呼倫貝爾黃花苜蓿MfMYB30基因的克隆及表達分析
2022-11-09 12:15:00周昕越劉葉飛韓慧杰劉亞玲
西北植物學報 2022年9期
周昕越,劉葉飛,邱 銳,韓慧杰,劉亞玲,趙 彥*
(1 內蒙古農業大學 草原與資源環境學院/農業部飼草栽培、加工與高效利用重點實驗室/教育部草地資源重點實驗室/教育部創新團隊——草地資源可持續利用的研究[IRT-17R59],呼和浩特 010018;2 內蒙古蒙草生態環境[集團]股份有限公司,呼和浩特 010030)
黃花苜蓿(MedicagofalcataL.)是多年生苜蓿屬豆科牧草,適口性好[1],營養價值非常高,具有很強的抗逆性[2]。在解決中國畜牧業面臨的蛋白質飼料缺乏、改善天然草地和草原土壤肥力下降等問題中具有重要的地位和作用[3]。由于現有黃花苜蓿品種的再生性和抗病性較差,容易受到生物和非生物因素的脅迫,從而影響品質和產量[4-6]。為了降低這些影響,植物進化出一系列復雜的抵御外界脅迫的反應機制,即通過轉錄因子整合外界信號和基因調控的方式產生相應的蛋白和代謝產物來適應脅迫,從而提高植物的抗逆性[7]。
植物中的MYB家族十分龐大,尤其是R2R3-MYB亞家族[8-9]。MYB類蛋白的N端通常含有約51個氨基酸組成的高度保守的MYB結構域,C端為轉錄激活結構域。MYB30轉錄因子屬于R2R3-MYB家族,R2R3-MYB轉錄因子擁有一個N端保守的MYBDNA結合結構域和一個通常定位在C端的可變的激活或抑制區域[10]。MYB30轉錄因子最早被發現是在植物抵抗病原菌侵染的HR(Hypersensitive Response)反應的最初階段[11]。研究顯示,MYB30轉錄因子參與了多種植物的生長發育和脅迫響應過程,如參與植物次生生長、激素信號轉導以及參與多種植物逆境防御反應[12],在信號通路中,MYB30轉錄因子通過與其他蛋白互作或是在蛋白水平上受到修飾,或是調節下游基因從而參與脅迫應答[13]。還有研究表明,MYB30轉錄因子具有生物學功能的多效性,可在非生物逆境抗性及抗病性中具有生物學功能[14]。對模式植物擬南芥的研究表明,MYB30轉錄因子可以加快擬南芥的開花過程,從而將其病原菌脅迫相應通路與開花調控過程聯系在一起[15]。桑樹MaMYB30基因可能參與桑樹對桑脈帶相關病毒的防御調控過程,證明該基因與植物抵御逆境脅迫關系密切[16]。MYB30還可以通過調控超長鏈脂肪酸(Very Long Chain Fatty Acid, VLCFA)的代謝參與低氧脅迫響應[17]。由此可見,MYB30轉錄因子基因在介導植物抵御生物與非生物脅迫中發揮著重要作用。
