PI3K/Akt信號通路在HIV感染中的研究進(jìn)展
2022-11-07 09:51:22馬沖,陶莊,王健
傳染病信息 2022年5期
關(guān)鍵詞:信號
馬 沖,陶 莊,王 健
磷脂酰肌醇3-激酶(phosphoinositide 3-kinase,PI3K)/蛋白激酶B(protein kinase B, PKB,普遍寫作Akt)在調(diào)控細(xì)胞的代謝、生長、增殖、存活、轉(zhuǎn)錄以及蛋白質(zhì)合成等方面具有重要作用,其功能的失調(diào)與腫瘤、內(nèi)分泌疾病、心血管疾病、神經(jīng)系統(tǒng)疾病和AIDS等多種疾病的發(fā)生密切相關(guān)[1-2],這使其成為人類疾病治療的重要靶標(biāo)。近年研究發(fā)現(xiàn)PI3K/Akt信號通路在HIV感染中具有重要作用,可通過下游多種效應(yīng)分子對HIV感染、HIV復(fù)制、病毒儲存庫及T細(xì)胞發(fā)育的各個階段等方面發(fā)揮重要的調(diào)節(jié)作用,因此,探索該信號通路在HIV/AIDS中的作用機制具有重要意義。本研究將對PI3K/Akt信號通路與HIV/AIDS的相關(guān)研究進(jìn)行整合,旨在為AIDS的治療提供新的思路與靶點。
1 PI3K/Akt結(jié)構(gòu)與功能
1.1 PI3K PI3K是一個脂質(zhì)激酶家族,通過特異性磷酸化肌醇磷脂中的3-羥基而產(chǎn)生第二信使[3]。PI3K分為Ⅰ~Ⅲ3類,Ⅰ類組成部分中,p110催化亞基可影響細(xì)胞增殖,參與免疫功能和炎癥的調(diào)節(jié)[4],P85亞基在受體結(jié)合、酶激活等過程中發(fā)揮作用[5];Ⅱ類PI3K在葡萄糖轉(zhuǎn)運、細(xì)胞遷移、細(xì)胞生長及細(xì)胞存活等方面具有重要作用[6];Ⅲ類PI3K是一種重要的自噬和囊泡運輸調(diào)節(jié)因子,在人體組織中廣泛表達(dá),對細(xì)胞增殖至關(guān)重要[7]。
1.2 Akt Akt是一種絲氨酸/蘇氨酸激酶,是PI3K/Akt/哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin, mTOR)信號通路的中心介質(zhì),能夠調(diào)節(jié)細(xì)胞生長,還可以實現(xiàn)對細(xì)胞存活的調(diào)節(jié)作用,其發(fā)揮作用的途徑主要是通過直接抑制促凋亡蛋白或抑制轉(zhuǎn)錄因子產(chǎn)生的凋亡信號,這對于調(diào)控細(xì)胞的凋亡、增殖、遷移、轉(zhuǎn)錄以及糖代謝起到十分重要的作用,是B細(xì)胞及T細(xì)胞發(fā)育、激活和分化的關(guān)鍵分子[8]。同時,Akt能夠抑制線粒體自噬,并促進(jìn)細(xì)胞在線粒體應(yīng)激條件下存活[9]。
1.3 mTOR mTOR是PI3K相關(guān)激酶家族的絲氨酸/蘇氨酸激酶,是一種調(diào)節(jié)細(xì)胞生長、存活、代謝和免疫的蛋白激酶,參與了B細(xì)胞與T細(xì)胞的活化與分化[10]。mTOR由mTOR復(fù)合物(mTOR complex, mTORC)1和mTORC2組成,mTORC1主要調(diào)節(jié)細(xì)胞生長和能量代謝,mTORC2主要控制細(xì)胞骨架重塑、細(xì)胞增殖和存活[11]。
