克氏針茅物候對氣候變暖和水分變化的響應及其光合生理生態機制
2022-11-12 03:29:02顧文杰周廣勝呂曉敏周夢子汲玉河周懷林
生態學報 2022年20期
顧文杰,周廣勝,2,3,4,*,呂曉敏,2,4,周夢子,2,4,周 莉,2,4,汲玉河,2,4,周懷林,4
1 中國氣象科學研究院, 北京 100081 2 鄭州大學地球系統科學與技術研究院生態氣象聯合實驗室, 鄭州 450001 3 南京信息工程大學氣象災害預報預警與評估協同創新中心, 南京 210044 4 河北固城農業氣象國家野外科學觀測研究站, 保定 072656
植物物候是植物長期適應溫度、降水等環境條件季節性變化所形成的彼此相適應的生長發育節律[1],它不僅是植物生長發育狀況的直觀反映[2],也是指示氣候變化的重要指標[3—4],已經廣泛應用于指導農事活動及防災減災[5]。同時,植物物候作為影響生態系統結構與功能的重要參數[6—7],在調節陸地生物化學循環中發揮著重要作用[8],是陸面過程模型及植物生產力模型的重要參數[9]。因此,研究植物物候變化規律及其調控機制對物候的準確預測及科學應對氣候變化均具有重要意義。
已有研究表明,全球植物春季物候整體呈提前趨勢,而秋季物候呈推遲趨勢[10—11],且這種現象在北半球高緯度地區更為顯著[12]。然而,研究發現內蒙古草原植物物候對氣候變化的響應存在顯著的空間差異性,南部地區植物物候呈現出返青期提前,枯黃期推遲的趨勢;而中北部地區則呈現出相反的變化趨勢[13]。區域氣候的差異及植物-氣候相互作用的復雜性導致植物物候研究仍存在很大的差異性與不確定性[14],亟需開展植物物候對氣候變化響應的生理生態機制研究。溫度[15]、降水[16]、光照[17]、空氣濕度[18]、CO2濃度[19]等環境因子是影響植物物候的重要因素,尤其是溫度和水分。溫度被認為是影響植物物候最重要的環境因子[20],植物只有在一定溫度環境中才能生長發育,且需要一定的積溫才能完成其生活周期[21]。水分不足限制植物對光、熱條件的利用效率[16],被認為是調節干旱半干旱地區植被活動的關鍵因子[22]。并且,溫度與降水之間的相互作用通過改變環境因子的可利用性[23]或植物對環境因子的敏感性[24]顯著影響植物物候變化。然而,現有研究大多關注物候變化與環境因子的關系,關于植物物候變化的生理生態機制仍然不清楚[25]。近來研究表明,物候變化與其凈光合速率、蒸騰速率、氣孔導度等光合生理生態因子密切相關[26],但仍缺乏植物物候對水熱環境變化響應的光合生理生態機制研究。
