基于環境DNA的日本河流生物多樣性文獻計量分析及其進展
2022-11-25 06:20:12劉澤軍胡茂川劉丙軍郝愛民井芹寧
西北水電 2022年5期
賀 凱,劉澤軍,胡茂川,劉丙軍,郝愛民,井芹寧
(1. 中山大學 土木工程學院,廣東 珠海 519082;2. 廣東省環境資源利用與保護重點實驗室,廣州 510640;3. 溫州大學 生命與環境科學學院,浙江 溫州 325035;4. 亞熱帶水環境生態保護浙江省國際科技合作基地,浙江 溫州 325035)
0 前 言
傳統水生生物監測主要依靠觀測者對物種進行形態學鑒定,因而對從事物種鑒定人員的專業知識要求較高,同時傳統的水生生物監測方法可能會給水生生物或者其所在的水生生態系統帶來一定程度的危害導致其具有一定的局限性[1]。而環境DNA技術作為新興的水生生物監測方法,基于當前基因測序技術,可以通過檢測游離在水環境中的DNA分子檢測出對應生物種類的存在。其相較于傳統水生生物形態學監測方法而言具有經濟、高效、準確、靈敏等優勢[2]。因此,近年來環境DNA檢測技術發展迅速,被廣泛應用于流域入侵物種、瀕危物種監測、生物量評估和生物多樣性調查等[3-5]。
環境DNA指在自然環境中廣泛分布的游離DNA分子,主要通過生物脫落的皮膚、唾液、糞便、配子、分泌物等方式獲得[6]。環境DNA分子大小不一,存在部分大于180 μm或小于0.2 μm的分子,DNA分子的大小可能與生物種類相關。運用環境DNA的水生生物調查研究始于1987年,由美國田納西大學研究團隊開發從環境(湖泊沉積物)中提取和純化細胞外DNA的方法[7],隨后法國Ficetola等[8]通過針對棲息地池塘中提取牛蛙(Ranacatesbeiana)DNA信息,以此研究牛蛙棲息習慣。Minamoto等首次運用環境DNA宏條形碼技術于魚類、兩棲動物、鳥類和哺乳動物等水環境生物監測研究[9]。Doi等也通過對鯉魚環境DNA分析檢測來證明環境DNA宏條形碼技術在魚類乃至水生生物的定量評估中的潛力[10-11]。 Miya開發具有代表性的通用引物“MiFish”用于環境DNA宏條形碼技術,目前該引物可檢測到200多個魚種[12-14]。日本通過出版環境DNA研究和分析手冊等[15-16]指導環境DNA技術應用研究,并建立基于大規模、高分辨率數據的生態系統解釋、預測和保護系統。
日本開展環境DNA研究較早,有著廣泛的應用案例。近年環境DNA研究也被引導強化同其他研究領域的融合與合作,如生態系統的可持續利用和環境保護。本文在系統介紹環境DNA技術及其影響因素的基礎上,梳理日本運用環境DNA技術在河流生態監測與多樣性評價方面的研究成果,旨在為中國未來生態監測和生物多樣性研究方面提供素材和借鑒。
1 日本環境DNA技術的發展歷程
日本在環境DNA技術的研究起步較早,Minamoto等[9]在對水族箱水樣進行DNA提取和分析后,確認所有水族箱魚種都被檢出。根據水族箱分析結果對3個自然河道現場采集的水樣開展環境DNA分析,并檢出自然河道中4個魚種,檢測結果同以往常規方法(觀察和捕捉方法)確定的魚種一致。環境DNA方法可以提供簡化的魚類等水生生物調查和監測,日本科學家在之后的研究中成功利用環境DNA技術檢出日本淡水湖中魚類的物種組成,證明該方法在日本水環境中魚類調查研究中的適用性。Takahara等[10]利用環境DNA技術在室內水箱和室外實驗池中發現鯉魚(CyprinuscarpioL.)豐度與水中環境DNA濃度之間存在正相關關系,并利用環境DNA對天然潟湖中的鯉魚生物量進行評估。從2008年至今,環境DNA技術在水生生態系統的研究經歷了定性到定量,由檢測單一物種到同時檢測多種物種,從調查淡水水域擴展到海洋,調查物種從小型底棲動物擴展到兩棲動物、哺乳動物等的發展歷程[17-20]。
