云南玉溪綠孔雀遺傳多樣性分析:與圈養種群的比較
2022-11-07 03:11:00劉思霞張馨元晏鳴霄杜焓瑜楊金城李廣龍官文淵楊守莊徐艷春
野生動物學報 2022年4期
劉思霞 張馨元 晏鳴霄 杜焓瑜 楊金城 李廣龍 官文淵 楊守莊 徐艷春,7*
(1.東北林業大學野生動物與自然保護地學院,哈爾濱,150040;2.中國科學院西雙版納熱帶植物園,勐臘,666303;3.浙江大學生命科學學院,杭州,310058;4.國家林業和草原局林產工業規劃設計院,北京,100010;5.衢州越鳥農業發展有限公司,衢州,324000;6.秦皇島野生動物園,秦皇島,066100;7.國家林業和草原局野生動物保護與利用工程技術研究中心,哈爾濱,150040)
綠孔雀(Pavomuticus)是一種大型雞形目(Galliformes)鳥類,曾在中國南方[1-2]和東南亞[3]廣泛分布,并在世界各地成為民族文化的重要載體。綠孔雀被分為3個亞種,即原產于馬來西亞和爪哇的指名亞種(P.m.muticus),分布于印度東北部至緬甸西北部的印度亞種(P.m.spicifer)和分布于印支地區的云南亞種(P.m.imperator)[4]。在過去的半個多世紀里,由于狩獵[5-6]、棲息地退化和喪失[5,7]以及人類干擾[8-9],所有分布區的綠孔雀種群數量都顯著下降,2009年綠孔雀被IUCN瀕危物種紅色名錄列為瀕危(EN)級(https://www.iucnredlist.org/species/22679440/131749282)。
歷史上,綠孔雀在中國的分布很廣,包括湖南、湖北、四川、廣西、廣東、云南和西藏等地理范圍[10],而今僅在云南省西部、中部和南部的22個縣有所分布[5]。1993年,實地調查記錄的種群總數量在800~1 100只[11],此后下降到大約500只[12],最新調查為235~280只[13]。我國境內的綠孔雀棲息地,除了占種群數量60%以上的雙柏和新平外,近60%的分布區域都在過去的30年里相繼消失,而雙柏和新平2個地區也面臨著棲息地破碎化的風險[5,14]。本研究借助云南省玉溪市新平和峨山交界處的羽毛樣本,分析該種群的遺傳多樣性,為深入了解綠孔雀種群狀況,制定保護計劃提供基本信息。
1 材料與方法
1.1 樣品采集