中國溫帶草原是世界第三大草原區[27],克氏針茅草原是該地區特有的草原群系,是典型草原的代表類型之一,在畜牧業生產中占有重要的地位[28]。并且,克氏針茅草原生態系統脆弱,植物物候已經受到氣候變化的顯著影響[29]。因此,本研究試圖以克氏針茅為研究對象,利用增溫與控水相結合的原位控制試驗資料,分析研究克氏針茅物候對氣候變暖和水分變化的響應及其光合生理生態機制,以增進物候對環境變化響應的理解,并為物候模型發展提供依據。
1 研究區域與研究方法
1.1 研究區概況
試驗于2020年在中國氣象局內蒙古錫林浩特國家氣候觀象臺(44°08′03″ N、116°19′43″ E,海拔990 m)開展。該地區位于內蒙古典型草原中部,是典型的溫帶半干旱大陸性氣候區,1955—2015年間該地區年均氣溫為2.5 ℃,年均降水量為283.6 mm[30],冬季寒冷干燥、夏季溫暖濕潤,太陽輻射較強。試驗樣地地勢平坦開闊,土壤類型主要為淡栗鈣土,土壤腐殖質層較薄。優勢植物有克氏針茅(Stipakrylovii)和羊草(Leymuschinensis),重要伴生種包括細葉蔥(Alliumtenuissimum)、糙隱子草(Cleistogenessquarrosa)、冷蒿(Artemisiafrigida)、矮蔥(Alliumamsopodium)、木地膚(Kochiaprostrata)、黃蒿(Artemisiascoparia)、阿爾泰狗娃花(Heteropappusaltaicus)等。
1.2 試驗材料與設計
試驗以克氏針茅草原作為試驗對象,采用紅外線輻射增溫與控水相結合的原位控制實驗方法。該地區氣溫呈顯著上升趨勢,但降水年際間波動大,總體趨勢不明顯[31],未來暖濕化與暖干化現象均有可能出現。因此,以該地區的自然溫度和降水為對照,設置5個溫度和降水處理:對照(T0W0)、輕度干熱(T+1.5W-50%)、重度干熱(T+2W-50%)、輕度濕熱(T+1.5W+50%)、重度濕熱(T+2W+50%) (表1)。每個處理均設置4個重復小區,共20個小區。小區面積為4 m2(2 m×2 m),相鄰小區間隔2 m。小區上方安裝有長3 m、寬3 m的遮雨棚。小區四周用高0.3 m、深1 m的隔水鐵板圍起,以減小地表和地下水分流動的影響。
