胞,采用qRT-PCR法檢測各組細胞slfn5 mRNA的相對表達量。

表1 slfn5引物的siRNA序列
1.3.3 細胞遷移能力和侵襲能力的測定 取轉染后的細胞作為材料。(1)細胞遷移實驗:Transwell小室的下室中加入500 μl含10%FBS的完全細胞培養基,上室接種300 μl不含FBS的細胞懸液(細胞密度約2×104/L)。培養24 h后,甲醇固定Transwell小室的下層細胞,結晶紫染色、清水沖洗,用棉簽輕輕拭去上層細胞,10倍顯微鏡下隨機選擇5個視野,觀察穿過小室濾膜的細胞并計數,細胞相對數量比越大表示細胞遷移能力越強。(2)細胞侵襲實驗:在Transwell小室的上室中預先均勻鋪上100 μl稀釋好的基底膠,37℃靜置約2 h,待其凝結成膠后,參照細胞遷移實驗進行,觀察穿過小室濾膜的細胞并計數,細胞相對數量比越大表示細胞侵襲能力越強。
1.3.4 轉染細胞遷移率的檢測 采用細胞劃痕實驗。取轉染后的各組細胞制備細胞懸液(約2×104/L),接種于6孔板。當細胞鋪滿6孔板底部時,用滅菌的移液槍槍頭垂直于板底均勻劃痕,無菌PBS洗去脫落細胞,于顯微鏡下觀察劃痕并拍照記錄;繼續培養24 h后再次取樣拍照。用Image J圖像處理軟件并測定0 h(T0h)及24 h(T24h)劃痕面積,計算細胞遷移率。細胞遷移率=(T0h-T24h)/T0h×100%。
1.3.5 轉染細胞中EMT相關蛋白表達的檢測 采用Westetn blot法。取各組轉染細胞加入細胞裂解液,采用BCA蛋白檢測試劑盒測定蛋白水平。樣品加入上樣緩沖液,根據SDS-PAGE試劑盒方法分離樣品,將條帶轉移到PVDF膜上,加入5%脫脂奶粉封閉1 h,洗膜后分別加入 SLFN5、Vimentin、Snail、N-cadherin 和 E-cadherin等一抗,4℃孵育過夜,加入二抗后室溫孵育45 min,顯影并用Image J軟件進行灰度分析,以Tubulin為內參,計算細胞中Vimentin、Snail、N-cadherin 和E-cadherin等EMT相關蛋白的相對表達量。
1.3.6 轉染細胞中Wnt/β-catenin信號通路相關蛋白表達的檢測 采用Westetn blot法。取各組轉染細胞加入細胞裂解液,參照BCA蛋白濃度測定試劑盒方法測定蛋白濃度;SDS-PAGE分離條帶并轉模,加入5%牛奶室溫封閉1 h,洗膜后加入β-catenin一抗,4℃搖床孵育過夜,加入二抗室溫孵育45 min,顯影并用Image J軟件進行灰度分析,以Tubulin為內參,計算βcatenin蛋白相對表達量。