2018—2019年,在云南省玉溪市新平與峨山交界處的野外調查中,從自然棲息地共收集到29份羽毛樣品(圖1)。每份樣品單獨包裝在一個紙質信封里,標明收集地點和時間。羽毛樣本編碼為YBD01~YBD29,4 ℃保存,命名為YBD種群。另外,選擇9只純種綠孔雀(編碼GP01~GP09)和6只純種藍孔雀(編碼BP01~BP06)作為參考,以鑒定羽毛所屬物種。為了研究YBD種群與中國現有圈養種群之間的遺傳關系,從不同的養殖機構共采集了33份綠孔雀樣品,其中16只是從中國云南瑞麗捕獲的野生綠孔雀的后代,其余個體的地理來源不明。所有樣品均為羽毛樣品,編碼為CAP01~CAP33。
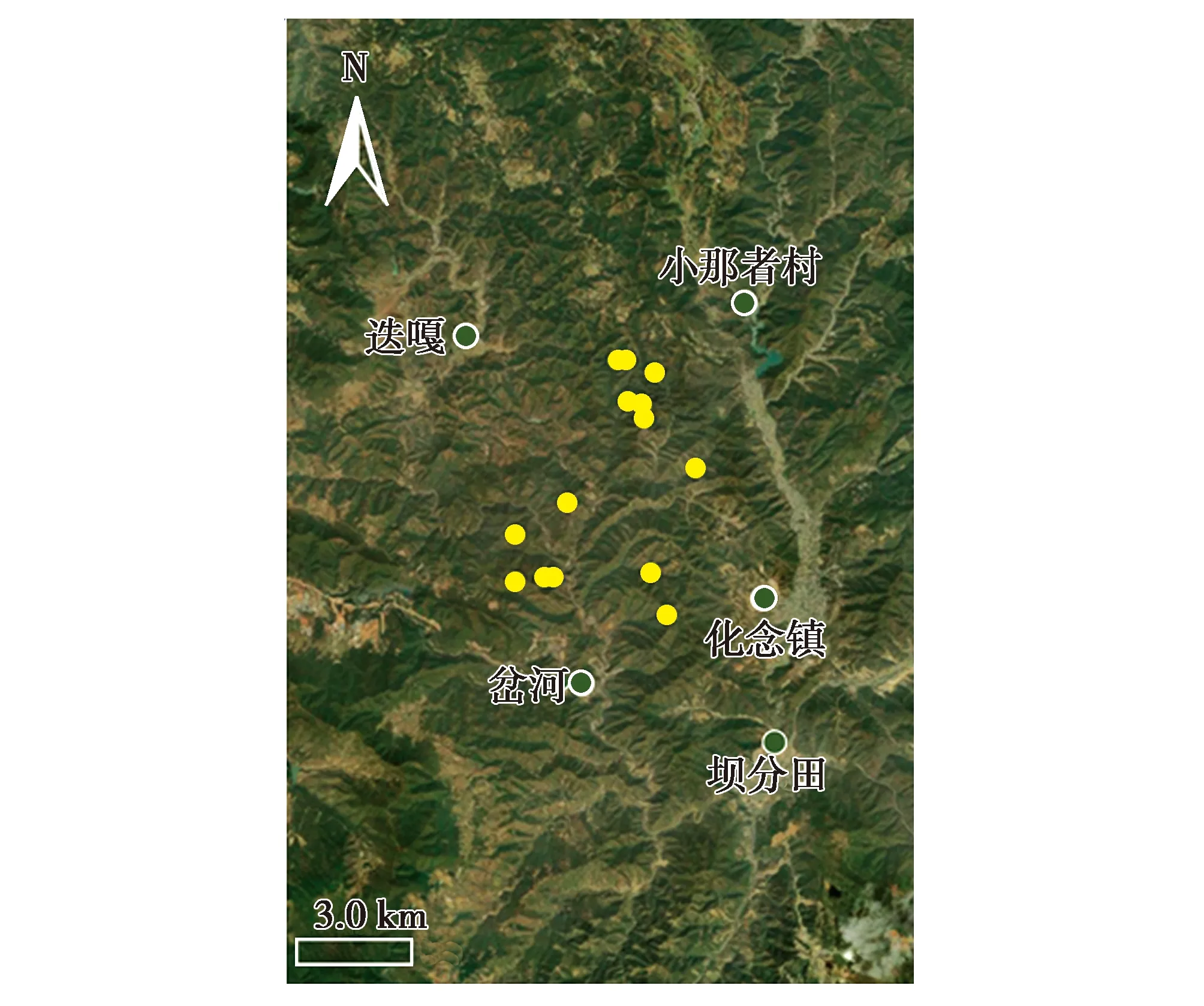
圖1 云南省玉溪市新平與峨山交界處孔雀羽毛的收集地點Fig.1 Geographic location of the collected feather samples in the juncture of Xinping and Eshan of Yuxi City,Yunnan Province,China
1.2 DNA提取
從每根羽毛上切下約5 mg的羽枝,放入離心管中。向試管中加入約1 000 μL 2% SDS溶液,把羽毛沒入液面以下,在65 ℃的水浴中孵育30 min。將羽毛樣品轉移到另一個含有75%乙醇的燒杯中漂洗30 min,再轉移到含有20 mL ddH2O的50 mL燒杯中,振蕩洗滌約30 min。清洗干凈的羽毛樣品放在濾紙上自然干燥,移入1.5 mL離心管中,剪成小塊備用。DNA的提取按照Volo等[15]的方法,消化時間為2 d,異丙醇沉淀冷凍12 h,冷凍離心16 000 r/min 30 min。使用Nanodrop 2000 C(Thermo Scientific,USA)測定DNA的濃度,并稀釋至~15 ng/μL用于PCR 擴增。
1.3 Cyt b基因片段的擴增和測序
Cytb基因 PCR 擴增引物參考Verma等[16]使用的鳥類常用擴增引物mcb398(5′-TACCATGAGGACAAATATCATTCTG-3′)和mcb869(5′-CCTCCTAGTTTGTTAGGGATTGATCG-3′)。采用50 μL的PCR反應體系:DNA(~45 ng)3 μL,上下游引物各1 μL(10 pmol/μL),2×EasyTaq?PCR SuperMix(全式金生物技術有限公司)25 μL和ddH2O 20 μL。反應程序:94 ℃預變性5 min;94 ℃變性30 s,52 ℃退火45 s,72 ℃延伸30 s,循環35次;72 ℃終延伸7 min,4 ℃保存。擴增產物在1.5%的瓊脂糖凝膠上80 V電泳40 min,用AxyPrepTMDNA Gel Extraction Kit試劑盒純化回收,采用Sanger法雙向測序,用DNASTAR軟件包上的SeqMan Pro對序列進行拼接[17]。