為研究日本環境DNA技術發展歷程運用CiteSpace[21]軟件對日本相關文獻進行關鍵詞聚類分析,數據庫為Web of Science (WOS)核心數據庫(2022年6月28日),搜索策略為TS=((“environmental DNA”OR “eDNA”) AND (Japan OR Japanese)),共搜索文獻141篇。歷年文獻發表數量及年被引次數見圖1,從圖中可以看出環境DNA發文數量及年被引用次數均呈逐年上漲的趨勢,而發展突變點出現于2013—2016年,表明在該時間段環境DNA開始步入大眾科研關注領域。
2 環境DNA技術流程介紹及其影響因素
環境DNA技術分析主要包括樣品采集、樣品處理、DNA提取、PCR擴增、基因測序、生物信息學以及統計學分析。具體技術處理流程如圖2所示。

圖1 日本環境DNA發文數量及年被引次數

圖2 環境DNA技術處理流程
Jo等以環境DNA的生物放大作用,以馬鮫魚(Trachurusjaponicus)為標本,對不同水溫和魚類生物量條件下的DNA脫落和降解過程進行實驗,發現降解過程隨環境DNA大小發生變化,同時環境DNA濃度及其降解率也隨著水溫的升高和生物量增加而增加[22]。溝口等利用環境DNA技術對海上風電設施附近開展魚類監測,用來識別其周圍水域的魚種,對漁業影響評估方法的改善有很大的促進作用[23]。Kabamoto等按照灌溉季節和非灌溉季節在全國10個農業改良項目區的122個地點開展采樣,通過環境DNA技術在農業水道中檢出15~40種魚類,通過比較采樣調查和以往調查數據結果,評估環境DNA技術在快速流動水環境(如農業水道等)中水生態調查的適用性并討論檢測假陽性等技術問題[24]。需要注意的是,其研究強調人類活動影響下環境DNA污染問題,如檢測魚DNA檢測分析中存在實驗室污染等。
聚合酶鏈式反應(PCR)是一種體外核酸擴增技術,用于放大與擴增特定的DNA片段。因此,確定要擴增的DNA片段對后續的eDNA測序分析具有至關重要的作用。在理想情況為了成功地進行eDNA測序分析,PCR引物對和擴增出的中間eDNA片段應滿足以下要求:① 穩定性,設計的通用PCR引物需要無偏差擴增各目標類群的基因片段,且不出現基因片段缺失;② 一致性,擴增的片段應包含足夠的變異以允許明確的分類分配;③ 特異性,引物應是目標類群的特定引物(如MiFish)。否則,來自非目標分類群的DNA(原核生物和非目標真核生物的DNA)將在擴增產物中占主導地位。根據文獻調研對通用PCR引物進行統計發現(見圖3),MiFish引物占比最多(51.2%),其次是Cyt b引物(34.9%),日本常見魚類環境DNA引物(部分)基因序列表如表1所示。這些通用引物,不僅在日本,在我國生物多樣性研究中也開展了相關應用研究[25-26]。從日本環境DNA研究地理分布圖上看,MiFish引物在日本全國的應用案例較多,最近的日本政府調查項目也主要采用MiFish引物開展eDNA分析進行生物多樣性監測[27]。例如:日本國土交通省開展全國河流環境普查試點研究[28];日本環境省生物多樣性中心推動環境DNA標準化項目(https://www.biodic.go.jp/edna/edna_top.html)。而Cyt b引物則是在以瀨戶內海為主的西日本地區應用較為廣泛(見圖4)。