綜上,本研究以呼倫貝爾黃花苜蓿(MedicagofalcataL.cv. Hulunbuir)為材料,利用RT-PCR技術克隆得到黃花苜蓿MfMYB30基因,對該基因進行生物信息學特征分析,利用熒光定量PCR技術分析MfMYB30在不同刈割天數后的相對表達,并在擬南芥原生質體中進行了亞細胞定位分析,以期為黃花苜蓿抗逆與再生基因篩選及分子育種應用提供依據。
1 材料和方法
1.1 試驗材料與處理
本研究選取的植物材料為呼倫貝爾黃花苜蓿,挑選顆粒飽滿的黃花苜蓿種子,用砂紙打磨破除硬實,放在培養皿里培養至發芽,將發芽的種子播種于無菌培養土中并放至人工氣候室中培養,栽培期間為長日照條件(16 h光照/8 h黑暗),晝/夜溫度為26 ℃/18 ℃,通風良好,定期澆水。當生長到開花期時,進行刈割脅迫處理。
選取3株長勢較好并處于開花期的黃花苜蓿進行模擬刈割處理,刈割留茬高度15 cm,分別處理3、5和7 d,各時間點選取葉片與莖組織于2 mL離心管,并選取一株未進行刈割處理的植株作為對照,同時取樣,液氮速凍,-80 ℃條件保存備用,生物學重復3次。
1.2 方 法
1.2.1 RNA的提取與cDNA的合成取-80 ℃保存備用的黃花苜蓿樣品按照TransZoL. Up Plus RNA kit試劑盒(北京全式金基因科技有限公司)參考說明書提取總RNA。利用紫外分光光度計測定RNA含量,經1.2%瓊脂糖凝膠電泳檢驗,將質量合格的RNA在-80 ℃條件保存備用。以測定后的高質量總RNA為模板,使用TransScriptⅡOne-Step gDNA Removal and cDNA Synthesis SuperMix(北京全式金基因科技有限公司)試劑盒進行cDNA第一鏈的合成,-20 ℃條件保存備用。