1.4 微衛星的分型
參考Du等[18]的方法,選擇PC3、PC19、PC25、PC36、PC41、PC46、PC67、PC125、PC142、PC148、PC151、PC201、PC236、PC281[19]、MCW0069和MCW0098[20]16個微衛星位點對野生和圈養種群的樣本分型,上游引物的5′端用FAM(藍色)、JOE(綠色)和TAMRA(黑色)3種熒光素標記。PCR反應體系均為15 μL:DNA(~45 ng)3 μL,上、下游引物(10 pmol/μL)各1 μL,2×EasyTaq?PCR SuperMix(全式金生物技術有限公司)5 μL和ddH2O 5 μL。反應程序:94 ℃預變性5 min;94 ℃變性30 s +Ta℃退火45 s + 72 ℃延伸30 s,循環35次;72 ℃終延伸10 min,4 ℃保存。PCR產物在3100x Genetic Analyzer(Applied Biosystems,USA)上電泳分離,用GeneMarker?收集片段信號[21]。為避免基因分型錯誤,每個個體進行3次基因分型。
1.5 羽毛樣品的母系物種鑒定
使用MEGA 5.05中的Clustal W程序[22],比較野外羽毛樣品和已知綠孔雀、藍孔雀的線粒體Cytb基因序列,確認它們的同源性,用EditSeq[17]將所有序列修剪到421 bp。根據Dinh等[23]的建議,用MEGA 5.05[22]和Kimura 2-parameter(K2P)模型計算每個待測羽毛樣本與已知純種綠孔雀和藍孔雀樣本之間的遺傳距離,同時計算已知綠孔雀種內兩兩個體間的遺傳距離(intra-d)與綠孔雀、藍孔雀種間兩兩個體間的遺傳距離(inter-d),這些遺傳距離用于確定每個樣本的母體物種起源。使用IBM SPSS Statistics 23(IBM Corp.New York,USA)檢驗每個羽毛樣品與2組已知孔雀個體的遺傳距離落入intra-d還是inter-d的范圍(95%置信區間),從而確定其所屬物種;用MEGA 5.05基于K2P距離構建系統發育樹,用iTOL v4.4.2(https://itol.embl.de/)進行可視化。
1.6 個體識別