采用不同功率的紅外線輻射增溫裝置對小區進行增溫處理。選用的紅外輻射燈管長為1 m,平行地面懸掛于小區中心,離地高度為2 m。利用不同遮雨量的遮雨棚裝置和灌溉進行水分處理。遮雨板材質為透光率95%以上透光PC板,分為帶孔遮雨板和不帶孔遮雨板兩種類型。通過改變遮雨棚帶孔遮雨板和不帶孔遮雨板的比例調控遮雨棚的遮雨量。每次降水時還需收集減水處理小區(T+1.5W-50%、T+2W-50%)遮雨棚截留的降水,并將其均勻灌溉到增水處理小區(T+1.5W+50%、T+2W+50%) (表1)。
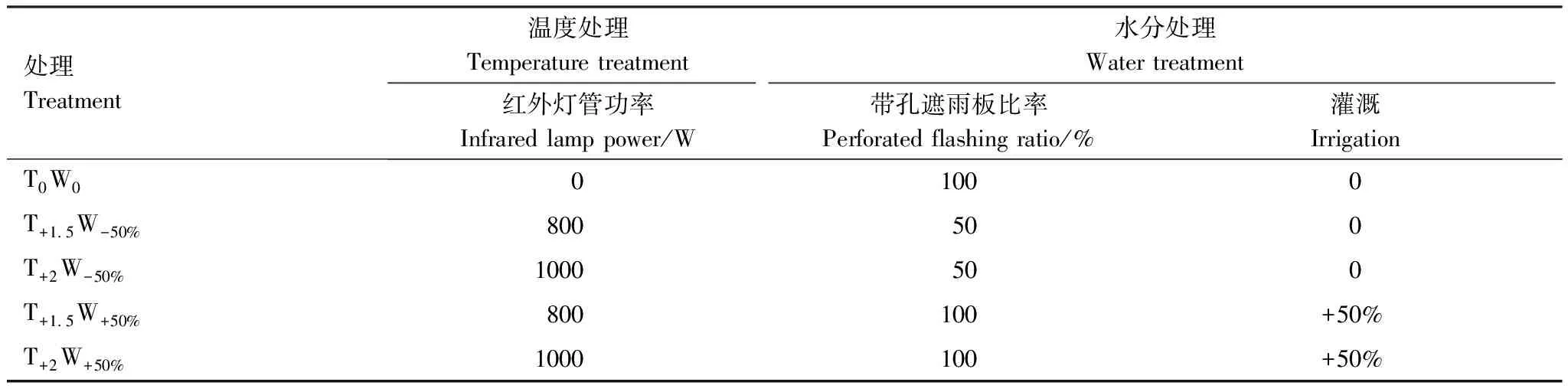
表1 不同小區的水熱處理方案
1.3 測定項目與方法
1.3.1土壤溫濕度
利用ECH2O土壤溫濕度檢測系統(METER, Pullman, WA, USA)測定各試驗小區的土壤溫度(Tsoil, ℃)和濕度(RHsoil,%)。該系統觀測頻率為30 min/次,利用熱敏電阻測定各小區0—10 cm的土壤溫度,并通過電容/頻域技術測量0—30 cm每10 cm一層的土壤分層體積含水量。
1.3.2物候觀測
依據《農業氣象觀測規范》[32]對克氏針茅物候進行人工觀測。每日上午和下午分別對試驗樣地小區進行巡視,記錄各小區克氏針茅到達各關鍵物候期的日期,并將日期記錄轉換為同年1月1日開始計算的日序。當觀測小區50%克氏針茅的葉片恢復彈性,由黃轉青,則認為達到返青期;當小區50%克氏針茅的針茅從葉鞘頂端或側端露出,記錄到達抽穗期;當小區50%克氏針茅的地上器官約有三分之二枯萎變色,記錄為枯黃期。
1.3.3葉片光合生理生態參數
為研究氣候變暖和水分變化情景下克氏針茅光合生理生態特征的變化,并進一步分析植物物候響應氣候變化的光合生理生態機制,利用Li- 6400便攜式光合作用系統(LI-COR, Lincoln, NE, USA)測定克氏針茅各關鍵物候期的光合生理生態參數,測定內容包括葉片凈光合速率(Pn, μmol m-2s-1)、氣孔導度(Gs, mol m-2s-1)和蒸騰速率(Tr, mmol m-2s-1)等。當克氏針茅到達返青期、抽穗期和枯黃期時,于晴朗無風天氣的9:30—11:30進行觀測。