2 PI3K/Akt信號通路在HIV感染中具有重要作用
2.1 PI3K/Akt激活能夠促進(jìn)HIV感染 HIV進(jìn)入靶細(xì)胞依賴于主要受體CD4與協(xié)同受體趨化性細(xì)胞因子受體5(chemotactic cytokine receptor 5, CCR5)/CXC趨化因子受體4(CXC chemokine receptor-4, CXCR4)的表達(dá),HIV包膜糖蛋白與靶細(xì)胞表面CD4的相互作用能夠使CD4磷酸化,通過與非共價相互作用的酪氨酸激酶解離而使受體內(nèi)吞,HIV包膜糖蛋白120(glycoprotein 120, gp120)與CCR5/CXCR4的相互作用導(dǎo)致病毒與靶細(xì)胞膜融合,從而建立了病毒入口,介導(dǎo)病毒進(jìn)入靶細(xì)胞。在此過程中,HIV gp120與CXCR4的相互作用能夠誘導(dǎo)PI3K/Akt信號通路激活,通過LIM和SSH2條拮抗途徑的激活而促進(jìn)絲切蛋白活性增強,有助于增強T細(xì)胞對HIV的易感性,從而促進(jìn)HIV進(jìn)入、DNA合成和細(xì)胞內(nèi)遷移等過程[12-13]。同時,PI3K亞基p110α在HIV進(jìn)入靶細(xì)胞過程中發(fā)揮重要作用,該亞基可調(diào)控HIV包膜糖蛋白介導(dǎo)的細(xì)胞融合;相反,人類第10號染色體缺失的磷酸酶和張力蛋白同源物(phosphatase and tensin homolog deleted on chromosome ten, PTEN)作為PI3K/Akt信號通路的下游蛋白,對HIV進(jìn)入靶細(xì)胞過程具有負(fù)調(diào)控作用,該蛋白可使PI3K產(chǎn)物去磷酸化并降低Akt活性,從而抑制HIV通過Akt依賴性途徑進(jìn)入靶細(xì)胞,并抑制細(xì)胞生長及其他PI3K/Akt依賴性生物過程,但PI3K/Akt激活后可下調(diào)PTEN的表達(dá),從而有利于HIV對靶細(xì)胞的感染[14]。見圖1。
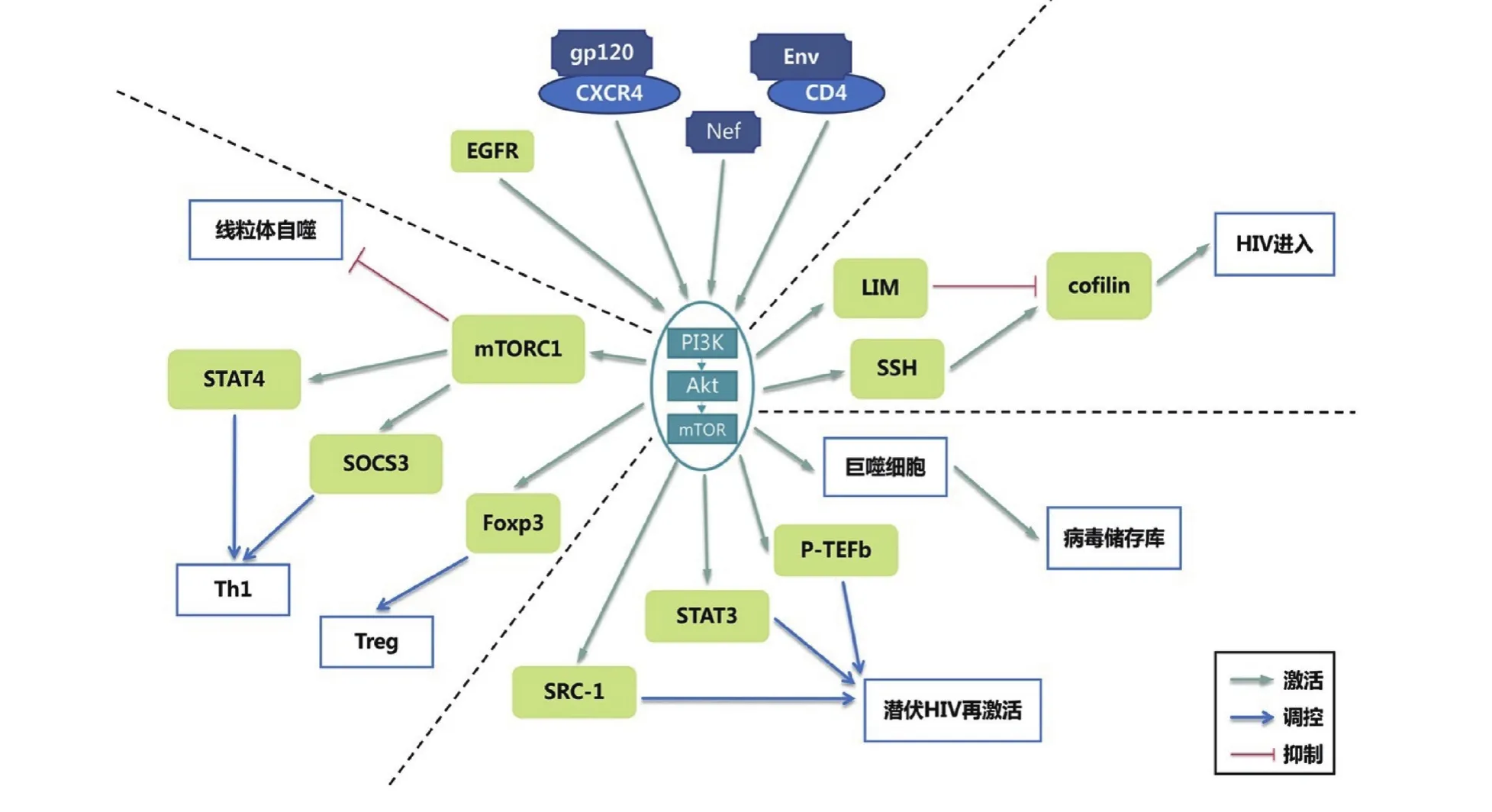
圖1 PI3K/Akt信號通路在HIV感染中的作用機制Nef. 負(fù)調(diào)控因子;Treg. 調(diào)節(jié)性T細(xì)胞;Env.包膜; STAT4. 信號傳導(dǎo)轉(zhuǎn)錄激活因子4;mTORC1. 哺乳動物雷帕霉素靶蛋白復(fù)合物1;SOCS3.細(xì)胞因子信號轉(zhuǎn)導(dǎo)抑制因子3;Foxp3.叉頭盒蛋白p3基因;Th1.輔助性T細(xì)胞1;SRC-1. 類固醇受體共激活劑-1;STAT3. 信號傳導(dǎo)轉(zhuǎn)錄激活因子3;p-TEFb. 正轉(zhuǎn)錄延伸因子bFigure 1 The mechanism of PI3K / Akt signaling pathway in HIV infection