1.4 統計學處理 采用GraphPad Prism統計軟件。計量資料以表示,兩組間比較采用兩獨立樣本t檢驗,多組間比較采用單因素方差分析。P<0.05為差異有統計學意義。
2 結果
2.1 兩組樣本組織中slfn5 mRNA表達的比較 相比正常組,卵巢癌組slfn5 mRNA相對表達量明顯增高,差異有統計學意義(P<0.01),見圖1。
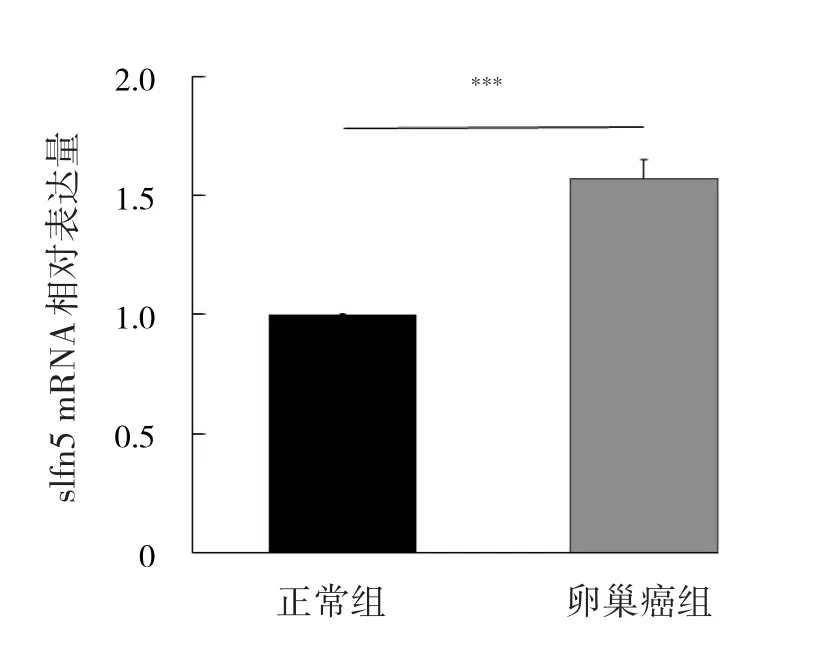
圖1 兩組樣本組織中slfn5 mRNA表達的比較
2.2 5組細胞中SLFN5蛋白表達的比較 與IOSE80相比,HO8910、SKOV3、OVCAR3細胞系SLFN5蛋白均顯著高表達,差異均有統計學意義(均P<0.01),其中SKOV3、OVCAR3相對表達水平更高,見圖2。故選取SKOV3、OVCAR3細胞系進行后續實驗。
2.3 轉染細胞中slfn5 mRNA表達的比較 與si-NC組相比,si-slfn5#1組、si-slfn5#2組細胞中slfn5 mRNA相對表達量均明顯降低,差異有統計學意義(均P<0.01),提示穩定沉默slfn5基因的SKOV3、OVCAR3細胞株構建成功,見圖3。
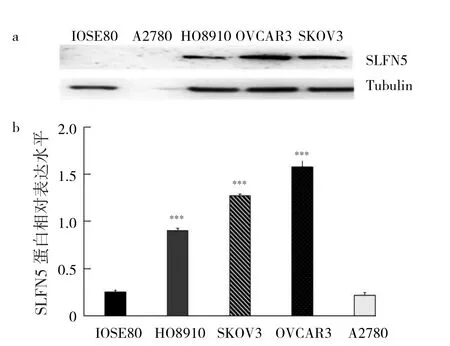
注:Tubulin為微管蛋白;與IOSE80比較,***P<0.001
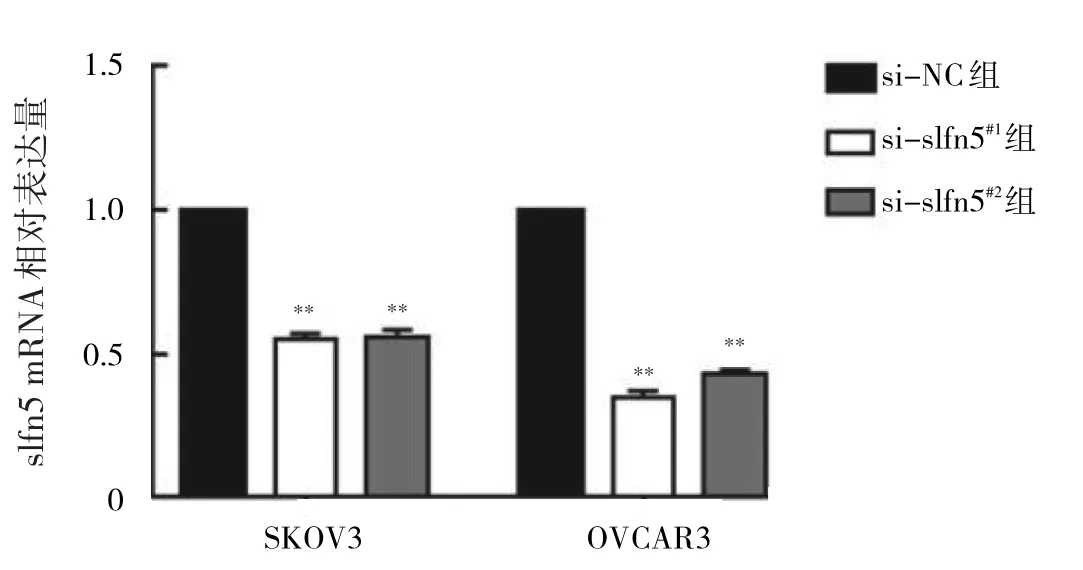
圖3 轉染細胞中slfn5 mRNA表達的比較
2.4 沉默slfn5基因對SKOV3、OVCAR3細胞遷移和侵襲能力的影響 Transwell遷移實驗結果顯示,相比si-NC組,si-slfn5#1組、si-slfn5#2組穿過基底膜的細胞相對數量比均減小,差異均有統計學意義(均P<0.01),提示沉默slfn5基因抑制了SKOV3、OVCAR3細胞的遷移。Transell侵襲實驗結果顯示,相比si-NC組,sislfn5#1組、si-slfn5#2組穿過基底膜的細胞相對數量比均減小,差異均有統計學差異(均P<0.01),提示沉默slfn5基因抑制了SKOV3、OVCAR3細胞的侵襲,見圖4。
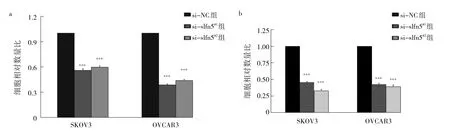
圖4 沉默slfn5基因對SKOV3、OVCAR3細胞遷移和侵襲能力的影響(a:遷移實驗的3組細胞相對數量比;b:侵襲實驗的3組細胞相對數量比)
