餌料、鹽度和溫度對刺巨藤壺幼蟲生長發育的影響
2022-11-26 08:17:50饒小珍
生態學報 2022年21期
林 崗,饒小珍,*
1 福建師范大學生命科學學院,福州 350117
2 福建師范大學福建省特色海洋生物資源可持續利用重點實驗室,福州 350117
無柄蔓足類 (藤壺) 是沿海生態系統的重要組成部分,也是主要的污損生物之一[1—3],常成群固著于海岸巖石、堤壩碼頭、艦船浮標、海水管道、平臺設施及養殖設備上,對人類涉海經濟活動產生極大危害和巨大經濟損失。國外對紋藤壺屬 (Amphibalanus) 的紋藤壺 (A.amphitrite)、網紋紋藤壺 (A.reticulatus) 和象牙紋藤壺 (A.eburneus)和藤壺屬 (Balanus) 的三角藤壺 (B.trigonus)等種類的幼蟲形態、各種生態因子對幼蟲發育的影響、特別是以紋藤壺為模式動物對金星幼蟲附著和變態的條件和機制等進行了大量研究,這些工作為蔓足類的生物學、生態學和防除研究奠定了堅實基礎[2—10]。國內中國科學院南海海洋研究所曹文浩等主要以網紋紋藤壺為對象也進行了一些相關報道[11—14]。
刺巨藤壺 (Megabalanusvolcano) 屬于節肢動物門 (Arthropoda)、甲殼亞門 (Crustacea)、蔓足下綱 (Cirripedia)、圍胸總目 (Thoracica)[15],棲息于熱帶和亞熱帶海域低潮帶或潮下帶,我國的東海和浙江舟山以南至西沙群島海域是其主要分布地。刺巨藤壺雖然是沿海污損生物的優勢種,但它的肌肉非常發達,也是沿岸居民常食用的海鮮之一[1,14]。巨藤壺屬的紅巨藤壺(M.rosa)在藤壺膠的研究中起重要作用[16—17];而另一種巨藤壺M.azoricus個體很大,閉殼肌和卵巢發達,在葡萄牙亞速爾群島作為一個經濟種類受到過度采集,資源量不斷下降[18—19],目前也嘗試開展人工增養殖的研究[18—20]。蔓足類的生活史通常由卵、6期無節幼蟲、1期不攝食的金星幼蟲和固著的成體組成[1—3]。幼蟲培養的成功與否是研究藤壺幼蟲發育、揭示其附著機理的基礎,也是開展防污材料篩選的前提條件[1—2]。本課題組對刺巨藤壺的精子結構進行了觀察[21],但有關刺巨藤壺幼蟲室內培養條件的研究未見報道。培養刺巨藤壺幼蟲適宜的餌料是什么?幼蟲生長發育適宜的溫度和鹽度范圍是多少?了解這些基本生態因子對幼蟲的影響可以明確刺巨藤壺幼蟲培養的適宜條件。本文以刺巨藤壺Ⅵ期無節幼蟲存活率、Ⅵ期無節幼蟲出現時間、金星幼蟲存活率、金星幼蟲出現時間和金星幼蟲體長等為觀察指標,研究了3種餌料及密度(中肋骨條藻Skeletonemacostatum、牟氏角毛藻Chaetocerosmuelleri和亞心型扁藻Platymonassubcordiformis)、鹽度和溫度等因子對刺巨藤壺幼蟲生長發育的影響,以便能在實驗室培養刺巨藤壺金星幼蟲,為藤壺生物學、海洋生態學及附著機理等方面的研究提供資料,也可為其人工增養殖的開展提供指導。