2.2 PI3K/Akt參與維持病毒儲存庫并促使?jié)摲腍IV再激活 抗反轉(zhuǎn)錄病毒治療(antiretro viral treatment, ART)無法清除記憶CD4+T細(xì)胞中的病毒儲存庫,宿主淋巴結(jié)、脾臟以及腸道等組織中的巨噬細(xì)胞也可作為機體的病毒儲存庫并持續(xù)復(fù)制HIV,而病毒儲存庫的持續(xù)存在也是HIV感染者停止ART后疾病進(jìn)展的重要因素[15-16]。研究發(fā)現(xiàn),巨噬細(xì)胞中的HIV能夠誘發(fā)Akt膜定位并激活A(yù)kt,活化的Akt通過下調(diào)促炎反應(yīng)并誘導(dǎo)抗炎反應(yīng)的轉(zhuǎn)變而激活巨噬細(xì)胞,使感染后的巨噬細(xì)胞壽命延長,提高受感染細(xì)胞的存活率,使其成為病毒儲存庫的重要組成部分,并能維持病毒儲存庫的存活以及HIV的持續(xù)復(fù)制[17-18],這說明PI3K/Akt信號通路可能在維持病毒儲存庫中具有重要作用。見圖1。
HIV從潛伏期中的再激活與正轉(zhuǎn)錄延伸因子b(positive transcription elongation factor b, P-TEFb)密切相關(guān),而PI3K/Akt信號通路參與P-TEFb表達(dá)的調(diào)控[19],該通路的激活可能通過促進(jìn)P-TEFb亞基的翻譯以促進(jìn)P-TEFb的生成。研究發(fā)現(xiàn),PI3Kp110α特異性小分子激動劑(57704)能夠通過激活PI3K/Akt信號通路而重新激活潛伏的HIV[20]。PTEN作為Akt的負(fù)調(diào)控因子,可通過去磷酸化將PIP3轉(zhuǎn)化為PIP2,減少Akt活化,從而抑制多個細(xì)胞過程,而抑制PTEN后Akt磷酸化增加,可激活潛伏的HIV[21]。樹突狀細(xì)胞(dendritic cell, DC)與CD4+T細(xì)胞相互作用后能夠使?jié)摲腍IV得到激活,接受T細(xì)胞抗原受體(T cell receptor, TCR)與DC雙重刺激能夠更頻繁地逆轉(zhuǎn)潛伏期,研究表明這一過程與PI3K/Akt信號通路相關(guān)[22]。此外,Barclay等[23]的研究顯示,未被HIV感染細(xì)胞的外泡囊(extracellular vesicles, EVs)含有大量非受體酪氨酸激酶(cell-sarcoma receptor coactivator, c-Src),c-Src能夠促使表皮生長因子受體(epidermal growth factor receptor, EGFR)激活并激活PI3K/Akt信號通路,從而激活信號傳導(dǎo)轉(zhuǎn)錄激活因子(signal transducer and activator of Transcription, STAT)3與可溶性補體受體1型(soluble complement receptor 1,SCR-1),最終逆轉(zhuǎn)HIV的潛伏,抑制c-Src后可導(dǎo)致EGFR、PI3K、Akt-1、STAT3、SCR-1等的抑制,并降低細(xì)胞內(nèi)HIV水平,證實上述蛋白在潛伏的HIV再激活中的重要性。以上研究表明,PI3K/Akt信號通路在促進(jìn)潛伏的HIV再激活方面具有重要意義,因此,也有研究者基于上述理論提出應(yīng)用潛伏期逆轉(zhuǎn)劑激活潛伏的HIV,使免疫系統(tǒng)與ART能夠清除病毒與受感染的細(xì)胞,為“休克和殺死”療法的應(yīng)用提供了一定的基礎(chǔ),目前Akt激活劑已作為潛伏期逆轉(zhuǎn)劑進(jìn)行了研究[24]。見圖1。