2.5 沉默slfn5基因對SKOV3、OVCAR3細胞遷移率的影響 與si-NC組相比,si-slfn5#1組、si-slfn5#2組的細胞遷移率均顯著低于si-NC組,差異均有統計學意義(均P<0.01),驗證了沉默slfn5基因能夠降低SKOV3、OVCAR3細胞的遷移能力,見圖5。
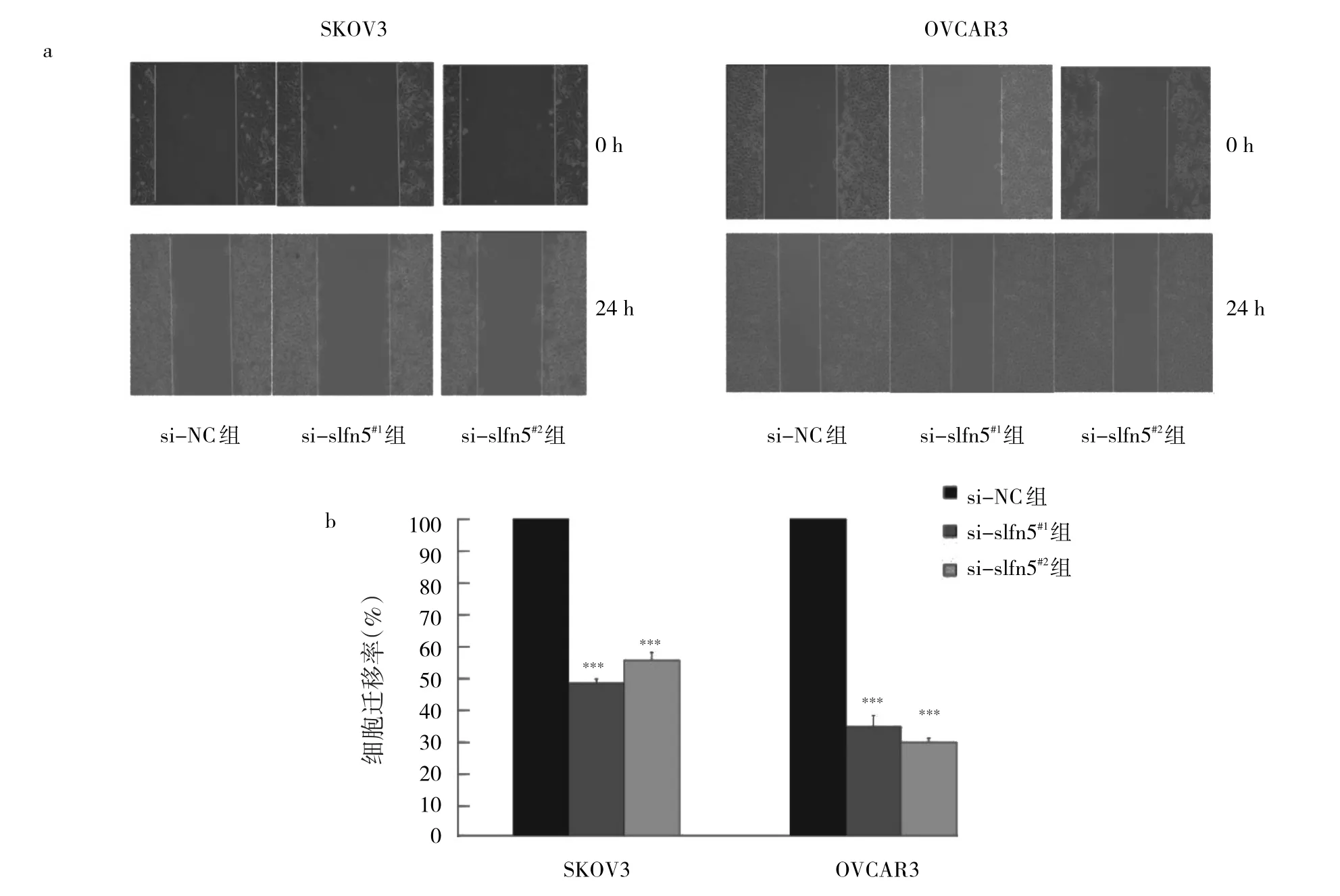
圖5 沉默slfn5基因對SKOV3、OVCAR3細胞遷移率的影響(a:SKOV3、OVCAR3細胞遷移圖;b:沉默slfn5基因后,SKOV3、OVCAR3細胞的遷移率)
2.6 沉默slfn5基因對SKOV3、OVCAR3細胞EMT相關蛋白表達的影響 Western blot檢測顯示,與si-NC組相比,si-slfn5#1組、si-slfn5#2組 Vimentin、Snail、N-cadherin等的相對表達量顯著降低,E-cadherin相對表達量顯著增加,差異均有統計學意義(均P<0.01)。E-cadherin的增加和Vimentin、Snail、N-cadherin的減少與卵巢癌細胞遷移和侵襲能力下降直接相關,提示沉默slfn5基因能抑制卵巢癌細胞發生EMT,見圖6。
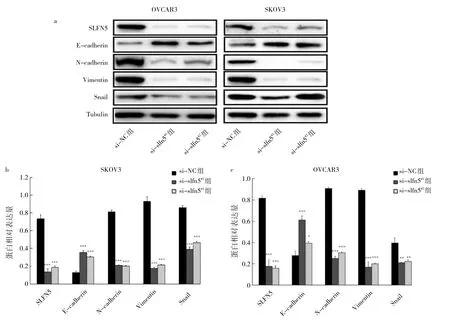
圖6 沉默slfn5基因對SKOV3、OVCAR3細胞EMT相關蛋白表達的影響(a:蛋白電泳圖;b-c:沉默slfn5基因的SKOV3、OVCAR3細胞EMT相關蛋白相對表達量)
2.7 沉默slfn5基因對SKOV3、OVCAR3細胞β-catenin表達的影響 與si-NC組相比,si-slfn5#1組、si-slfn5#2組β-catenin相對表達量明顯降低,差異均有統計學意義(均P<0.01),見圖7。β-catenin是Wnt/β-catenin信號通路關鍵調控因子,其相對表達量下降提示Wnt/βcatenin信號通路活性降低。
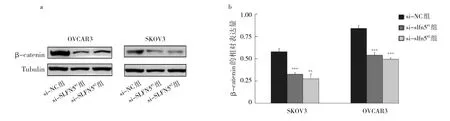
圖7 沉默slfn5基因對SKOV3、OVCAR3細胞β-catenin表達的影響(a:蛋白電泳圖;b:沉默slfn5基因的SKOV3、OVCAR3細胞βcatenin蛋白相對表達量)
3 討論