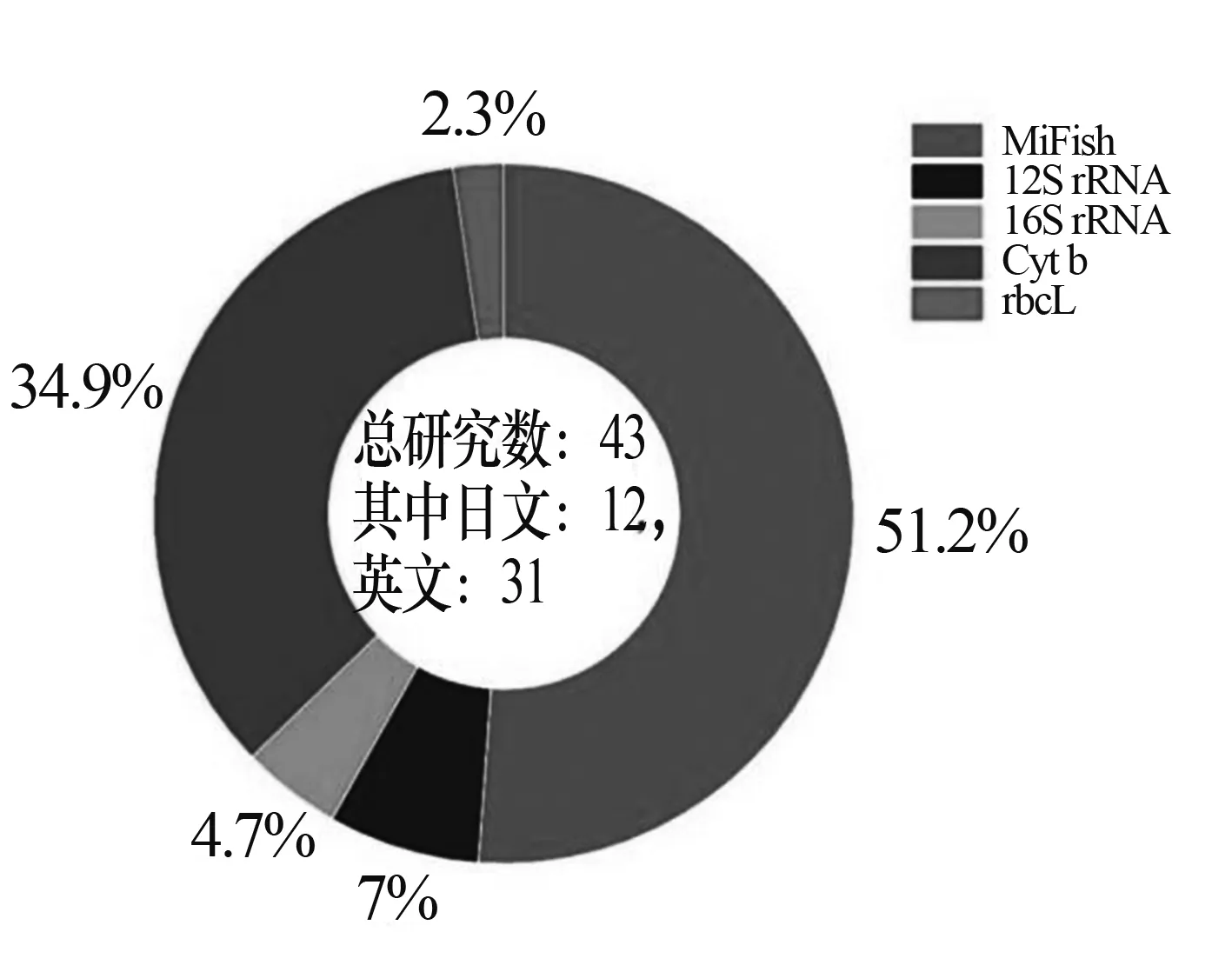
圖3 日本環境DNA發表論文引物頻率組成

圖4 日本環境DNA研究地理分布

表1 日本常見魚類環境DNA引物(部分)基因序列表
3 環境DNA技術在日本河流生物多樣性研究中的應用
隨著環境DNA技術的不斷發展和基因庫的不斷完善,越來越多研究人員運用環境DNA技術開展水生生物的物種鑒定。同時由于外來入侵物種、瀕危物種和稀有物種的監測調查一直是傳統生物監測研究的難題,環境DNA技術為上述問題的解決提供了一種新的思路。本部分針對生物多樣性調查、外來入侵物種的監測、瀕危物種和稀有物種的監測以及生物量評估方面的環境DNA研究及其技術應用進行介紹梳理。
通過Web of Science搜索文獻導入CiteSpace軟件,去重后剩余140篇有效文獻,文獻研究時間段為2005—2022年,本次研究以3 a為時間間隔,選取每年引用頻率前25的關鍵詞進行聚類分析,共得到15類聚類(單一聚類關鍵詞小于5類時不顯示),有效聚類9類(0號 影響因素、1號 浮游植物、2號 日本鰻魚、3 號日本竹莢魚、5號 日本鰻鱺、7號棲息地相關、8號 海洋、9號 大腸桿菌 14號、鑒別),由于當前研究處于發展階段,文獻較少,無法進行關鍵詞突現性研究,本次研究以不同年間關鍵詞共現作為趨勢研究,相關聚類見圖5(a)(2008—2010年)、圖5(b)(2020—2022年),其中圓形大小表示關鍵詞出現次數,同一圓形不同顏色環形為不同年份出現次數,連線表示關鍵詞存在共同出現,粗連線表示a、b特定年間關鍵詞存在共同出現。從圖中可以看出,除環境DNA,qPCR等綱領性關鍵詞外,以生物質、豐度、河流、魚類、數量、瀕危物種、洄游、溫度、降解、琵琶湖等詞出現頻率較高,表明當前環境DNA研究更多關注于物種(多指魚類)豐度、瀕危物種以及季節性物種的監測領域,對于實驗條件的關注則集中在溫度以及引物、環境DNA的降解速度等領域。圖5(a)(2008—2010年)顯示日本早期環境DNA技術主要以基因宏條形碼技術、qPCR擴增為主,并開始應用于琵琶湖流域生物監測。從圖5(b)(2020—2022年)可以看出當前日本環境DNA技術應用開始全面擴寬。在研究區域上,由傳統河湖研究邁向海洋;在技術研究上開始深入引物設計,引物種類得到擴充,對實驗最優條件也進行了一定的探索;在技術應用上從種群監測步向多元研究,在物種多樣性保護、瀕危物種、入侵物種監測、季節性洄游魚類監測等領域均進行了較多研究。關鍵詞聚類表明當前日本對環境DNA技術的研究已進入較深入階段。

圖5 日本環境DNA關鍵詞聚類分析
3.1 生物多樣性調查
生物多樣性的有效保護對于人類的生存和生態系統的維持至關重要。生物多樣性評估基本上依賴于物種監測,但傳統的物種形態學監測難以滿足生物多樣性研究要求。因此,通過從環境樣本中提取DNA來獲取物種、種群和群落信息的方法能夠彌補傳統物種鑒定的不足。在魚類多樣性調查中,環境DNA宏條形碼方法作為一種全面檢測魚類等水生生物鑒定技術被提出。Miya等[12]開發的通用PCR 引物組(MiFsh-U/E)目前已在全球范圍內廣泛使用。該引物是根據880個物種的線粒體完整基因組序列和160個板鰓亞類(鯊魚和鰩魚)物種的部分線粒體序列,針對12S rRNA基因的長可變區(163~185 bp)設計,檢出實驗室180種海洋魚類中的168種(93.3%),并在附近的珊瑚礁調查中也檢測到93種。