2.3 PI3K/Akt信號通路對T細(xì)胞的調(diào)節(jié) TCR與CD28可導(dǎo)致PI3K/Akt信號通路激活,其下游蛋白mTOR的磷酸化可促進(jìn)Akt最大限度激活,磷酸化的Akt能夠?qū)細(xì)胞多個階段的功能發(fā)揮作用[25]。PI3K/Akt信號通路對調(diào)節(jié)性T細(xì)胞(regularory T cell, Treg)的發(fā)育具有積極作用,TCR信號轉(zhuǎn)導(dǎo)在促使PI3K/Akt激活后,能夠?qū)е虏骖^盒蛋白p3基因(forkhead box p3, Foxp3)的過度表達(dá),從而導(dǎo)致Treg細(xì)胞活化增加[26]。但是該過程在HIV感染中具有一定爭議,一方面,Treg直接降低了活化的T細(xì)胞數(shù)量,減少了HIV在靶細(xì)胞中的復(fù)制;但另一方面,Treg激活可抑制CD4+T細(xì)胞的特異性免疫應(yīng)答,有助于通過調(diào)節(jié)抗HIV的免疫反應(yīng)來建立高水平的HIV復(fù)制,降低機體免疫應(yīng)答能力,且Treg表面相關(guān)受體(CXCR4)還可增加其對HIV的易感性[27]。相反,也有研究顯示PI3K/Akt信號通路對于Treg具有負(fù)調(diào)節(jié)作用,缺少PI3K p110δ的小鼠胸腺中的Treg數(shù)量增加[28],mTOR負(fù)向調(diào)節(jié)Treg的發(fā)育與分化[29]。此外,CD4+T細(xì)胞是HIV攻擊的靶細(xì)胞,導(dǎo)致輔助性T細(xì)胞(helper T cell, Th)破壞,降低細(xì)胞免疫功能[30],mTORC1通過STAT4和SOCS3正向調(diào)節(jié)Th1的分化,但mTORC2對Th1的分化作用還有待進(jìn)一步驗證。見圖1。
2.4 PI3K/Akt改變線粒體功能 線粒體是人類細(xì)胞中參與能量產(chǎn)生的主要細(xì)胞器,HIV感染以及ART可引起線粒體DNA(mitochondrial DNA, mtDNA)損傷以及線粒體膜電位改變,導(dǎo)致活性氧(reactive oxygen species, ROS)產(chǎn)生增加,膜電位下降,干擾機體氧化磷酸化過程中相關(guān)蛋白的產(chǎn)生,使線粒體功能損傷而促使CD4+T細(xì)胞死亡[31],線粒體自噬能夠特異性清除受損的線粒體,維持線粒體的功能以及細(xì)胞的穩(wěn)態(tài)[32]。PI3K/Akt信號通路在調(diào)節(jié)自噬中具有不可或缺的作用,mTOR是調(diào)節(jié)自噬的關(guān)鍵信號分子[33]。最近的研究認(rèn)為,mTORC1是線粒體自噬的負(fù)調(diào)節(jié)因子,雷帕霉素(mTOR抑制劑)能夠增強線粒體自噬,并抑制細(xì)胞凋亡[34]。其他研究顯示,磷酸化的Akt通過激活下游mTOR而調(diào)節(jié)線粒體介導(dǎo)的細(xì)胞凋亡,抑制Akt磷酸化后,可見線粒體膜電位增加,細(xì)胞內(nèi)ROS水平下降[35]。因此,PI3K/Akt信號通路可能參與HIV感染導(dǎo)致線粒體功能受損的過程,并參與線粒體自噬的調(diào)控,抑制PI3K/Akt信號通路可能促使線粒體功能恢復(fù),延緩AIDS進(jìn)展。見圖1。