羽毛樣本在野外撿拾,可能存在同一個體多次重復,因此在正式的種群分析之前,需進行個體識別以確定真實樣本量(鳥類數量)。首先,用Micro-Checker v2.23[24]檢測各個微衛星分型結果的無效等位基因的概率,據此矯正等位基因和基因型頻率,用于進一步種群遺傳分析。用Gimlet Software v1.3.1[25]計算多位點的累積個體識別概率(cumulative probability of identity,P(ID)),即2個無關個體或同胞之間在多個微衛星位點上基因型完全一致的可能性[26]。為了消除種群結構的影響,計算修正基因型相似概率(P(ID)biased);考慮到樣本中存在血緣關系的可能,還計算了同胞或親子之間基因型相似概率(P(ID)sibs),作為個體識別的最低可信度。最后,用Cervus 3.0確認被鑒定為同一個體的多個樣品是否在所有位點的基因型完全相同,或者是符合最低接受標準:僅在1個位點相差1個等位基因[27]。
1.7 遺傳多樣性及種群遺傳結構分析
使用Popgene 1.32計算觀察等位基因數(Na)、有效等位基因數(Ne)、觀察雜合度(Ho)和期望雜合度(He);用Cervus 3.0計算YBD種群和CAP種群的每個微衛星位點的多態信息含量(PIC);用Popgene 1.32計算2個種群的種群近交系數(fixation indices of subpopulation,FIS)、每個位點的遺傳分化指數(FST)、基因流(Nm,Nm=0.25(1-FST)/FST)、Nei氏遺傳距離和遺傳相似度,并進行哈迪-溫伯格平衡(H-W)檢驗。為了推斷YBD和CAP種群之間的遺傳分化程度,用Arlequin 3.5[28]計算2個群體之間的遺傳分化指數FST,使用STRUCTURE 2.3.1分析種群的遺傳結構[29]。最佳K值(理論種群數量)和種群遺傳結構圖形結果使用在線軟件(http://clumpak.tau.ac.il/)生成。
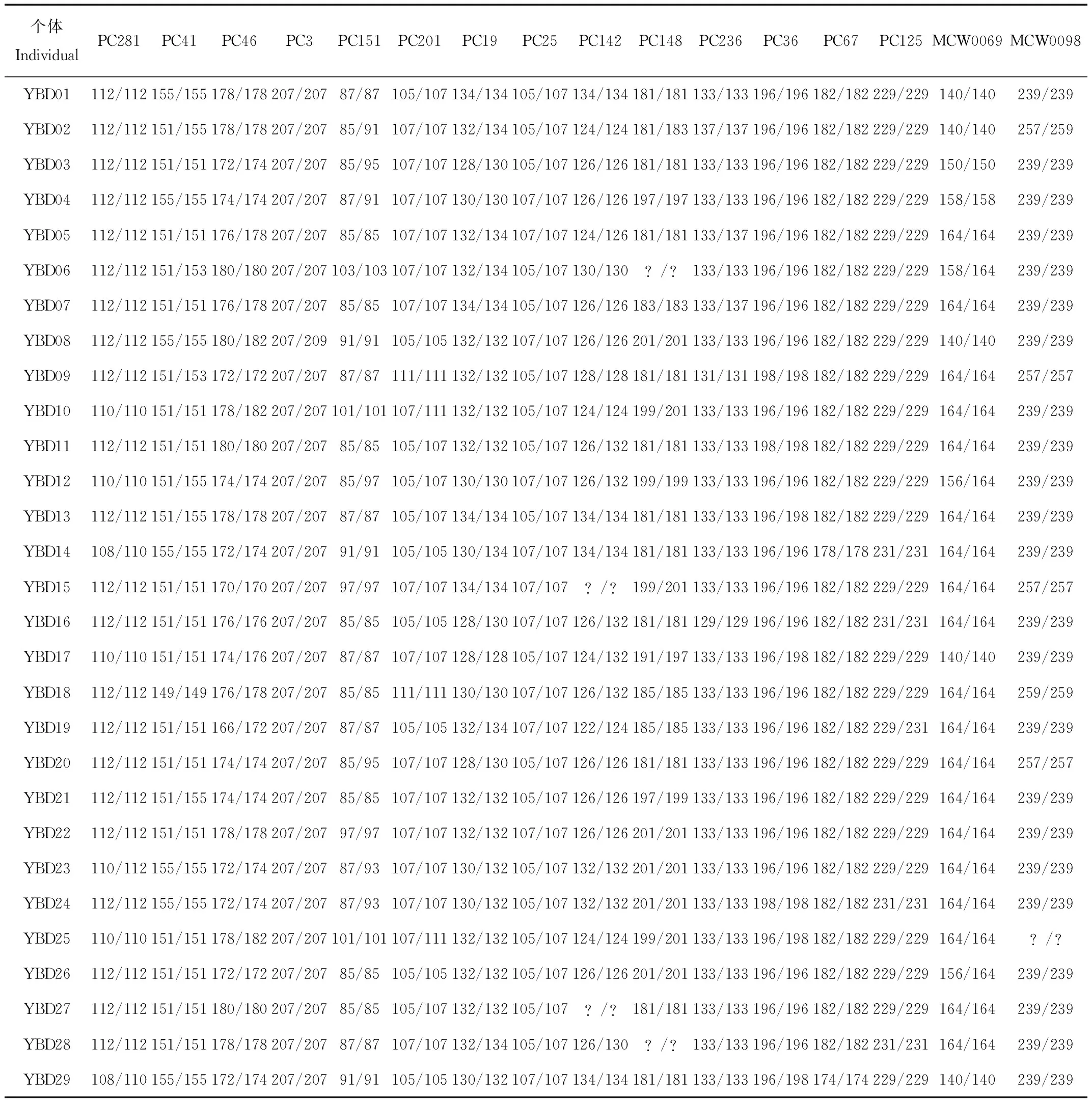
附錄1 云南玉溪綠孔雀種群的SSR分型結果
2 結果與分析
2.1 羽毛樣品的母系物種鑒定
作為參考種群的已知綠孔雀(GP)和藍孔雀(BP)的種間遺傳距離(inter-d)為3.43%~3.68%,95%的置信區間為[3.41%~3.61%];參考綠孔雀種群種內遺傳距離(intra-d)為0~0.72%,95%的置信區間為[0.26%~0.46%]。羽毛樣品和已知綠孔雀之間的K2P距離為0.120~0.602,均落入intra-d的95%的置信區間,而與藍孔雀的K2P距離在3.531%~3.788%,落入inter-d的95%的置信區間,且在intra-d的95%的置信區間之外(表1)。系統發育樹顯示29個羽毛樣品都與綠孔雀聚為一支(圖2),Bootstrap支持度為0.493~1.000。結果表明,所有的羽毛樣品的母系均為綠孔雀。