在其他水生生物多樣性調查中,Kayano等通過檢測附著板中的環境DNA,表明淡水珍珠牡蠣在河流中的大量存在對水生環境中的附著微動物種群產生影響,并影響河流中氮的遷移轉化[31]。Narumi等人使用線粒體12s rRNA基因靶點的定量PCR技術對沉積物中的水蚤(Daphniagaleata)的DNA濃度進行檢測,并量化上述物種在水中的生物量[32]。Tateishi等通過研究日本絨螯蟹蛻皮對環境DNA釋放的影響,揭示了日本絨螯蟹(Eriocheirjaponica)的分布和季節性變化,發現其幼體在海水中生長,成熟體棲息在淡水區的棲息規律。Aratani等通過開發龍虱科(Dytiscidae)和蝎蝽科(Nepidae)的環境DNA引物集,用于保護列入世界自然遺產的奄美大島等地的水生昆蟲,并評估當地的生物多樣性種群特征[33]。Hirai對太平洋中的海洋浮游生物進行了全面的宏條形碼分析,而這些浮游生物未成熟的個體很難通過形態學來識別,研究重點是占主導地位和種群高度多樣化的尾鰭動物等浮游生物,運用環境DNA宏條形碼技術評估尾鰭動物的多樣性和群落結構[34]。此外,環境DNA技術也被應用于重要漁業物種的飲食分析和浮游生物監測評估食物網結構和生態系統。菅原建立一種專門檢測貽貝的環境DNA方法,并將其應用于湖泊環境的監測調查[35]。池田專注于mtDNA、16s rRNA和12s rRNA區域,并開發相關引物集用于檢測水生昆蟲,并提出黃嘴蜉蝣(Epeorusaesculus)存在未知亞種的可能。Sakata開發并應用了兩棲動物的通用引物[36]。Akatsuka等調查高懸浮物條件下的水樣采集方法,以確定海草床的變化,并在入海口開展海帶鰻鱺(Zosterajaponicaeelgrass)群落及其生物量的數值計算。同時結果表明,受目標習性和繁殖期影響,水樣采集的適應時間因地而異,對生物多樣性實地調查具有重要作用[37]。Yonezawa等通過水中的環境DNA揭示了棲息在水邊并以水中的魚和水生昆蟲為食的亞洲水鼩(Chimarrogaleplatycephalus)的棲息地分布[38]。 此外,Ishige等分析馬來西亞婆羅洲鹽池水樣的環境DNA,發現猩猩(Pongopygmaeus)、爪哇野牛(Bosjavanicuslowi)、亞洲象(Elephasmaximus)等多種動物,均顯示了環境DNA同傳統形態學觀察監測相比的優越性[39]。Ushio等使用新開發的MiBird引物在從動物園鳥籠的飲水區采集的樣本檢測出鳥類的環境DNA[13]。
3.2 外來入侵物種的監測
外來物種入侵是指外來種由原生地經自然或人為途徑進入另一個生態環境,并在該生態環境系統中定居、自行繁殖建群和擴散而逐漸占領新棲息地的一種生態現象[40-42]。環境DNA技術可以促進對水生生物入侵物種的檢測和監測,促進入侵物種的監測和預防工作開展。Takahara等利用環境DNA技術對入侵的藍鰓太陽魚(Lepomismacrochirus)環境DNA進行了檢測,發現藍鰓太陽魚入侵情況在人類活動的影響較少的島嶼池塘中較輕,具有比傳統觀察方法更好的生物入侵檢測率[43]。Maruyama等發現入侵物種藍鰓太陽魚中較小的個體每單位體重釋放更多的DNA到水中,從而影響環境DNA濃度估算生物量和種群密度的結果[44]。