測定時,每個小區選取1株具有代表性并到達該物候期的克氏針茅,將葉片平展且不重疊地鋪滿葉室,測定部位為葉片中部,依據儀器使用手冊將空氣流速設定為300 mmol/s;葉室CO2濃度控制為大氣CO2濃度,即400 μmol/mol。光照強度的設定因物候期而不同,通過測定克氏針茅葉片的光響應曲線確定,返青期為1200 μmol m-2s-1,抽穗期為1500 μmol m-2s-1,枯黃期為2000 μmol m-2s-1。葉片水分利用效率(WUE, mmol/mol)計算如下[33]:
WUE=Pn/Tr
(1)
1.4 數據處理
利用Microsoft Excel 2019和SPSS 21.0進行數據整理、匯總與分析。采用配對樣本t檢驗方法,分析不同水熱處理下小區土壤溫濕度的差異;利用雙因素方差分析方法,分析增溫和水分對克氏針茅物候的影響;溫度、水分和物候期對克氏針茅光合生理生態特征的影響則采用三因素方差分析方法,并用Duncan多重比較方法分析不同處理間物候及光合生理生態特征差異。為闡明光合生理生態參數和土壤溫濕度對克氏針茅物候的直接或間接作用方式及影響,利用SPSS AMOS 21.0軟件構建通徑分析模型進行分析。
2 結果與分析
2.1 試驗小區土壤溫濕度變化特征
各試驗小區土壤溫度和濕度在克氏針茅生長季內的波動趨勢較為一致,但是土壤溫度和濕度在不同增溫和控水處理之間存在顯著差異(圖1)。與對照相比,增溫處理小區的土壤溫度顯著升高;增水處理(T+1.5W+50%、T+2W+50%)小區的土壤濕度顯著高于對照,減水處理(T+1.5W-50%、T+2W-50%)小區的土壤濕度顯著低于對照。并且,增溫處理和水分處理間存在相互影響,在相同溫度處理下增水處理小區的土壤溫度顯著低于減水處理小區;在相同水分處理下,增溫2 ℃處理小區的土壤濕度顯著低于增溫1.5 ℃處理小區。
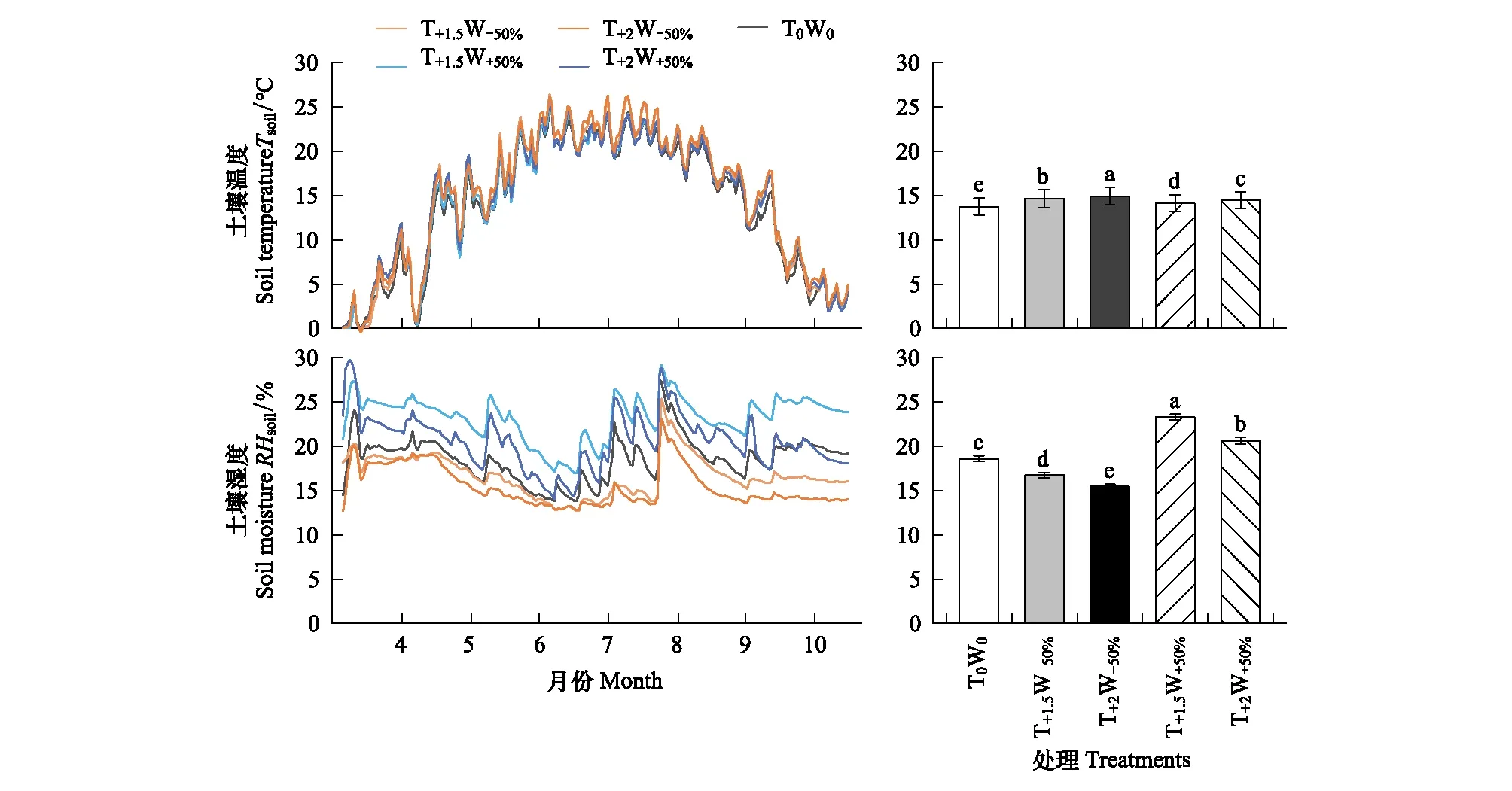
圖1 不同增溫和控水處理下土壤溫度和濕度的動態變化及生長季平均土壤溫度和濕度的變化Fig.1 Changes of soil temperature and moisture & average soil temperature and moisture under different warming and precipitation treatments不同小寫字母表明不同處理間差異顯著(P < 0.05)
2.2 克氏針茅物候期變化