3 PI3K/Akt作為HIV感染治療靶點
由于PI3K/Akt信號通路在誘導(dǎo)HIV進(jìn)入靶細(xì)胞、維持病毒儲存庫、促進(jìn)潛伏HIV再激活、改變線粒體功能等方面具有重要作用,因此,靶向PI3K/Akt治療可能會成為治療HIV的重要療法。
3.1 抑制HIV進(jìn)入靶細(xì)胞 在HIV感染的治療中,抑制HIV進(jìn)入靶細(xì)胞的研究是HIV感染領(lǐng)域的重要研究方向,目前正在研發(fā)的抑制劑主要包括HIV吸附抑制劑、HIV受體拮抗劑及HIV與靶細(xì)胞融合抑制劑等。Hamada等[14]的研究表明,gp120與CD4的相互作用可導(dǎo)致PI3K亞基快速激活,PI3K p110α特異性抑制劑(PIK-75)能夠抑制HIV進(jìn)入T細(xì)胞,降低HIV誘導(dǎo)的Akt磷酸化;并且PIK-75能夠抑制病毒與細(xì)胞的融合,使得HIV 包膜與靶細(xì)胞表面CD4的相互作用受到阻斷,從而阻止HIV包膜介導(dǎo)的細(xì)胞融合過程,這提示PI3K抑制劑可用于抑制HIV進(jìn)入靶細(xì)胞的過程。另有研究表明,PI3K抑制劑不僅能夠阻斷HIV對CD4+T細(xì)胞的感染,還可對已激活的CD4+T細(xì)胞生命周期發(fā)揮一定的抑制作用,抑制p85亞基(PI3K亞基)能夠阻斷Toll樣受體(Toll-like receptor, TLR)2對HIV感染的增強作用[36]。
3.2 抑制HIV在細(xì)胞間的復(fù)制及遷移 HIV負(fù)調(diào)控因子(negativefactor, Nef)是一種多功能病毒輔助蛋白,不具有酶活性,在HIV感染早期大量表達(dá),在增強病毒感染與復(fù)制方面具有重要作用,而Nef的缺失可有效減緩AIDS的發(fā)展[37]。HIV Nef通過影響TCR的信號轉(zhuǎn)導(dǎo)而防止受感染細(xì)胞過早激活誘導(dǎo)的死亡,并促進(jìn)HIV在靶細(xì)胞中的復(fù)制[38]。HIV趨化因子能夠?qū)⑹芨腥炯?xì)胞由黏膜運輸至淋巴結(jié)而感染T細(xì)胞,隨后受感染的T細(xì)胞在淋巴結(jié)內(nèi)的活動可刺激B細(xì)胞以建立體液免疫反應(yīng),HIV Nef能夠通過干擾趨化因子而防止受感染的T細(xì)胞遷移至淋巴結(jié),從而破壞機體的免疫反應(yīng)[39]。HIV Nef分別通過N末端和C末端與PI3K和Akt相互作用,從而激活PI3K/Akt信號通路,提高受感染細(xì)胞的存活率,蛋白酶抑制劑(protease inhibitors, PI)能夠以時間和劑量依賴性方式抑制HIV Nef誘導(dǎo)的Akt過度活化和T細(xì)胞增殖,并可阻斷病毒在T細(xì)胞間的復(fù)制[40-41]。因此,基于HIV Nef激活PI3K/Akt過程探索阻斷HIV感染細(xì)胞的遷移及抑制HIV在T細(xì)胞中的復(fù)制,對于優(yōu)化HIV治療方案具有重要意義,可能成為潛在的治療策略。