表1 以421 bp的Cyt b基因序列估算的羽毛樣品YBD01~29與純種綠孔雀(GP)和藍孔雀(BP)之間的K2P遺傳距離
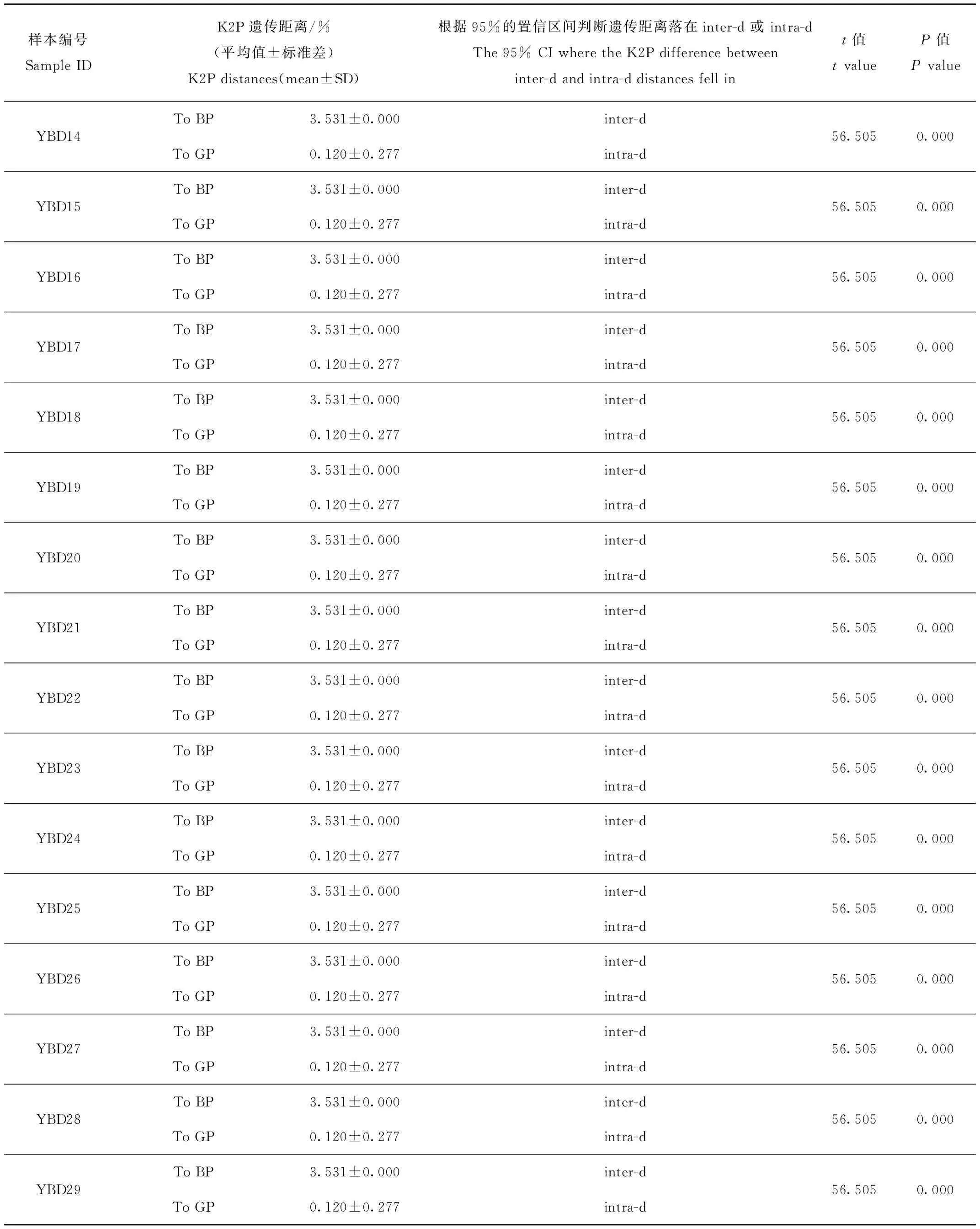
續表1
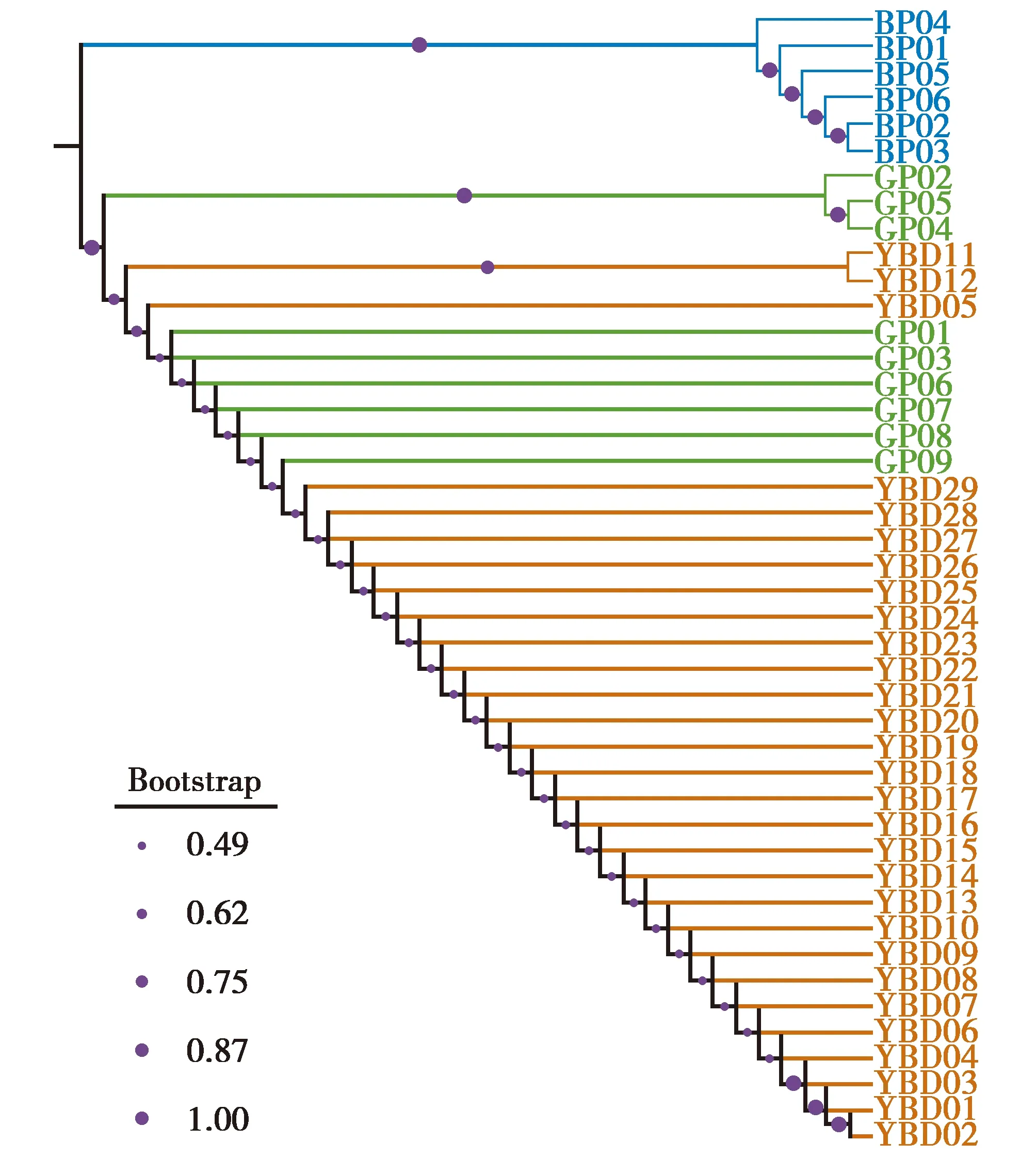
圖2 基于Cyt b基因421 bp序列的K2P遺傳距離所做的NJ系統發育樹顯示羽毛樣品(橙色)、純種綠孔雀(綠色)和純種藍孔雀(藍色)的關系Fig.2 The NJ tree of feather samples (orange) and reference samples of the green (green) and blue peafowl (blue) based on K2P genetic distance inferred from 421 bp sequence of Cyt b gene
2.2 羽毛樣本的個體識別
Micro-Checker在PC36、PC67、PC125、MCW0069和MCW0098這5個位點檢測到無效等位基因。為了保證個體識別結果的可靠性,后續分析中把這5個位點舍去,保留未檢測到無效等位基因的其余11個位點,即PC46、PC148、PC151、PC142、PC19、PC236、PC41、PC25、PC281、PC201和PC3。圖3顯示,累積P(ID)值隨著位點數的增加而下降。分子生態學研究中常以P(ID)值在0.001 0~0.000 1為同一性認定標準,即累積P(ID)達到0.001 0或0.000 1以下時,如果這2個樣品具有不同的基因型,則可判定為來自不同的個體[30]。本研究數據顯示,當微衛星位點數達到8個時,P(ID)sibs的聯合P(ID)達到0.000 963,而對于相同的8個位點,P(ID)biased的聯合P(ID)達到0.000 000 036 49。當位點更多時,累積P(ID)會進一步下降,增加基因型錯配的機會,影響個體識別結果的準確性[30](圖3)。同時考慮到綠孔雀在野外的稀有性,數值小于0.001 0的聯合P(ID)足以可靠地識別個體。
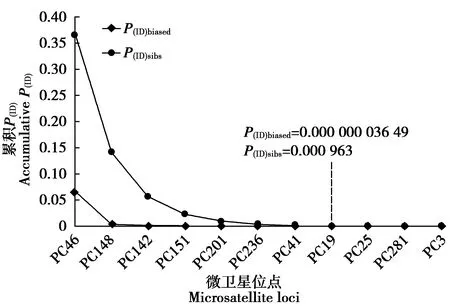