人類slfn(也有稱SLFN)家族基因于1998年首次被報道,在免疫系統和惡性腫瘤中起重要作用[1],并被確認在哺乳動物系統和器官中廣泛表達[9-10]。slfn5基因被認為與幾種惡性腫瘤的細胞侵襲和轉移密切相關,如Mavrommatis等[11]發現slfn5基因高表達可抑制惡性黑色素瘤的錨定非依賴性生長,而敲除slfn5基因則可以增強惡性黑色素瘤細胞侵襲三維膠原的能力。Sassano等[5]研究發現,slfn5通過負調控MMP-1和MMP-13抑制腎癌細胞的遷移和侵襲。slfn5還被認為是一種乳腺癌抑制因子,可通過直接調節乳腺癌轉錄因子——鋅指E盒結合同源框(zinc-finger E-box binding homeobox,ZEB)蛋白1的表達來實現抗癌作用[12]。但slfn5基因對卵巢癌發生、發展的作用還未見報道。本研究檢測了23例患者的卵巢癌組織和癌旁組織中slfn5 mRNA的表達水平,結果發現相對于癌旁組織,卵巢癌組織中slfn5 mRNA表達量顯著上調。相比卵巢表面上皮細胞,卵巢癌HO8910、SKOV3、OVCAR3細胞系均表現出SLFN5蛋白的高表達,證實了卵巢癌細胞和癌組織中存在slfn5基因表達上調。
腫瘤遷移和侵襲是兩個密切相關的病理過程,與腫瘤轉移息息相關。卵巢癌轉移是影響卵巢癌治療、預后以及復發的重要因素,因此,對卵巢癌細胞遷移和侵襲的機制研究是卵巢癌研究的重中之重。slfn5基因被證實在控制腎細胞癌(renal cell carcinoma,RCC)細胞的運動性和侵襲性方面發揮關鍵作用[11]。slfn5基因表達促進腦膠質瘤細胞的運動性和侵襲性[13]。本研究表明,沉默slfn5基因可以顯著抑制卵巢癌的遷移和侵襲能力。對轉染沉默slfn5基因的卵巢癌細胞的劃痕實驗、Transwell實驗結果證實下調slfn5基因抑制了卵巢癌細胞的遷移能力和侵襲能力。沉默slfn5基因后,SKOV3、OVCAR3細胞的遷移、侵襲能力被有效抑制,細胞遷移率顯著低于對照組,提示slfn5基因可能通過促進人卵巢癌細胞系的EMT實現細胞遷移和侵襲。
EMT最早是在胚胎發育中發現的一個現象,成體的表皮細胞受到損傷后,也會出現相應的EMT。腫瘤細胞在發生、發展中,常常通過EMT丟失部分上皮細胞的特性來獲得一些間質細胞的特性,從而獲得更強的侵襲能力[14]。EMT的主要特征包括E-cadherin等上皮標志物表達下降甚至缺失,而間質標志物如N-cadherin、Vimentin蛋白表達的增加[15]。本研究中,沉默slfn5基因后,卵巢細胞系E-cadherin蛋白水平上調,Vimentin、Snail和N-cadherin下調,提示slfn5基因在卵巢癌細胞EMT中起抑制作用。E-cadherin下調導致腫瘤細胞間的黏附性減弱,從而促進了腫瘤細胞的脫落,后者隨血液和淋巴轉移[3],被認為是上皮-間質細胞轉型的重要特征。