1.2.2MfMYB30基因的克隆根據轉錄組測序數據篩選的差異表達候選基因,獲取MfMYB30基因全長序列,用Oligo7.0軟件設計擴增引物(表1),以反轉錄cDNA第一鏈為模板,進行PCR擴增。PCR擴增體系為50 μL,包含cDNA模板2 μL,高保真酶PrimeSTAR 25 μL,上、下游引物各2.5 μL,ddH2O補至50 μL。擴增程序為94 ℃預變性5 min,94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸90 s,35個循環,72 ℃延伸5 min,16 ℃保存。將擴增產物進行瓊脂糖凝膠電泳檢測合格后膠回收,然后將其連接到克隆載體1300-cYFP上,采用熱激法轉化大腸桿菌感受態DH5α,挑取陽性單菌落,經過菌落以及菌液PCR檢測后送往廣州睿博生物技術有限公司進行測序。

表1 所用引物信息
1.2.3 生物信息學分析對克隆得到的MfMYB30基因序列進行生物信息學分析,運用NCBI中的BLAST (https://bL.ast.ncbi.nL.m.nih.gov/BL.ast.cgi)功能進行序列比對;利用NCBI中的Open Reading Frame Finder(https://www.ncbi.nL.m.nih.gov/orffinder/)工具,找到cDNA序列中的最大開放閱讀框;利用SMART(http://smart.embL.-heideL.berg.de/)在線工具分析保守結構域;利用ExpasyProtParam tooL.(https://web.expasy.org/protparam/)工具分析蛋白質理化性質;利用PRABI(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_ automat.pL.?page=npsa_sopma.htmL.)和SWISS-MODEL.(https://swissmodeL..expasy.org/interactive)在線軟件分別預測蛋白二級結構。利用SignaL.P-5.0(https://services.heaL.thtech.dtu.dk/service.php?SignaL.P-5.0)預測蛋白質信號肽;TMHMM-2.0(https://services.heaL.thtech.dtu.dk/service.php?TMHMM-2.0)分析蛋白跨膜結構;利用Plant CARE在線軟件(https://bioinformatics.psb.ugent.be/webtooL.s/pL.antcare/htmL./)分析預測順式作用元件。利用MEGA7.0軟件、ITOL在線軟件及鄰接法(Neighbor-Joining, NJ)(bootstrap=1 000)構建系統進化樹,分析其與其他物種之間的進化關系。