1 材料與方法
1.1 實驗材料
1.1.1 刺巨藤壺成體室內培養與幼蟲的采集
刺巨藤壺成體于2017年 5月下旬采自福建省平潭綜合實驗區蘆洋鄉馬腿村 (26.0173°N,119.6653°E),海區水溫24.1℃、鹽度28.8,成體峰吻徑20—27 mm、高43—62 mm。親體在實驗室放置于200 L的養殖桶中培養育肥,每日投喂藻類和鹵蟲,并定時清污。養殖條件為海水鹽度29—30、pH為7.9—8.0、水溫25—27℃,所用海水經過濾、紫外線消毒處理。每日檢查,觀察到刺巨藤壺成體釋放出Ⅰ期無節幼體,以篩網立即收集Ⅰ期無節幼體,將收集到的Ⅰ期無節幼蟲置于容量為 3000 mL 的玻璃燒杯中。Ⅰ期無節幼體在12 h內將發育為Ⅱ期無節幼體,利用幼蟲的趨光性用吸管吸取健康活潑的Ⅱ期無節幼蟲作為實驗材料。
1.1.2 藻類的培養
實驗所需中肋骨條藻、牟氏角毛藻和亞心型扁藻由福建師范大學藻種培養室提供。用f/2培養基進行藻類單種擴大培養。以對數生長期的藻類進行投喂。藻類培養溫度在(24±1)℃,光照周期為D:L=18h:6h。
1.2 實驗設置
實驗在500 mL的燒杯中進行,400 mL海水引入100只刺巨藤壺無節幼蟲,幼蟲密度為0.25只/mL,實驗過程中不充氣。投餌時,先用血球計數板確定藻類原液濃度,再經換算,在每個實驗組中投入相應的餌料量。
每天全換水1次,投餌1次。在換水之前隨機用吸管吸取約10只幼蟲在顯微鏡下觀察和記錄其生長狀況。換水時利用幼蟲的趨光性吸取培養液中存活的幼蟲到新的燒杯。一旦觀察到出現Ⅵ期無節幼蟲和金星幼蟲時,統計燒杯中存活的無節幼蟲和金星幼蟲數量。直到每個燒杯中的無節幼蟲發育為金星幼蟲或者全部無節幼蟲死亡時實驗結束。
1.2.1 餌料實驗
中肋骨條藻設置5×104個/mL、10×104個/mL、20×104個/mL、40×104個/mL和80×104個/mL 5個密度梯度,牟氏角毛藻設置5×104個/mL、10×104個/mL、20×104個/mL、40×104個/mL和80×104個/mL 5個密度梯度,亞心型扁藻設置2.5×104個/mL、5×104個/mL、10×104個/mL、15×104個/mL和20×104個/mL 5個密度梯度。每個實驗組三個平行。海水鹽度為29—30、水溫25—26℃。
1.2.2 鹽度實驗