此外,Uchii等研究結果表明,根據線粒體DNA的單核苷酸差異可以區分同一物種內非本地基因轉移,并量化鯉魚中本地和非本地DNA的相對比例,進而開發量化基于單核苷酸差異的本地和非本地DNA的相對比例的檢測方法[44]。Mizumoto等人通過環境DNA手段發現日本蟾蜍(Bufojaponicusformosusk)的繁殖區廣泛分布在北海道的12條河流的32個地點,由于其毒性而影響本地物種的生存,但其夜間的性質和隱藏習慣導致其很難被發現和消除[45]。同時Jo等人對多個池塘樣本采用多重實時PCR方法,發現不同季節的3種非本地魚種(Lepomismacrochirus;藍鰓魚,Micropterussalmoides;大嘴鱸和Micropterusdolomieu:大口黑鱸)和3種瀕臨滅絕的本地魚類(Hemigrammocyprisrasborella:間紋鯉;Oryziaslatipes:日本青鳉;Misgurnusanguillicaudatus:泥鰍)在不同季節檢出率不同。對其分布情況進行評估,總體結果顯示夏季的環境DNA檢測率較高,并發現非本地物種在人類活動影響大、池塘表面積大、pH值高的池塘中更容易被檢出,而本地物種在非本地物種檢測率高的池塘中檢出率較低[47]。Fkumoto等使用環境DNA來確定大鯢(Andriasjaponicus)和中國大鯢(Andriasdavidianus)的分布,這是一個通過入侵并與本地物種雜交影響本地物種棲息地的入侵物種,根據其活動周期等確定相應的調查時期[48]。Yamakawa和Miya修改MiFish 引物的部分序列,用于檢測沼澤中的非本地木龜(Chelydraserpentina),并正在開發MiTurtle引物集,其序列對淡水龜類具有高度特異性。在研究區域除了非本地木龜(C.serpentina),還發現了日本石龜(Pelodiscussinensisver.japonica)、棱皮龜(Mauremysreevesii)、日本石龜(Mauremysjaponica)、非本地物種包括紅耳龜(Trachemysscripta)、箱龜(CuoragalbinifronsMoegi,居住在中國南部)、鱷龜(Macrochelystemminckii)、馬蹄龜(Sternotheruscarinatus)等[49]。針對入侵物種柳杉(Limnopernafortunei)在許多地方造成供水設施的阻塞問題,Ito等建立其環境DNA調查方法,并且正在Kasumigaura(霞浦)水系統中進行入侵監測。同時在5個水庫對入侵物種柳杉進行常規調查方法(目測、采樣和幼蟲觀察)和環境DNA調查。在所有用常規方法檢測到柳杉的水庫以及用常規方法未檢測到的水庫中,都檢測到柳杉DNA。基于種群密度和環境DNA濃度之間的關系,在實驗水庫中也揭示了定量趨勢[50]。池田對日本特有物種日本小龍蝦(Cambaroidesjaponicus)和可能影響其生境的入侵物種內田小龍蝦(Pacifastacusleniusculus)在北海道的分布進行了調查[36],環境DNA調查在檢測這兩個物種方面有優勢,因為它們隱藏在礫石下,所以很難觀察和捕獲。這項研究的結果顯示,內田小龍蝦可能入侵上游地區,影響本地物種。
3.3 瀕危物種和稀有物種的監測