圖3 微衛星位點累積P(ID)的變化趨勢Fig.3 The declining trend of cumulative probability of identity (P(ID)) with addition of microsatellite loci
用8個微衛星位點通過Cervus軟件進行個體識別,結果顯示羽毛樣本中YBD10和YBD25、YBD11和YBD27、YBD23和YBD24在8個位點上基因型相同,或者僅在1個位點上只有1個等位基因不同,其余樣品顯示1個以上等位基因的錯配(附錄1)。最后,確定野外綠孔雀樣本為26只。
2.3 YBD和CAP種群的遺傳多樣性
利用Popgene檢測了YBD種群和CAP種群在11個微衛星位點的種群遺傳學參數。YBD種群的Na為2~8個,平均為(4.727±2.328)個;CAP種群的Na為2~6個,平均為(4.091±1.300)個;YBD種群和CAP種群之間等位基因頻率差異為0~0.846。2個種群之間Na在PC46上差異最大,YBD種群有8個等位基因,而CAP種群只有4個。總的來說,Na在YBD種群中比在CAP種群中更多(表2)。
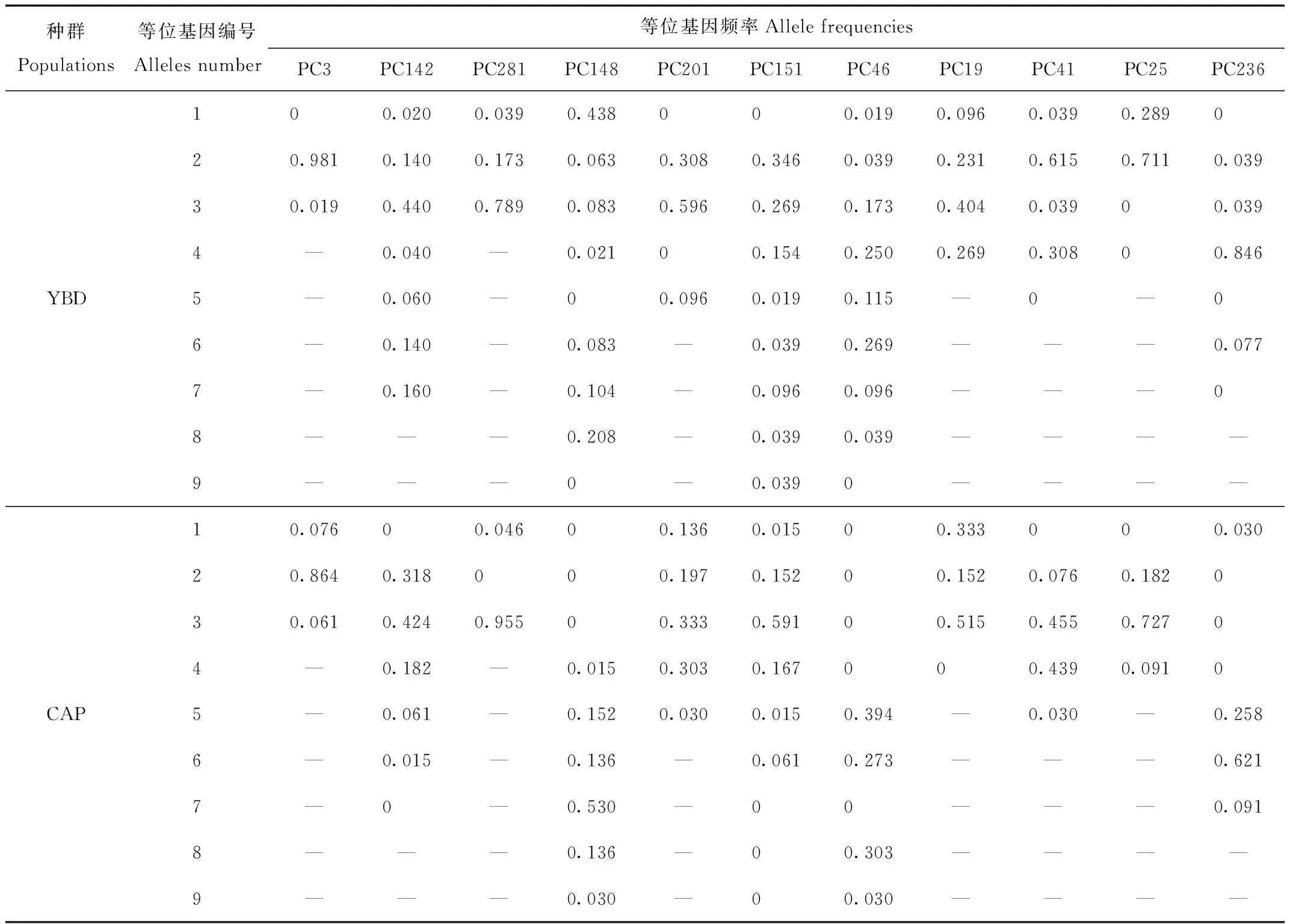
表2 11個微衛星位點在YBD和CAP種群中的等位基因和等位基因頻率
表3顯示,YBD種群中Ne的數量為1.039~5.240,平均為(2.768±1.294),而CAP種群Ne的數量為1.095~3.828,平均為(2.434±0.818)。在2個種群中,所有位點的Na都顯著大于Ne,表明2個種群都面臨遺傳多樣性喪失的風險。YBD種群的平均Ho和平均He分別為(0.258±0.164)和(0.546±0.251);CAP種群平均Ho和平均He分別為(0.479±0.228)和(0.531±0.201)。哈迪-溫伯格平衡(H-W)檢驗表明,YBD種群中有2個位點(PC3和PC25)處于平衡狀態(P>0.05),其余9個位點顯著偏離平衡狀態(P<0.05);相比之下,CAP種群中有7個位點處于平衡狀態(P>0.05),其余4個位點(PC201、PC236、PC148和PC151)表現出顯著偏離(P<0.05)。這11個位點在YBD和CAP種群中的平均PIC分別為(0.493±0.242)和(0.489±0.177)。YBD種群的FIS在0.720 (PC236)~-0.405 (PC25),平均值為(0.456±0.357),2個位點的FIS<0,9個位點的FIS>0,表明存在近親繁殖的趨勢;CAP種群的FIS在0.441 (PC151)~-0.199 (PC25),平均值為(0.102±0.217),5個位點的FIS<0,6個位點的FIS>0,表明近親繁殖趨勢不明顯(表4)。