設置7個鹽度梯度分別為18、22、26、30、34、38和42。高鹽組用海水晶、低鹽組用反滲水進行配制。所用餌料為牟氏角毛藻,藻類密度控制在50×104個/mL,水溫25—26℃。每個實驗組三個平行。
1.2.3 溫度實驗
設置6個溫度梯度分別為14℃、18℃、22℃、26℃、30℃和34℃。各溫度組用控溫系統進行調控。所用餌料為牟氏角毛藻,藻類密度控制在50×104個/mL,海水鹽度為29—30。換水前各溫度組使用的海水提前進行預溫處理。每個實驗組三個平行。
1.3 實驗觀察和統計的指標
1.3.1 Ⅵ期無節幼蟲的發育時間和存活率
Ⅵ期無節幼蟲的發育時間為每實驗組最早出現Ⅵ期無節幼蟲時發育所用的時間(d)。
Ⅵ期無節幼蟲存活率=出現Ⅵ期無節幼蟲時統計存活的無節幼蟲數量/起始的無節幼蟲數量×100%。
1.3.2 金星幼蟲的發育時間和存活率
金星幼蟲的發育時間為每實驗組最早出現金星幼蟲時發育所用的時間(d)。
金星幼蟲存活率=實驗結束時,每實驗組中累計獲得的金星幼蟲總數/起始的無節幼蟲數量×100%。
1.3.3 金星幼蟲體長的測量
用目鏡測微尺測量金星幼蟲的體長。將同一實驗條件培養的金星幼蟲收集到同一燒杯中,若金星幼蟲超過20只,隨機抽取20只測量;若不足20只金星幼蟲,對所有蟲體進行測量。
1.4 數據分析
2 結果
2.1 餌料對刺巨藤壺幼蟲發育的影響
2.1.1 中肋骨條藻對刺巨藤壺幼蟲發育的影響
中肋骨條藻能夠培養刺巨藤壺無節幼蟲至金星幼蟲階段(圖1)。5×104個/mL密度組中的幼蟲只能發育到Ⅵ期無節幼蟲階段,無法發育到金星幼蟲,其Ⅵ期無節幼蟲的存活率僅6.00%;其它各組均高達60.00%以上。金星幼蟲的存活率在10×104個/mL組僅2.33%,而80×104個/mL組的高達41.33%。經過單因素方差分析表明,骨條藻密度對刺巨藤壺Ⅵ期無節幼蟲存活率(F=33.835,P=0.000)和金星幼蟲存活率(F=14.833,P=0.001)有極顯著影響。經過多重比較,Ⅵ期無節幼蟲的存活率除5×104個/mL組外其它各組間未表現出明顯差異(P≥ 0.420)。金星幼蟲存活率在10×104個/mL組和20×104個/mL組(P=0.165)、20×104個/mL組和40×104個/mL組(P=0.154)、40×104個/mL和80×104個/mL組(P=0.513)之間的存活率無顯著差異。
隨著骨條藻密度的增加,Ⅵ期無節幼蟲和金星幼蟲發育時間漸縮短(圖2)。5×104個/mL組的Ⅵ期無節幼蟲發育最慢長達11 d,40×104個/mL和 80×104個/mL密度組的幼蟲發育最快只需4 d。到達金星幼蟲階段的時間,10×104個/mL組和20×104個/mL密度組發育較慢分別需12 d和11 d;40×104個/mL組和80×104個/mL密度組發育較快,分別需7 d和6 d。