由于瀕危物種和稀有物種生物量較少,傳統的調查方法存在精確度偏低與破壞棲息生存環境等問題,僅靠傳統的捕捉和觀察調查很難進行評估,而環境DNA分析技術能有效規避這些問題。Sakata等[51]在Omonogawa(大野川)河的99個水樣中的2個檢測到細鱗鱊(Acheilognathustypus),并在檢出水區開展捕捉調查,確認其中一個地點存在成熟的細鱗鱊雄性和雌性以及作為產卵宿主的雙殼類。這一發現是該河流11 a來的首次發現,有效證明環境DNA調查是確定大河中稀有物種分布和繁殖區域的有用工具。在這些地方Akamatsu等利用線粒體的DNA區開發一套引物探針,并在野外應用,發現檢測到的DNA片段與潛水目測觀察到的個體數量呈正相關,表明環境DNA可用于以符合國家規定的方式進行調查[52]。
3.4 生物量及生態影響評估
環境DNA技術有助于魚類生物量或豐度的非侵入性量化,但目前DNA采集方法和環境條件對水樣中環境DNA濃度的影響并沒有系統的標準和了解,這樣對運用環境DNA技術量化評估魚類生物量造成限制。Doi等人在2016年利用潛水觀察和攝食痕跡等方法調查河流中的常駐香魚,對香魚(Plecoglossusaltivelis)進行定量分布調查和環境DNA研究,兩者之間存在明顯的關系,證明環境DNA是一個來估計河水中香魚的豐度與生物量、分布的有效方法[10]。日本山口大學的研究小組利用環境DNA來評估大壩建設對流域生境的影響,并驗證改進項目的效果。研究表明,隨著支流流量的增加,香魚棲息地的密度在夏季也會增加,并建議在支流中建設適合香魚棲息的主要區域[53-54]。Hanaoka等使用環境DNA來驗證為恢復江之川濱原大壩下游的河流環境而實施的沉積物供應項目的效果,發現由于土壤徑流的改善作用,沙質底部魚類生物量有所增加[55]。此外,利用環境DNA技術揭示繁殖季節環境DNA濃度的晝夜變化以及繁殖行為中環境DNA顆粒大小特征并通過核DNA與線粒體DNA的比率評估產卵行為,根據河水溫度和環境DNA濃度關系的實地研究數據,利用氣候變化導致的河水溫度上升模型,預測未來香魚棲息地的減少。根據香魚幼崽釋放的DNA數量及河水環境DNA濃度調查幼崽的實際數量,并評估河口堰塞湖流速降低對其的影響。洪水事件中,香魚生境密度和生境使用的變化研究中發現,香魚在洪水來臨之前與護岸比率呈負相關,與流速呈正相關;而在洪水來臨之后,只有護岸比率呈負相關,表明香魚生境受到洪水干擾導致的支流規模的影響[56]。Imashiro等同時檢查Shimanto(四萬十市)河中的香魚和Flavobacteriumpsychrophilum(一種引起香魚冷水病的革蘭氏陰性長桿菌)的環境DNA,發現冷水病不是發生在5月(即原來的高峰期),而是在11月和12月發生在河口附近的產卵地[57],同時研究顯示,產卵和攜帶真菌的香魚的存在是感染擴散的驅動力。Ushio等通過使用illumina MiSeq并添加內部DNA標準,成功獲得了沿海地區70多種魚類的定量環境DNA時間序列數據。基于這些數據,Kondo對魚類之間以及魚類與水母之間的生態關系進行了時間序列分析,并與物理觀測數據相結合,建立了預測海灣中竹莢魚(Trachurusjaponicus)表層、中層和底部豐度的模型[58]。
3.5 環境DNA評估河道設施對魚類影響