續表3
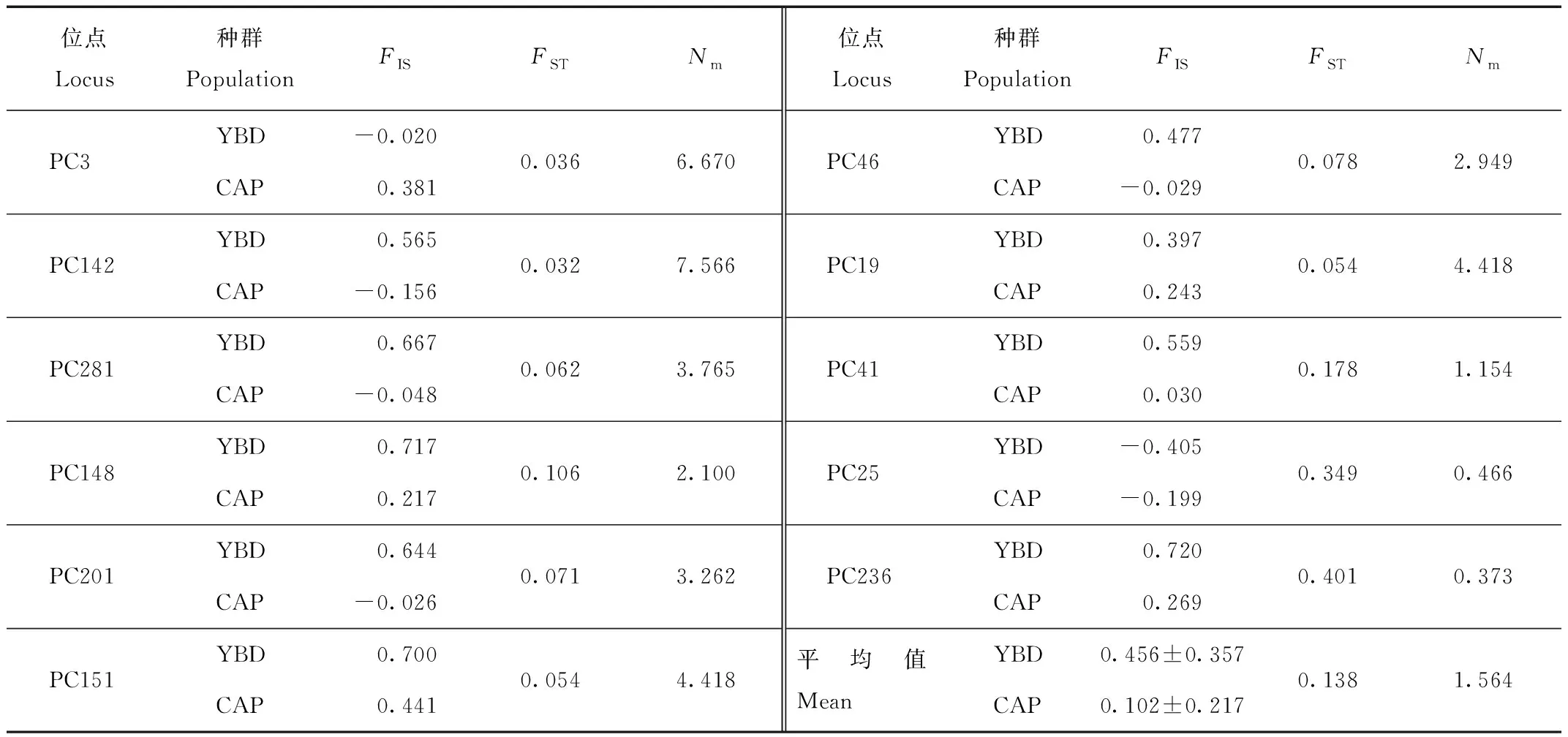
表4 YBD和CAP綠孔雀種群的11個微衛星位點的F檢驗結果
2.4 種群遺傳分化與遺傳結構分析
11個位點遺傳分化指數FST的平均值為0.138。根據Wright的分類標準[26],3個位點(PC41、PC25和PC236)遺傳分化程度較大(FST>0.150),6個位點(PC281、PC46、PC151、PC201、PC19和PC148)處于中度分化狀態(0.050 在進行種群遺傳結構分析時,預設K值(理論種群數)為2~4,每個K值重復運算10次,K=3時的DeltaK最大,表明本研究中所有參試個體可劃分為3個種群(圖4):CAP種群可劃分為2個亞群(紅色和綠色),而YBD種群為單一亞群(藍色),2個CAP亞群的遺傳成分占比很小。這說明CAP種群可能由來自不同地理區域的祖先發展而來,而2個亞群之間的基因交流十分有限。同時,CAP種群的祖先并非來自YBD種群。 圖4 YBD和CAP種群的STRUCTURE分析(K=3)Fig.4 STRUCTURE analysis of YBD and CAP populations (K=3) 云南省各地藍孔雀的養殖比較多,有研究懷疑養殖的藍孔雀發生逃逸,并在野外建立了種群[13,31],因此,首先需要鑒定羽毛是否來自綠孔雀。物種鑒定采用母系遺傳的線粒體DNA,如果母系來源于藍孔雀,排除在后續研究;母系屬于綠孔雀的樣品,仍需采用核DNA鑒定其核基因組是否來自綠孔雀。Cytb基因片段鑒定母系物種是野生動物法醫學鑒定的標準遺傳標記[18]。鑒定時以轉換(transition,Ts)為堿基替換模式計算K2P遺傳距離,以種間和種內的K2P距離范圍為參照[23],考察羽毛樣品與已知純種綠孔雀和藍孔雀之間的距離,落在種內還是種間的范圍內,95%的可信度確定其屬于綠孔雀還是藍孔雀;同時也用K2P遺傳距離構建NJ系統發育樹,通過Bootstrap再次考察其屬于哪個物種,本研究2種方法都一致支持所有羽毛樣本的母系來源均為綠孔雀(表1,圖2)。 核基因的鑒定中,高質量DNA正常分型的16個位點[18],有5個檢測到無效等位基因,說明從角化的羽翈獲得的DNA質量較低,對核基因的分型質量有明顯的影響。采用11個分析正常的位點計算累積P(ID),隨著位點的增加,累積P(ID)下降的速度(圖3),選定8個位點即可滿足個體識別可靠性的評估,又能減少分型誤差帶來的影響。