1.2.4MfMYB30基因的表達分析根據克隆得到的MfMYB30基因序列,采用實時熒光定量PCR方法,設計qRT-PCR引物(表1),以黃花苜蓿Actin基因作為內參基因,檢測黃花苜蓿分別在模擬刈割脅迫處理3、5和7 d后的表達情況。試驗進行3次生物學重復。反應結束后對結果進行分析。使用Excel按照2-ΔΔCT方法進行計算,樣品間的差異性用SPSS 25.0進行檢驗。
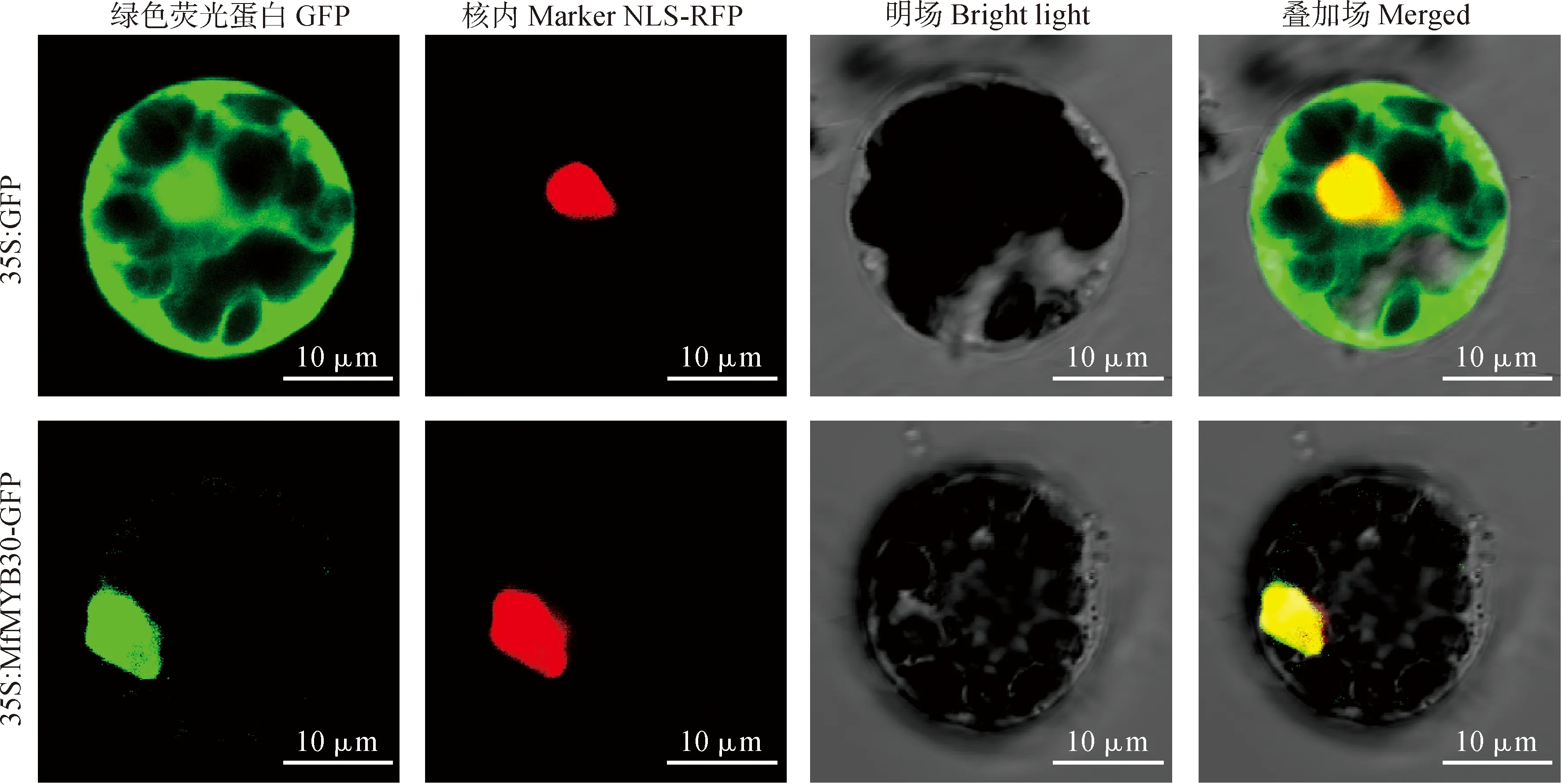
1.2.5 MfMYB30亞細胞位置的預測及定位利用在線軟件WoLFPSORTPrediction對MfMYB30蛋白進行亞細胞定位預測分析。采用酶切連接法構建以PUC19為骨架的pUC19-HA-MfMYB30-GFP瞬時表達載體。以空載體GFP作為對照,將構建好的載體pUC19-HA-MfMYB30-GFP及核定位信號(nucle-us localization signal,NLS)RFP融合紅色熒光蛋白的載體共同轉化擬南芥原生質體,利用激光共聚焦顯微鏡觀察熒光信號,進行MfMYB30亞細胞定位。
2 結果與分析
2.1 MfMYB30基因的克隆
以呼倫貝爾黃花苜蓿總RNA反轉錄的cDNA為模板進行PCR 擴增,得到大小為1 000 bp左右的片段,所得片段大小與預期理論值一致。對片段進行回收純化、載體連接及轉化,最后將陽性菌液送至廣州睿博生物技術有限公司進行測序,測序結果顯示片段長1 060 bp(圖1)。進一步分析表明該序列開放閱讀框(ORF)957 bp,編碼318個氨基酸(圖2),并將其命名為MfMYB30。
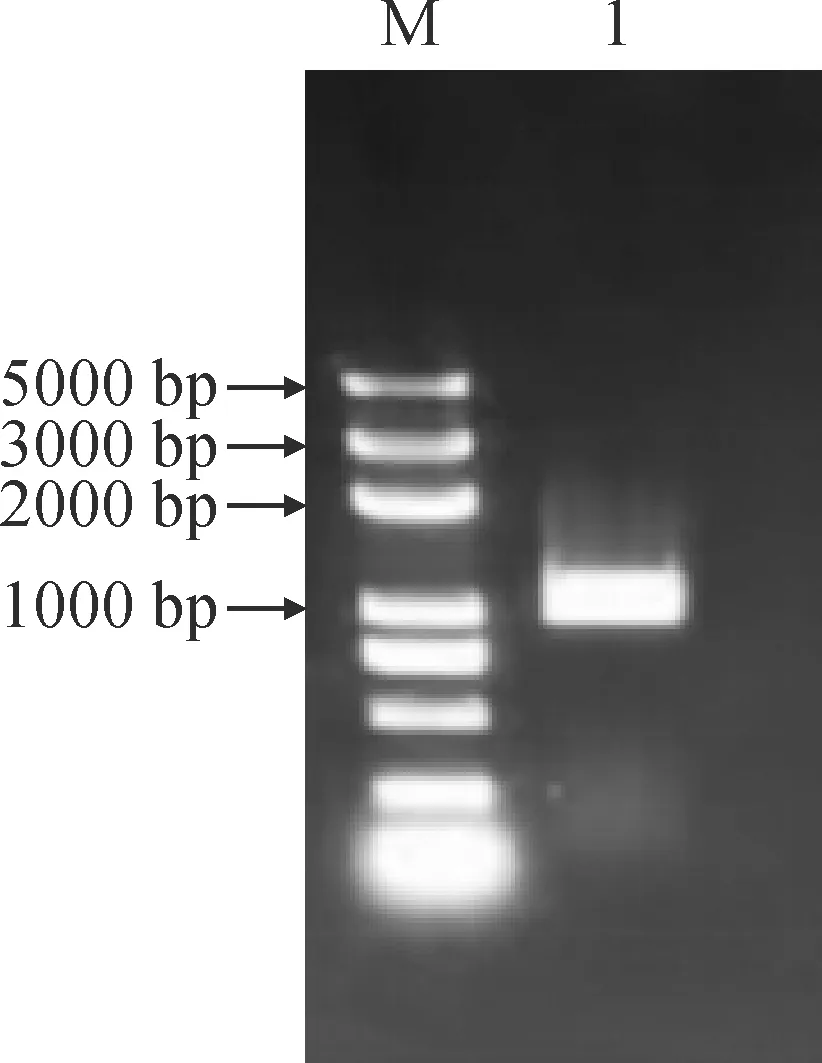
M. DL5000;1.MfMYB30

圖2 MfMYB30基因的完整開放閱讀框及推測氨基酸序列
2.2 黃花苜蓿MfMYB30蛋白結構特征分析