N-cadherin是一種神經性鈣粘附蛋白,是細胞間動態黏附的重要結構成分,在惡性腫瘤細胞中的表達比正常上皮表型細胞高[16-17]。本研究發現,EMT相關蛋白的表達水平與slfn5基因和SLFN5蛋白的表達水平有關,slfn5基因是EMT過程中的關鍵調控因子,這為抑制卵巢癌轉移提供了新的潛在治療靶點。
β-catenin是一種多功能蛋白,通過與細胞骨架的相互作用,協助細胞對細胞外的信號和影響作出響應,其典型作用是通過Wnt/β-catenin信號通路激活核內靶基因的表達[18]。EMT過程中,β-catenin介導E-cadherin受體與細胞骨架的附著,該細胞骨架與膜上的E-cadherin結合,完成從膜到核的易位[19]。βcatenin定位于正常上皮細胞和非侵襲性腫瘤細胞的細胞膜中,與E-cadherin分離后在細胞質中積累并在EMT期間轉移至細胞核,以激活EMT相關基因的轉錄,特別是促進Snail和ZEB1等轉錄因子的表達[20]。本研究構建了沉默slfn5基因的卵巢癌細胞系,Western blot檢測發現,相比對照組,轉染組細胞中Snail、N-cadherin、Vimentin和 β-catenin顯著下調,E-cadherin顯著上調。Snail是E-cadherin的公認抑制劑,可直接抑制E-cadherin的轉錄并誘導EMT,Snail還可以通過誘導E-cadherin阻遏物(如ZEB1)的表達并上調間質標記Vimentin,從而導致上皮細胞黏附力的喪失,誘導EMT并促進細胞遷移,侵襲并最終轉移到遠處器官[21]。本研究中轉染細胞E-cadherin顯著高表達,Snail及其他EMT蛋白表達趨勢與β-catenin一致,與SLFN5表達趨勢一致,提示slfn5可能通過β-catenin-Snail-E-cadherin信號轉導來促進腫瘤轉移,即slfn5基因可能通過β-catenin介導的Snail/E-cadherin信號通路在EMT和卵巢癌細胞轉移中起作用[6]。
作為腫瘤細胞轉移的關鍵步驟,EMT在腫瘤的診斷、分期和治療中起著重要的作用。利用EMT過程中涉及的分子發現新的腫瘤治療靶標以及開發新的診斷和治療藥物可能成為癌癥早期診斷和治療的新策略[21]。本研究通過沉默slfn5基因調控EMT相關蛋白的表達,并抑制細胞遷移和侵襲,其機制可能是抑制βcatenin表達,調節了β-catenin介導的Snail/E-cadherin信號通路,從而降低Wnt/β-catenin信號通路活性。