最終以極高的可信度從29份樣品中鑒定了26只個體。 YBD種群的Na為2~8個,平均為4.727個,說明表觀上含有的遺傳變異較多,但是Ne平均值只有2.768,大多數位點(9/11)都顯著偏離H-W,這意味著強烈的遺傳漂變和非隨機交配將拉高遺傳多樣性丟失的風險。相比而言,CAP種群的Na為2~6個,平均為4.091個,Ne平均為2.434,均明顯地低于YBD種群,11個位點中有7個處于H-W狀態,表明遺傳多樣性丟失的風險小于YBD種群。 2個種群的雜合度都不高,CAP種群的Ho平均為0.479,而YBD種群的Ho只有0.258,說明遺傳多樣性很低;2個種群的He都高于各自的Ho,表明雜合度低于預期主要源自近交。種群近交系數(FIS)進一步印證這一點,YBD種群的近交程度(平均0.456)明顯高于CAP種群(平均0.102)。綠孔雀野外種群以小規模家族群為主[11,32],YBD種群很可能因為生境中的隔離因素導致族群間的基因交流困難,繁殖以局部近交為主。STRUCTURE分析顯示,YBD種群內部沒有發生顯著的遺傳分化(圖4)。種群歷史上的基因交流是比較頻繁而有效的,表現出來的近交并非是生境中景觀異質性形成長期的隔離,而是近期才出現的。 STRUCTURE分析表明(圖4),CAP種群包含2個遺傳譜系,一個是以秦皇島野生動物園飼養的綠孔雀為代表,最初建群者來自瑞麗地區;另一個譜系的建群者來源地沒有可靠的記錄。瑞麗地區、玉溪地區和CAP種群中的第2個譜系的綠孔雀之間有著極其明顯的遺傳分化,預示著整個云南種群的內部可能遺傳結構非常復雜,甚至有著深度的分化。 本研究顯示我國云南綠孔雀的地理分化可能非常顯著,需要開展系統的譜系地理學研究,厘清各地種群之間的遺傳關系,確認進化顯著單元。在開展遷地保護時,避免通過再引入打破進化顯著單元的獨特性。具體到玉溪地區的種群,內部沒有顯著的遺傳分化,遺傳多樣性水平較低,近期的近交可能導致遺傳多樣性的進一步丟失。管理上以加強族群之間的基因交流為目標,通過生態廊道的修建,或在不同族群之間進行個體轉移,增加遺傳交流的機會,扭轉遺傳多樣性隨近交的深化而不斷丟失的趨勢。 致謝:感謝秦皇島野生動物園和中國科學院西雙版納熱帶植物園提供研究樣品。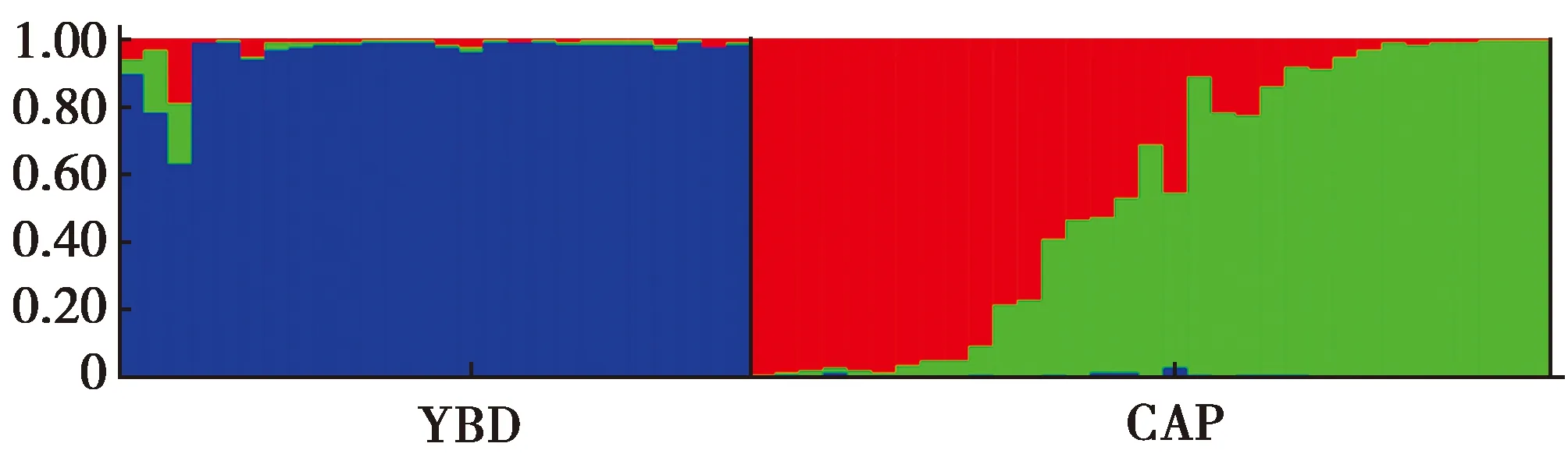
3 討論
3.1 羽毛樣品的物種鑒定和個體識別
3.2 YBD種群的遺傳多樣性
3.3 YBD種群與CAP種群的遺傳關系
4 結論與建議