不同增溫和水分處理下,克氏針茅到達各物候期的時間存在顯著差異(表2)。與對照相比,T+1.5W-50%、T+2W-50%、T+1.5W+50%、T+2W+50%處理的返青期分別提前2 d、3.7 d、2 d和3.5 d,抽穗期分別提前3.8 d、4.5 d、10.5 d和12.5 d,枯黃期分別推遲6.3 d、7.7 d、6 d和7d,導致生長季分別延長8.3 d、11.4 d、9 d和10.5 d。雙因素方差分析表明,溫度是影響克氏針茅返青期物候的重要因素,水分主要影響克氏針茅的抽穗期,溫度和水分協同作用對各物候期的影響不顯著(表3)。Duncan多重比較表明,不同水分處理間克氏針茅的抽穗期差異顯著,增溫使枯黃期顯著推遲(表2)。
2.3 克氏針茅光合生理生態特征變化
不同水熱處理下,克氏針茅各物候期的光合生理生態特征差異明顯(圖2)。在克氏針茅的返青期,增溫增水處理(T+1.5W+50%、T+2W+50%)的凈光合速率、氣孔導度和蒸騰速率均顯著高于對照(P< 0.05),水分利用效率低于對照,但未達到顯著水平;增溫減水處理(T+1.5W-50%、T+2W-50%)的凈光合速率略高于對照,但蒸騰速率低于對照。減水處理使克氏針茅的水分利用效率提高,且在重度干熱處理(T+2W-50%)更為顯著(P<0.05)。
與返青期相比,不同水熱處理下克氏針茅抽穗期的凈光合速率、氣孔導度和蒸騰速率均明顯下降。增溫增水處理(T+1.5W+50%、T+2W+50%)的凈光合速率、氣孔導度和蒸騰速率均高于對照和增溫減水處理(T+1.5W-50%、T+2W-50%),且在增溫和水分協同作用下T+1.5W+50%處理的凈光合速率、氣孔導度和蒸騰速率顯著高于T+2W+50%處理(P<0.05)。水分利用效率對增溫增水處理的響應不顯著,但增溫減水處理使克氏針茅的水分利用效率顯著升高(P< 0.05)。在枯黃期,克氏針茅的光合能力進一步降低,不同水熱處理間的克氏針茅光合生理生態特征差異減小。增溫處理下,克氏針茅的凈光合速率及水分利用效率顯著高于對照(P< 0.05),表明枯黃期的推遲與克氏針茅的光合能力提高密切相關。
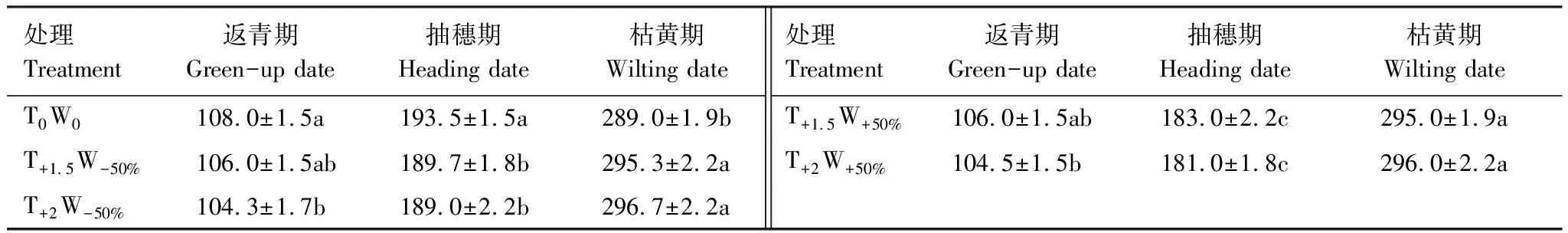
表2 不同增溫和控水處理下克氏針茅物候的變化

表3 不同增溫和控水處理下克氏針茅物候的雙因素方差分析 (F值)