當前使用環境DNA技術評估河道設施對魚類影響的研究較少。Aranishi等對關東河干流Shidzumi水庫附近60 km長度的香魚洄游河段進行了其縱向分布的環境DNA調查,發現在水庫上游13 km及水庫內廣泛檢測到香魚DNA,而在下游未檢測到其DNA,驗證了大壩對季節性洄游魚類的阻礙作用[59]。Yamanaka等通過對3個目標洄游魚種(溫帶鱸魚、平頭鯔魚、鯔魚)的DNA片段的物種特異性檢測,證明了目標物種的eDNA的存在與否與已知的物種季節性遷移模式一致,且在配置魚梯的大壩上游最靠近河口的地方檢測到溫帶鱸魚和平頭鯔魚的DNA,而在另外兩個未配置魚梯的大壩上游未檢測到上述兩者的DNA,表明魚梯能幫助這些物種通過繞過大壩的魚梯成功向上游遷移[60]。Morita等利用電捕魚與環境DNA結合的方法研究了日本北海道島西南部大島渚半島30條有水壩的溪流中白斑紅點鮭魚(Salvelinusleucomaenis)種群滅絕的脆弱性,比較了1999年至2014年間溪流中白點鮭種群的滅絕概率,發現其局部滅絕的概率與流域面積成反比,而安裝魚梯重新連接隔斷的生境后,能有效緩解白斑紅點鮭魚的局部滅絕危機[61]。
4 總結與展望
利用環境DNA了解生物多樣性需要對多物種進行宏條形碼的全面檢測并結合特定物種的檢測來確定物種的存在與否和它們的生物量。日本環境DNA協會已經出版了一本全面總結環境DNA作為一種調查和分析技術的手冊《環境DNA調查和實驗手冊》,同時日本環境省自然保護局生物多樣性中心也出版了《使用環境DNA分析技術的淡水魚類調查方法手冊》等技術資料。
環境DNA技術最嚴重的缺陷是存在污染的風險,即可能出現假陽性的結果。環境DNA在樣品的采集、運輸、保存等過程中都容易發生交叉污染,在實驗室分析過程中亦會受到試劑的污染。污染問題是造成環境DNA實驗結果誤差的一個重要影響因素。可能導致分析錯誤的因素包括假陰性的問題,環境DNA技術需要依靠數據庫,但是現如今的數據庫生物信息并不完善,特別是針對感興趣的生物體的生態環境(如低活性或者有限的時間內活動),缺乏參考物種名稱的序列信息,以及良好的檢測系統的發展不足影響環境 DNA 的最終檢測結果。此外,環境DNA分析方法需要進一步規范完善,很多研究人員在環境DNA樣品采集、保存、提取等過程采取的方法各不相同,并沒有形成系統的分析方法,今后的研究中可以對環境DNA技術操作流程進行進一步的優化和完善。環境中的DNA會被微生物和紫外線輻射所降解,所以目前很難用具體的數值來預測和評估其降解率、平流和擴散。環境DNA技術的檢測結果在時間和空間這兩個維度的精度較低,檢測精度有待提升。環境DNA技術可以通過特定物種DNA的有無直觀的表征河道建筑對魚類活動的影響,可以通過檢測洄游性魚類的DNA來評估河道設施對河流生態健康的影響,但由于當前環境DNA技術仍處于發展階段,在定量評價上需要未來的進一步研究。
每一種調研方法都有其優點和缺點,因此了解和使用環境DNA作為生物多樣性研究的工具的優缺點是非常重要的。環境DNA可以大大增加在時間和空間上獲得生物多樣性數據的可能性,它可以用來了解生物體與環境中各種因素之間的相互作用。通過進一步優化環境DNA樣品采集、保存方法、DNA提取等分析方法,預計環境 DNA技術未來將進一步發展跨學科的綜合研究,以解讀生物體與其環境中各種因素之間的相互作用,例如其將食物鏈的能量流動、動物攝食等研究領域的作用展示出來。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
航空世界(2020年10期)2020-01-19 14:36:20
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