利用Expasy-ProtParam tool對MfMYB30基因的蛋白質理化性質進行預測,結果表明,分子量為86.85 kD,理論等電點為5.11,分子式為C3257H5456N1060O1363S174,氨基酸組成為Ala(33.9%)、Cys(16.4%)、Gly(21.2%)、Thr(28.5%),脂肪系數為33.87,不穩定系數為44.43,該蛋白為不穩定蛋白。利用Protscale預測MfMYB30基因編碼蛋白的親疏水性,結果顯示,最大親水分值為-0.633,最大疏水分值為2.033,總平均親水指數(grand average of hydropathicity, GRAVY)為0.736,為疏水性蛋白。利用SMART分析MfMYB30蛋白的保守結構域,有2個SANT保守結構(分別位于13~63 bp和66~114 bp)和1個低復雜性結構域(Low-complexity domain, LCD),均具有與MYB家族相似的結構(圖3)。MfMYB30 蛋白的信號肽以及跨膜結構域預測分析表明,MfMYB30蛋白無信號肽和跨膜結構域。

A.MfMYB30蛋白的保守域分析; B.MfMYB30蛋白質的二級結構:藍色.α螺旋;紅色.延伸鏈;紫色.無規則卷曲;綠色.β轉角
通過 SPOMA 在線工具分析二級結構,預測表明MfMYB30含有33.02%的α-螺旋,5.66%的延伸鏈,59.75%的不規則卷曲。α-螺旋對于黃花苜蓿MfMYB30蛋白質結構來說具有一定穩定性,無規則卷曲在功能上充當酶和蛋白質的活性中心(圖4)。

Mf.黃花苜蓿;Mt.蒺藜苜蓿;Cc.木豆;Gs.野大豆;Mp.刺毛黛豆;Ss.密花豆;Va.赤豆;Vr.綠豆;Vu.豇豆
2.3 MfMYB30基因的元件分析
利用Plant CARE在線軟件分析表明(表2),在MfMYB30基因上游啟動子區含有基本元件TATA-box和CAAT-box,光響應順式作用元件 G-box、Box-4、LAMP-element和MRE,與抗病、再生相關的響應元件P-box、TCA-element和ARE,此外,還有與轉錄因子MYB結合位點有關順式作用元件Box Ⅲ、MYB和MYB-like sequence。
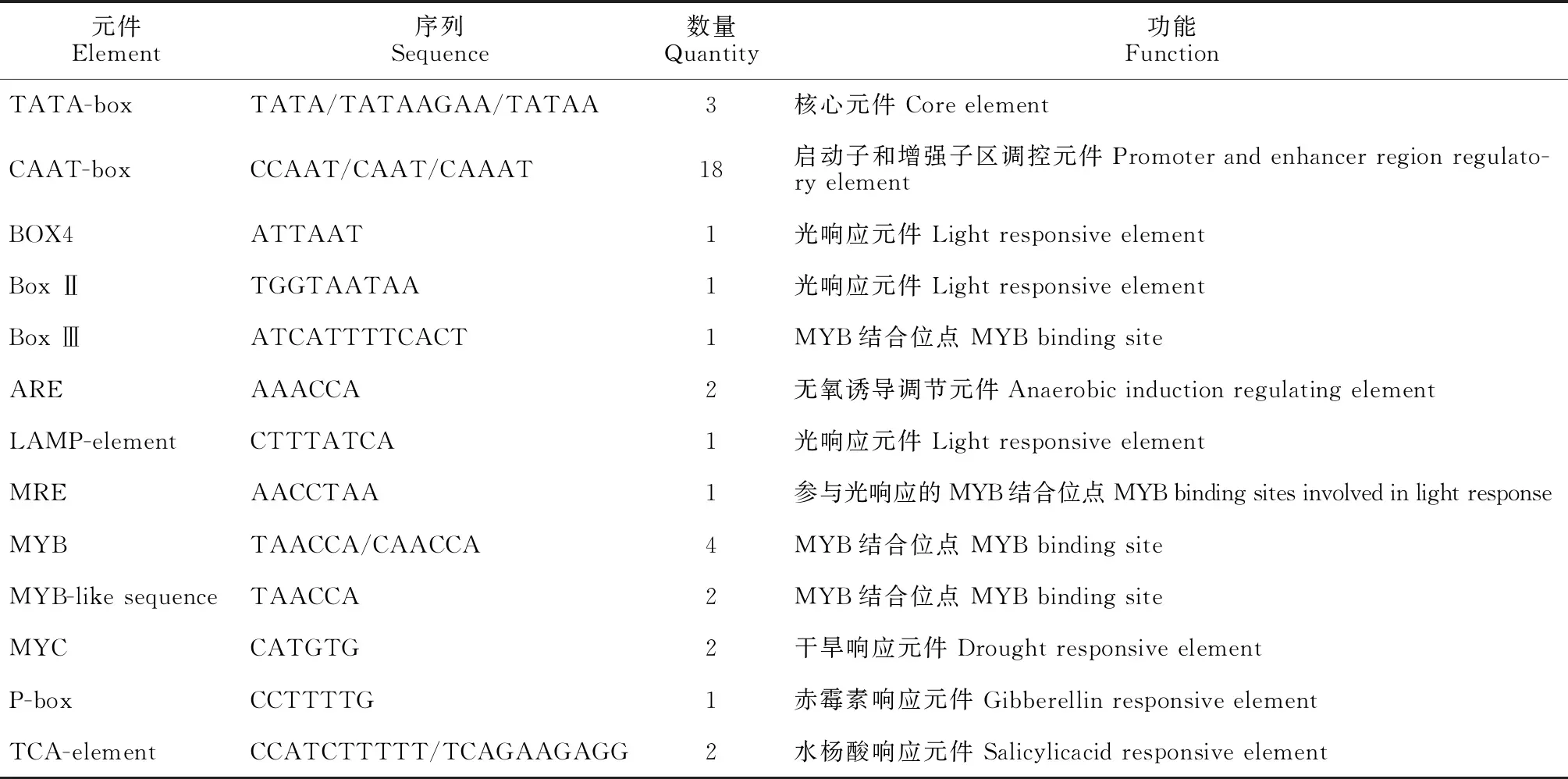
表2 MfMYB30基因啟動子序列順式作用元件
2.4 MfMYB30編碼氨基酸序列比對與系統進化樹分析