三因素方差分析表明,不同物候期的溫度和水分協同作用使得克氏針茅的光合生理生態特征變化顯著(表4)。不同物候期的克氏針茅的凈光合速率、蒸騰速率、氣孔導度和水分利用效率差異顯著,水分變化顯著影響克氏針茅的蒸騰速率、氣孔導度、凈光合速率和水分利用效率,但溫度更多與物候協同作用影響克氏針茅的光合生理生態特征。
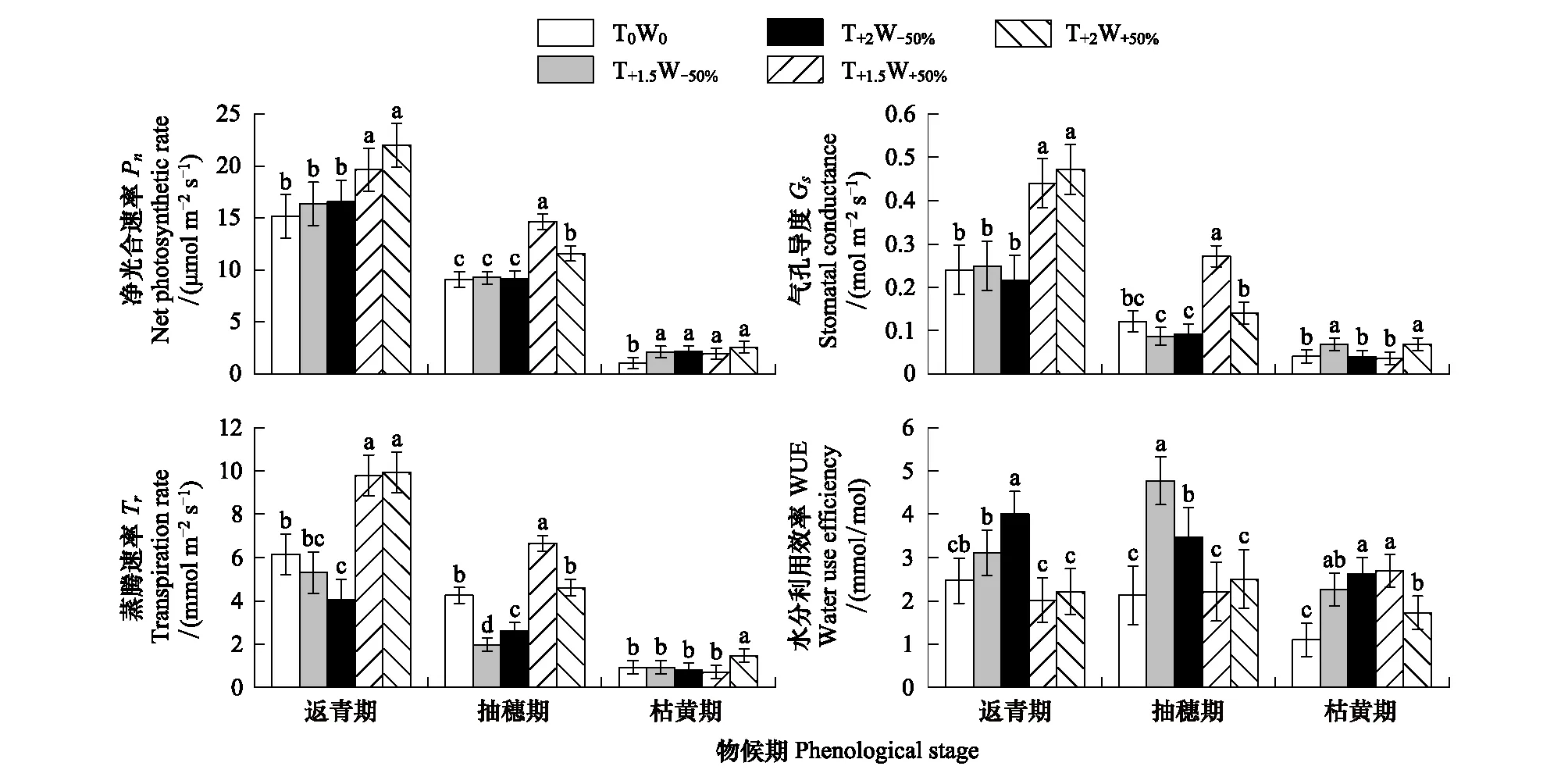
圖2 不同處理下克氏針茅主要物候期的凈光合速率、氣孔導度、蒸騰速率和水分利用效率的變化Fig.2 Changes in net photosynthetic rate, stomatal conductance, transpiration rate and water use efficiency of main phenological stages of Stipa krylovii under different treatmentsGS:返青期 Green-up stage;HS:抽穗期 Heading stage;WS:枯黃期 Wilting stage;不同小寫字母表明不同處理間差異顯著(P<0.05)

表4 不同處理下克氏針茅光合生理生態特征的三因素方差分析結果 (F值)
2.4 克氏針茅物候變化的光合生理生態機制
通徑分析(圖3)表明,光合生理生態參數、土壤溫度和土壤濕度對克氏針茅的返青期、抽穗期和枯黃期的解釋率分別為95%、58%和62%,且對克氏針茅物候期直接作用最大的因子均為凈光合速率,說明凈光合速率是導致克氏針茅物候變化的主要光合生理生態因子。返青期和抽穗期的凈光合速率對克氏針茅物候的直接效應分別為-0.961和-0.864,表明凈光合速率的增加促進返青期和抽穗期的提前;枯黃期的凈光合速率對克氏針茅物候的直接效應為0.790,表明凈光合速率的增加導致枯黃期推遲。返青期的土壤溫度對物候的間接效應主要通過水分利用效率影響蒸騰作用及氣孔導度,進而影響凈光合速率,溫度升高促使克氏針茅的返青期提前(-0.191)。抽穗期的土壤溫度對物候的間接效應(-0.181),主要通過水分利用效率影響蒸騰作用,而土壤濕度對物候的間接效應(-0.750),主要是通過氣孔導度影響凈光合速率。枯黃期的土壤溫度通過影響凈光合速率間接影響植物物候(0.256),土壤溫度對枯黃期的總效應為正值,表明溫度升高導致枯黃期物候推遲(表5)。
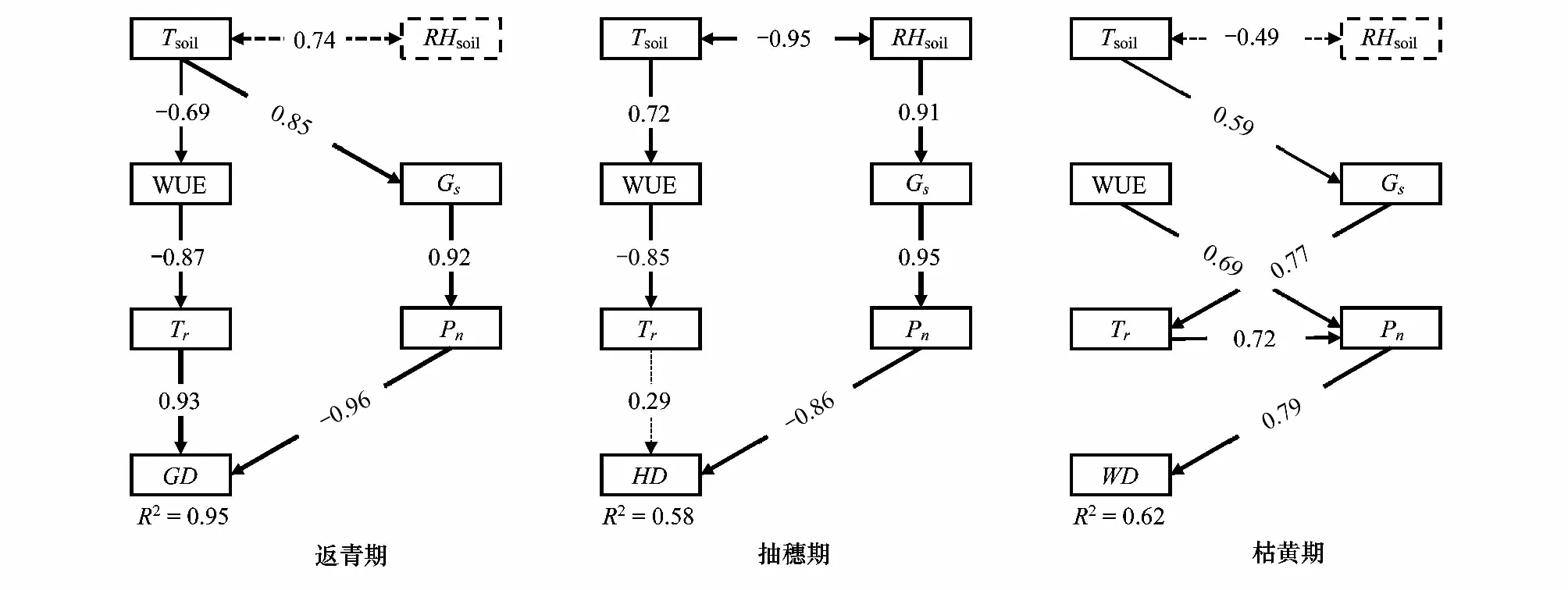