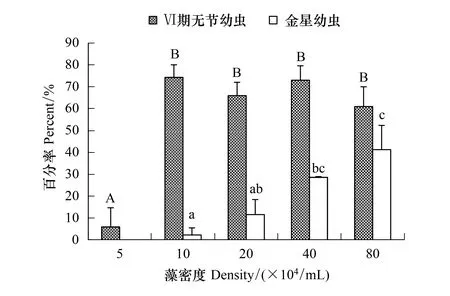
圖1 中肋骨條藻密度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲存活率的影響
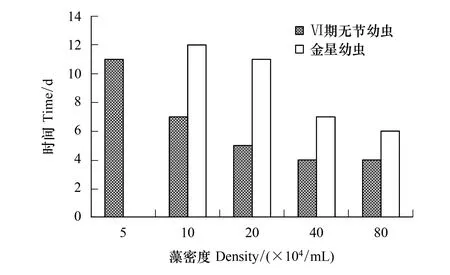
圖2 中肋骨條藻密度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲發育時間的影響
在不同骨條藻密度培養的金星幼蟲體長見表1。低密度10×104個/mL組的金星幼蟲體長最短,高密度80×104個/mL組的最長。總的來看,隨著骨條藻密度的增大,金星幼蟲的體長增加。經單因素方差分析表明藻密度對金星幼蟲體長有極顯著影響(F=39.387,P=0.000)。經過多重比較,各密度組間金星幼蟲體長均有極顯著差異 (P<0.01)。
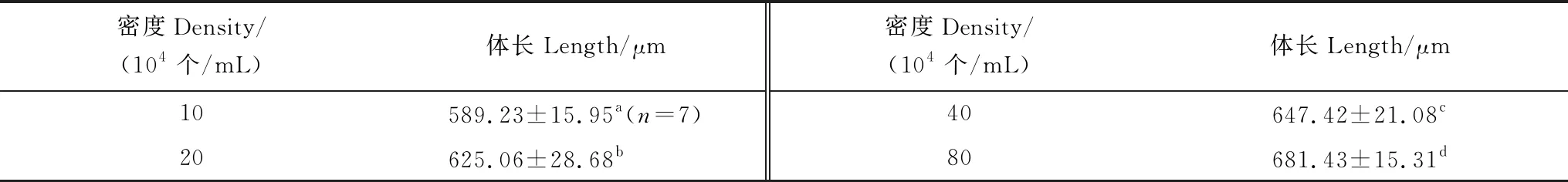
表1 中肋骨條藻密度對刺巨藤壺金星幼蟲體長的影響
總之,綜合幾個觀察指標,中肋骨條藻培養刺巨藤壺幼蟲的適宜密度是40×104個/mL和 80×104個/mL。
綠色礦山評價指標眾多,包括定性和定量指標。本文采用層次分析法(AHP)與模糊綜合評價法相結合方式,將定量指標與定性指標一起進行多級模糊綜合評價。
2.1.2 牟氏角毛藻對刺巨藤壺幼蟲發育的影響
牟氏角毛藻能夠培養刺巨藤壺無節幼蟲至金星幼蟲階段(圖3)。5×104個/mL 密度組中的幼蟲只能發育到Ⅵ期無節幼蟲階段,無法發育到金星幼蟲階段。Ⅵ期無節幼蟲的存活率5×104個/mL組僅15.00%,而80×104個/mL組高達74.67%。金星幼蟲的存活率10×104個/mL組僅5.33%,而80×104個/mL組高達43.67%。經過單因素方差分析表明,角毛藻密度對刺巨藤壺Ⅵ期無節幼蟲存活率(F=95.439,P=0.000)和金星幼蟲存活率(F=49.634,P=0.000)有極顯著影響。經過多重比較,Ⅵ期無節幼蟲存活率 5×104個/mL組和10×104個/mL組與各組間均有極顯著差異(P<0.01),20×104個/mL組和40×104個/mL組(P=0.188)、40×104個/mL組和80×104個/mL組之間的存活率無顯著差別(P=0.852)。金星幼蟲的存活率在10×104個/mL組和20×104個/mL組與其它組間均有極顯著差異(P< 0.01),而40×104個/mL和80×104個/mL密度組之間的存活率無顯著差異(P=0.419)。