將MfMYB30氨基酸序列提交至NCBI在線BLASTn檢索,利用DNAMAN軟件對綠豆、豇豆、野大豆等8種豆科植物的MYB30蛋白進行多序列比對分析,結果表明,MfMYB30編碼的氨基酸序列與不同豆科植物MYB30氨基酸序列表現出高度相似性,大約在65%~99%之間,其中與同科植物蒺藜苜蓿MtMYB(Medicagotruncatula, XP_003618488.1)的氨基酸序列相似性最高,相似度達到了99%(圖5)。
為了更好地了解MfMYB30蛋白的系統發育關系,將大豆(14個)、蒺藜苜蓿(5個)、紫花苜蓿(3個)、擬南芥(5個)和木豆(2個)MYB蛋白,用MEGA7.0軟件與ITOL軟件對30個MYB蛋白構建系統進化樹(圖5),根據進化樹分支, 將30 個MYB 轉錄因子劃分為6類, 其中黃花苜蓿MfMYB30蛋白與紫花苜蓿MsMYB4、擬南芥AtMYB30、木豆CcMYB30、蒺藜苜蓿MtMYB30和大豆GmMYB60聚為一類,表明氨基酸序列差異較小,親緣關系較近,具有較高的相似性, 在功能上可能也具有相似性。
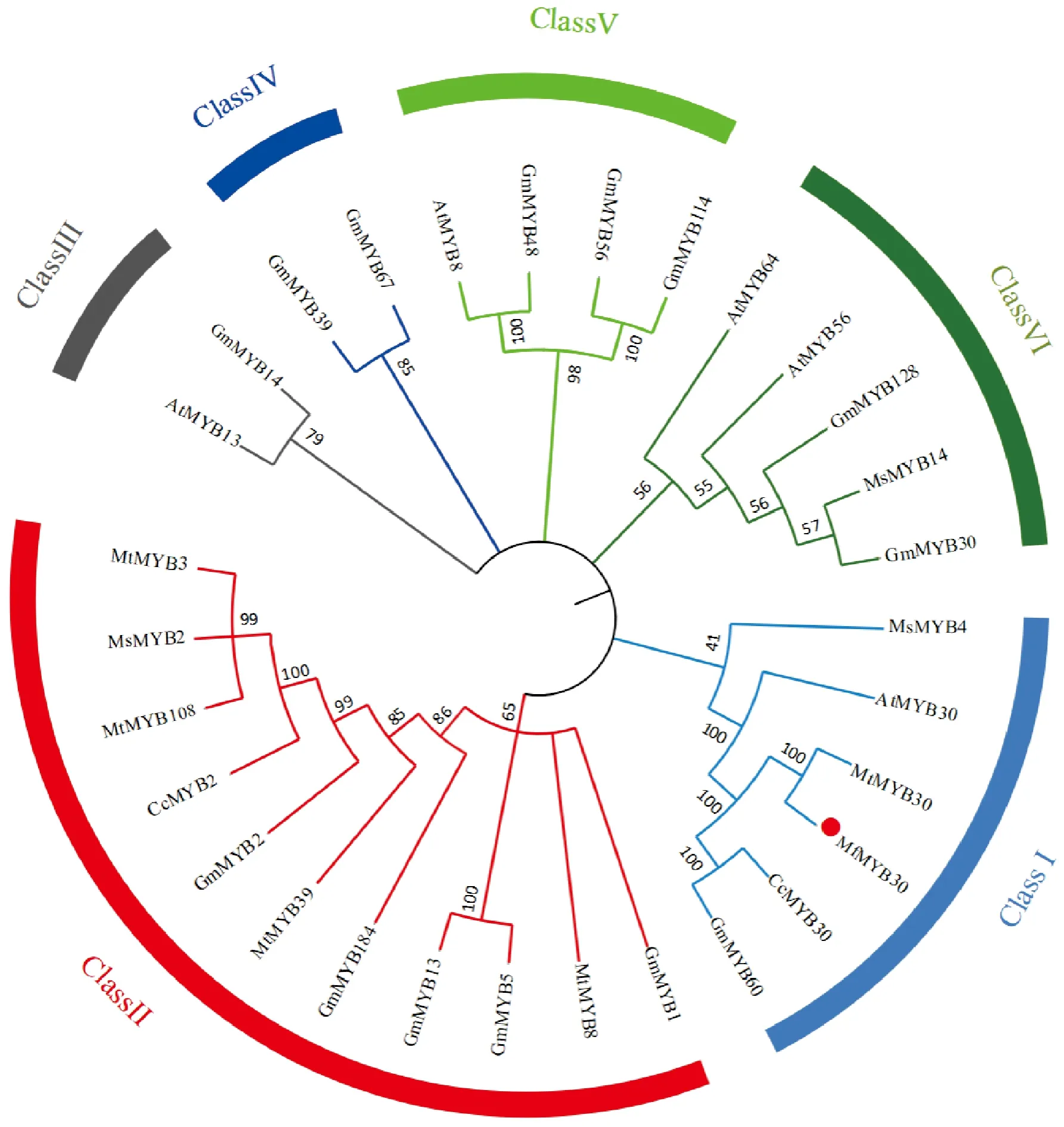
Mf.黃花苜蓿;Mt.蒺藜苜蓿;Ms.紫花苜蓿;Cc.木豆;Gm.大豆;At.擬南芥
2.5 黃花苜蓿MfMYB30基因的表達分析
為初步分析MfMYB30基因的功能,采用實時熒光定量PCR方法,以刈割脅迫不同天數后的黃花苜蓿葉片與莖組織的cDNA為模板,并以黃花苜蓿Actin基因作為內參基因,檢測黃花苜蓿刈割(模擬放牧)不同天數MfMYB30基因的表達情況。結果表明:在模擬刈割脅迫不同天數處理下,MfMYB30基因相對表達量呈現“降-升-升”的變化趨勢,在刈割7 d后達到峰值(圖6)。說明刈割(模擬放牧)處理對MfMYB30的表達有影響,黃花苜蓿面對非生物脅迫(刈割或放牧)時MfMYB30可能具有一定的調節功能。
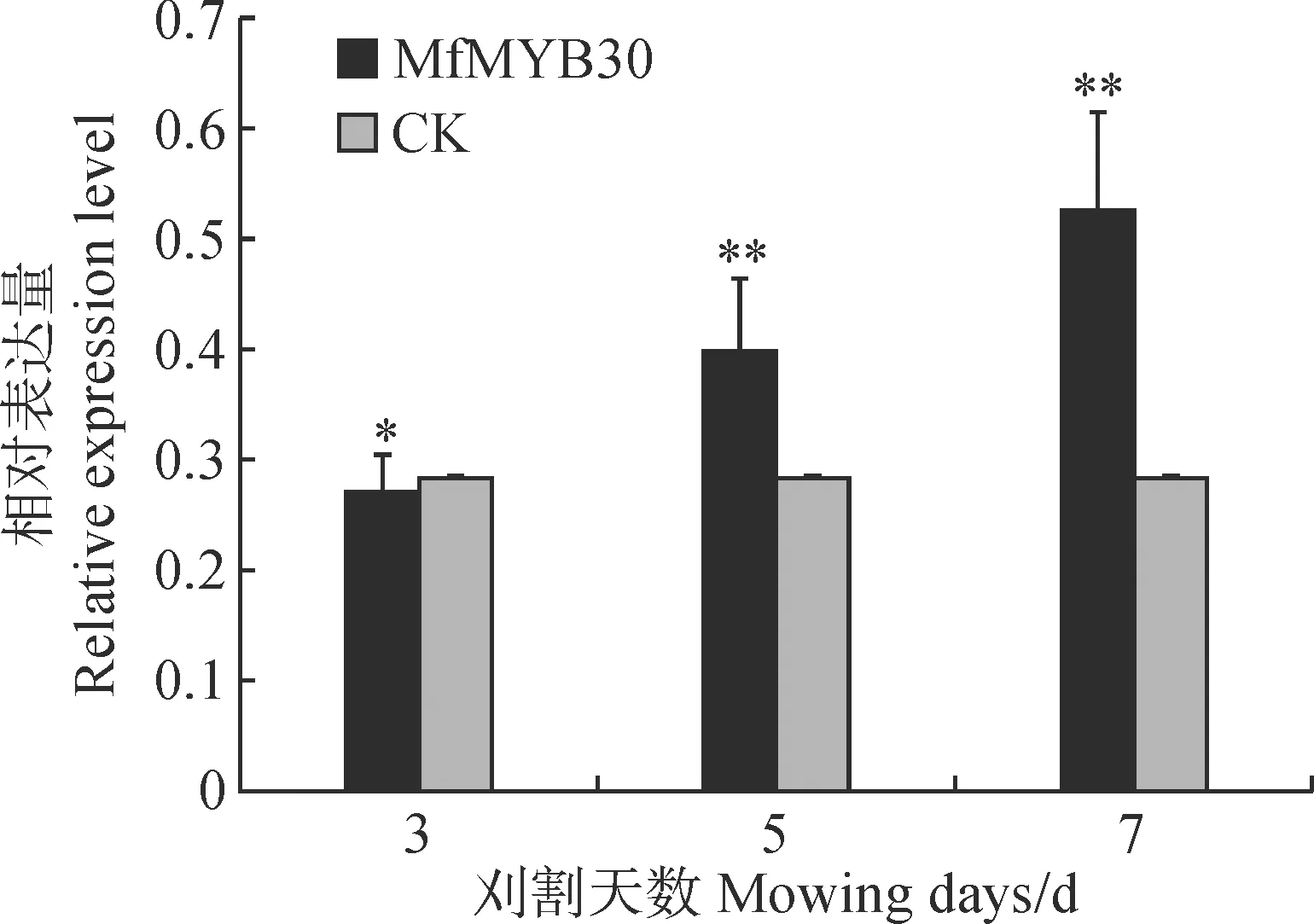
星號分別表示處理與對照之間基因表達水平存在顯著性差異(*. 0.01 通過WolF PSORT Protein Subcellular localization Prediction在線工具對MfMYB30蛋白進行亞細胞定位預測,分析該蛋白位于細胞的細胞核中。為進一步驗證MfMYB30蛋白的作用機制,我們構建了以pUC19為骨架的pUC19-HA-MfMYB30-GFP瞬時表達載體,通過擬南芥原生質體瞬時轉染技術進行了亞細胞定位分析,利用激光共聚焦顯微鏡觀察顯示,單獨的GFP熒光呈現彌散的狀態,而轉入MfMYB30-GFP融合蛋白后,熒光聚集在細胞核中,進一步證實MfMYB30蛋白亞細胞定位在細胞核中(圖7)。 