圖3 克氏針茅物候和光合生理生態因子的通徑分析Fig.3 Path analysis of phenology and photosynthetic ecophysiological characteristics of Stipa kryloviiTsoil:土壤溫度 Soil temperature;RHsoil:土壤濕度 Soil moisture;WUE:水分利用效率 Water use efficiency;Gs:氣孔導度 Stomatal conductance;Tr:蒸騰速率 Transpiration rate;Pn:凈光合速率 Net photosynthetic rate;GD:返青期 Green-up date;HD:抽穗期 Heading date;WD:枯黃期 Wilting date;----:P < 0.05;:P > 0.05

表5 克氏針茅物候和光合生理生態因子的通徑分析
3 討論與結論
以氣候變暖為主要特征的全球氣候變化已經成為事實[34],已經影響土壤理化性質和植物生理生態特征,進而影響土壤-植物系統[35]。本研究基于紅外線輻射增溫與控水相結合的原位模擬試驗發現,土壤溫度和濕度之間存在相互影響。已有研究表明,氣候變暖會導致土壤溫度升高,對土壤濕度的影響則表現出顯著的季節差異[36]。夏季增溫導致蒸散加強,土壤濕度降低[37];冬季增溫則增大土壤與大氣間的溫差,使水汽更易液化滲入土壤,導致土壤濕度升高[35]。
氣候變化已經導致草原植物物候發生顯著改變。1965—2013年,青藏高原地區牧草返青期、抽穗期和開花期平均每10年分別提前7.4 d、6.2 d和6.4 d,而枯黃期則呈現推遲趨勢(2.1 d/10 a),溫度升高被認為是導致物候變化的主要原因[38]。但受氣候變暖影響,內蒙古通遼地區氣候暖干化,水分脅迫成為導致該地區羊草、冰草(Agropyroncristatum)和委陵菜(Potentillachinensis)返青期推遲,枯黃提前的重要原因[39]。生存分析模型表明,氣溫升高2 ℃將導致草原植物返青期提前1—4 d[40];開頂式箱(OTC)增溫模擬表明,增溫2.2 ℃使釘柱委陵菜(Potentillasaundersiana)的生殖期提前3—5 d,但使高山嵩草(Kobresiapygmaea)的生殖期推遲5—7 d,增溫導致的水分脅迫被認為是造成高山嵩草物候期推遲的原因[41]。