3.3 抑制病毒儲存庫 HIV對PI3K/Akt信號通路的激活能夠?qū)λ拗骷?xì)胞壽命的延長發(fā)揮重要作用,因此,以PI3K/Akt為靶點的抑制劑可能對病毒儲存庫具有抑制作用。有研究顯示,5, 7-二羥基-6-甲氧基黃酮可通過抑制PI3K/Akt信號通路,而減弱其對病毒儲存庫的保護(hù)作用,并且能夠促進(jìn)已感染的巨噬細(xì)胞凋亡而發(fā)揮抑制病毒儲存庫的作用[42]。
3.4 免疫調(diào)節(jié) HIV感染導(dǎo)致CD4+T細(xì)胞功能缺陷及凋亡,細(xì)胞凋亡誘導(dǎo)的淋巴細(xì)胞持續(xù)減少促使機體免疫失衡。紅色諾卡氏菌細(xì)胞壁骨架(nocardia rubra cell-wall skeleton, Nr-CWS)作為免疫增強劑,可促進(jìn)巨噬細(xì)胞、DC細(xì)胞、自然殺傷(natural killer, NK)細(xì)胞、CD4+T及CD8+T細(xì)胞活性的增強[43]。相關(guān)試驗表明,Akt磷酸化可由Nr-CWS誘導(dǎo),Nr-CWS激活PI3K/Akt信號通路后能夠促進(jìn)PAX8的表達(dá),上調(diào)的PAX8可進(jìn)一步促進(jìn)Th1細(xì)胞的分化與成熟[44]。同時,I型干擾素(Type I interferons,IFN-I)是在急性病毒感染中由病毒刺激漿細(xì)胞樣樹突狀細(xì)胞(plasmacytoid dendritic cells, pDC)產(chǎn)生,可導(dǎo)致HIV感染者的記憶CD4+T細(xì)胞凋亡,并直接參與免疫損傷[45]。泛素特異性肽酶(ubiquitin specific peptidase 18, USP18)是IFN-1的負(fù)調(diào)節(jié)蛋白,抑制USP18的表達(dá)可促使機體以Akt依賴性的方式抑制記憶CD4+T細(xì)胞凋亡[46]。但通過激活PI3K/Akt信號通路調(diào)節(jié)免疫可能與該通路介導(dǎo)的HIV進(jìn)入、病毒儲存庫等過程具有一定矛盾性,因此該類治療策略還需綜合多種因素并進(jìn)一步深入研究。
4 小結(jié)與展望
自HIV被發(fā)現(xiàn)以來,人類在預(yù)防HIV傳播和HIV/AIDS的管理方面取得了較大進(jìn)步,ART也使其發(fā)展為慢性可控性疾病,但是病毒儲存庫、免疫重建不良以及耐藥等問題成為目前HIV/AIDS治療的難題。PI3K/Akt信號通路在HIV/AIDS多方面發(fā)揮作用,不僅在HIV與靶細(xì)胞融合過程中發(fā)揮重要作用,還參與維持病毒儲存庫與HIV的潛伏。同時,PI3K/Akt信號通路在T細(xì)胞發(fā)育的各個階段均可發(fā)揮作用,其作用根據(jù)T細(xì)胞的成熟階段、譜系及環(huán)境的不同而發(fā)生改變,而以PI3K/Akt為靶點的研究也顯示,相關(guān)抑制劑能夠在抑制HIV進(jìn)入靶細(xì)胞、清除病毒儲存庫等方面發(fā)揮作用。因此,基于PI3K/Akt信號通路探索抗HIV藥物研發(fā)、清除病毒儲存庫及改善HIV/AIDS患者免疫功能等方面可能具有較大發(fā)展空間。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