圖7 MfMYB30蛋白的亞細胞定位 MYB類是一類重要的轉錄因子,在植物生長發育和抗逆過程中起著重要作用。其中,R2R3-MYB亞家族是植物中發現的最豐富的MYB家族,在植物特有的信號通路中,初生和次生代謝、細胞分化、形態發育和對生物脅迫和非生物脅迫的響應等發揮著重要作用[18]。 本研究根據轉錄組測序數據基礎篩選差異表達基因,克隆得到黃花苜蓿一個MYB(MfMYB30)基因。結構分析表明,MfMYB30編碼的蛋白由318個氨基酸組成,N端具有2個SANT 結構,2個結構域分別由51和49個氨基酸組成,屬于典型的R2R3-MYB轉錄因子結構特征。通過分析其編碼的氨基酸序列,發現MfMYB30蛋白與蒺藜苜蓿MtMYB30蛋白具有很高的相似性,在系統進化上,該蛋白與蒺藜苜蓿MtMYB30親緣關系較近,并且與處于進化樹同一分枝的其他植物的同源蛋白具有相同的保守結構域,說明MfMYB30在進化過程中具有功能上的高度保守性。亞細胞定位結果分析表明, MfMYB30蛋白定位在細胞核。 刈割是牧草的主要利用方式之一,科學合理的刈割制度是確保牧草生產力和品質,保證草地畜牧業的可持續發展的重要前提[3,19],當植物受到刈割脅迫后,容易受到病原菌的感染,植物受病原菌侵染后,誘導表達的抗病蛋白能夠直接與病原體的侵染結構接觸,抑制、干擾病菌的生長進程,激發植物產生系統獲得性抗性,發揮廣譜抗病作用[20-21]。前人研究表明,MYB30轉錄因子是通過調控乙酰輔酶A(acyL-CoA)延伸酶復合體基因的表達,進而調控超長鏈脂肪酸(Very-Long-Chain Fatty Acid,VLCFA)合成基因的表達以及HR反應,包括 KCS、FDH、KCR、ECR、PAS2/HCD、CER2、CER3 和 LTPG1[22]。而MYB30同時也能調控抗病相關激素水楊酸(SA)的合成[23]。在BR(Brassinosteroids)信號通路中,MYB30和BES1(BriL.-Ems-Suppressor 1)相互作用,共同調控了BR響應基因的表達,從而影響模式植物擬南芥下胚軸的伸長[24]。