本研究發現,增溫處理下克氏針茅植株的返青期提前2—4 d,抽穗期提前3—13 d,枯黃期推遲6—8 d,物候期變化趨勢與已有研究結果一致[42]。研究還發現,克氏針茅在抽穗期對水分變化敏感,增溫增水處理使抽穗期提前,較增溫減水處理提前6—9 d。
環境因子是影響植物生長發育的重要因素,也是植物生理生態變化的誘因。植物光合作用本質上是一系列酶促反應,溫度升高顯著提高植物光合作用的重要環節RuBP的羧化和再生的最大速率[43]。本研究也發現,增溫處理下克氏針茅的凈光合速率升高。但有研究表明,植物凈光合速率、氣孔導度和蒸騰速率對升溫的響應存在物種間差異,增溫促進了發草(Deschampsiacaespitosa)的光合作用,但抑制了遏藍菜(Thalaspiarvense)的光合作用[44]。水分是限制植物葉片光合作用的關鍵因素,降水增加將導致植物光合速率、氣孔導度和蒸騰速率升高,而水分脅迫將導致植物氣孔導度和蒸騰速率下降,水分利用效率提高[45],本研究也觀測到一致的現象。
植物物候是植物對溫度、降水等環境條件季節性變化的適應結果,是植物可利用氣候資源的綜合體現。植物的凈光合速率體現了植物利用氣候資源將CO2和水轉化為有機物的快慢[46]。本研究通過原位控制試驗,將環境因子、光合生理生態因子和植物物候聯系起來,并分析了克氏針茅物候對氣候變暖和水分變化的響應及其光合生理生態機制,得到以下結論:(1)溫度升高導致克氏針茅的返青期和抽穗期提前、枯黃期推遲,水熱協同作用對克氏針茅物候的影響主要體現在抽穗期。(2)在返青期和抽穗期,增溫增水導致克氏針茅的凈光合速率、氣孔導度和蒸騰速率顯著升高,而增溫減水則導致水分利用效率顯著升高。對于枯黃期,增溫使凈光合速率和水分利用效率升高,不同水分處理間光合生理生態特征無顯著差異。(3)不同物候期,影響克氏針茅物候的最重要直接因子均為凈光合速率,凈光合速率升高導致克氏針茅返青期、抽穗期提前,枯黃期推遲。(4)克氏針茅物候期與環境因子及其光合生理生態特征密切相關。凈光合速率是植物物候變化的決策因子,溫度和降水則是植物物候變化的限制因子。
致謝:感謝中國氣象局內蒙古錫林浩特國家氣候觀象臺對試驗提供的大力支持。
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