脫落酸(ABA)是被認為是非生物和生物脅迫反應的關鍵調控因子,MYB30是ABA信號通路的正調節物,它能被SUMO E3連接酶SIZ1所SUMO化,這對MYB30在ABA信號通路中的作用非常重要,MYB30可以調控脫落酸信號下的根細胞伸長[25-26]。還有研究表明,MYB30能調控擬南芥蠟粉的合成,表皮蠟粉在植物抵御病蟲害和降低紫外線福射等方面起著重要作用[27-29]。通過上述分析,黃花苜蓿MfMYB30基因可能在響應刈割脅迫反應起作用,提高植物的抗逆性,進而促進植物的再生。 實時熒光定量PCR是一種快速準確的核酸定量分析技術,具有特異性強、靈敏度高、速度快等優點。本研究RT-PCR的結果進一步表明了MfMYB30很有可能參與黃花苜蓿中刈割脅迫調控,在刈割3、5和7 d處理下,MfMYB30在5 d和7 d后的表達均有明顯上調。由此推測在刈割脅迫條件下MfMYB30可能起正調控作用。 由于在生命活動中真正起作用的是基因的表達產物蛋白質,而蛋白質的功能與其在細胞中的亞細胞定位密切相關,蛋白質位于細胞的不同部位所行使的功能也不同,因此對蛋白質進行亞細胞定位是研究其功能必不可少的環節之一[30]。為了進一步探究MfMYB30的作用機制,確定黃花苜蓿MfMYB30蛋白在細胞中的存在位置,本實驗構建了pUC19-HA-MfMYB30-GFP瞬時表達載體,并通過擬南芥原生質體瞬時轉染技術進行分析,在激光共聚焦顯微鏡下觀察并拍照。結果表明,MfMYB30定位于細胞核中,是一個核蛋白。 綜上所述,本研究從黃花苜蓿轉錄組中篩選和克隆獲得1個表達顯著上調的MfMYB30基因,并通過生物信息學方法和RT-qPCR分析對其生物學特性與功能進行研究,以期為闡明黃花苜蓿MfMYB30基因在響應刈割(或放牧)脅迫的分子機制提供理論基礎。同時通過RNA-Seq測序和數據分析發現,在獲得的黃花苜蓿轉錄組數據中除MYB轉錄因子外還存在其他差異表達基因,挖掘和解析這些重要基因的功能有助于進一步提高黃花苜蓿的抗逆性,也為利用分子生物學手段培育抗逆性強的黃花苜蓿品種奠定基礎。2.6 黃花苜蓿MfMYB30蛋白的亞細胞定位
3 討 論
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
電子制作(2018年18期)2018-11-14 01:48:24
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