隨著角毛藻密度的增加,Ⅵ期無節幼蟲和金星幼蟲發育時間漸縮短(圖 4)。5×104個/mL組的Ⅵ期無節幼蟲發育時間長達10 d,而40×104個/mL組和80×104個/mL組發育最快只需5 d。金星幼蟲發育時間10×104個/mL組的長達12 d,而80×104個/mL組的發育最快只需7 d。
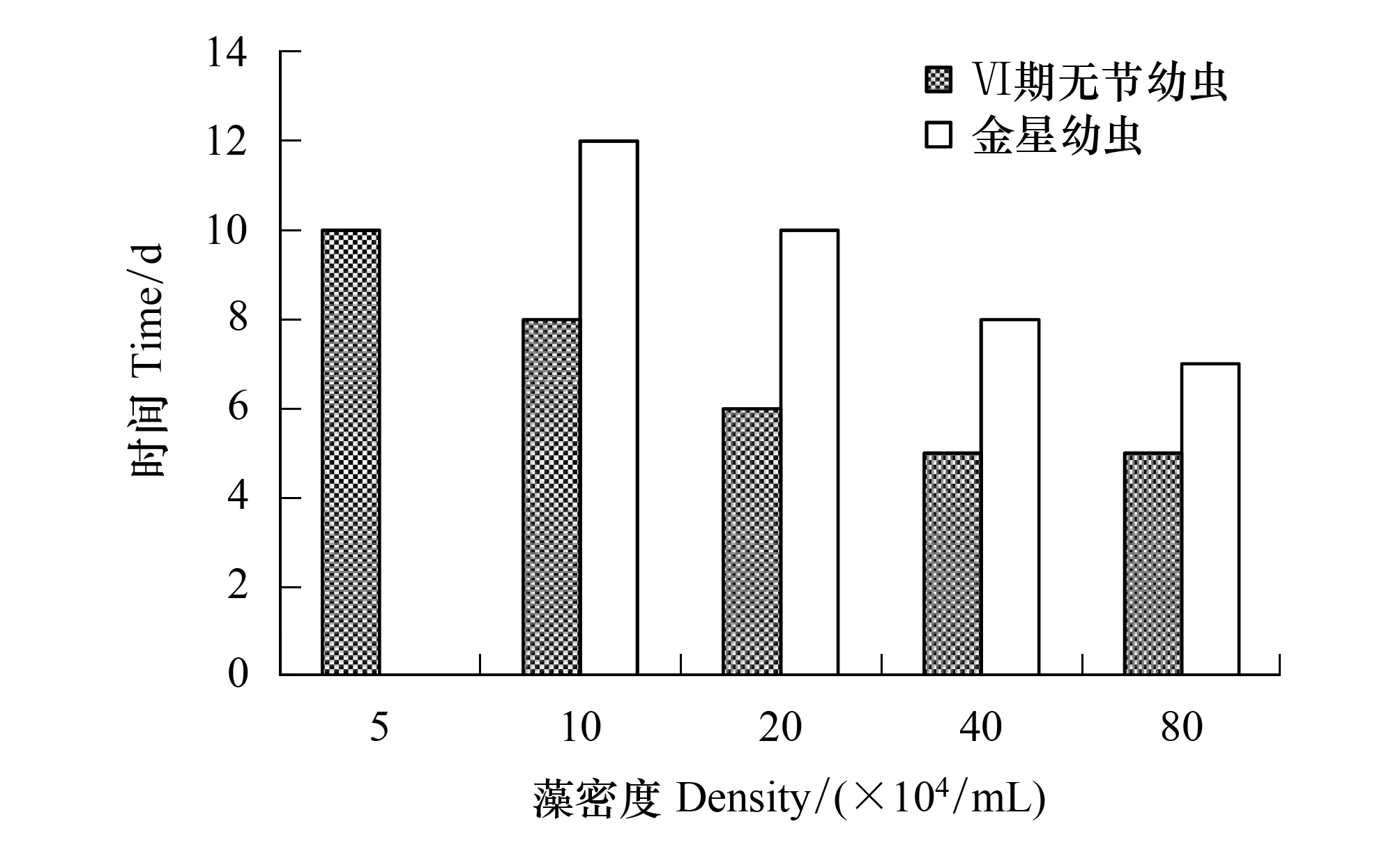
圖4 牟氏角毛藻密度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲發育時間的影響
牟氏角毛藻密度對刺巨藤壺金星幼蟲體長的影響見表2。低密度10×104個/mL組的金星幼蟲體長最短,高密度80×104個/mL組的最長。總之,隨著角毛藻密度的增大,金星幼蟲的體長增加。經單因素方差分析表明角毛藻密度對金星幼蟲體長有極顯著影響(F=26.613,P=0.000)。經過多重比較表明,10×104個/mL組和20×104個/mL組的金星幼蟲體長與其它各組間均有極顯著差異 (P< 0.01),而40×104個/mL組和80×104個/mL組間的金星幼蟲體長沒有顯著差異。
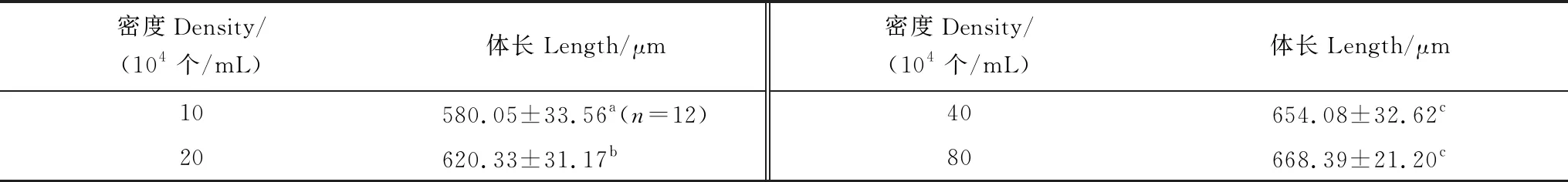
表2 牟氏角毛藻密度對刺巨藤壺金星幼蟲體長的影響
總之,綜合幾個觀察指標,牟氏角毛藻培養刺巨藤壺幼蟲的適宜密度是40×104個/mL和80×104個/mL。
2.1.3 亞心形扁藻對刺巨藤壺幼蟲發育的影響
亞心形扁藻能夠培養刺巨藤壺無節幼蟲至金星幼蟲階段(圖5)。2.5×104個/mL和5×104個/mL低密度組的幼蟲只能發育到Ⅵ期無節幼蟲階段;其它3個高密度組的幼蟲可以發育至金星幼蟲階段。Ⅵ期無節幼蟲的存活率在15×104個/mL組高達84.33%。但各密度組Ⅵ期無節幼蟲變態至金星幼蟲的存活率顯著下降;金星幼蟲存活率最高為20×104個/mL組僅14.33%。經單因素方差分析表明,扁藻密度對刺巨藤壺Ⅵ期無節幼蟲(F=32.035,P=0.000)和金星幼蟲的存活率(F=14.347,P=0.005)有極顯著影響。經過多重比較,2.5×104個/mL組和5×104個/mL組間的Ⅵ期無節幼蟲存活率未表現出明顯差異(P=0.147),10×104個/mL組、15×104個/mL組和20×104個/mL組(P≥ 0.329)三組間也沒有顯著差異。10×104個/mL組的金星幼蟲存活率與其它兩個密度組間有顯著差異,15×104個/mL組與20×104個/mL組間的存活率無顯著差異(P=0.173)。
隨著扁藻密度的增加,Ⅵ期無節幼蟲和金星幼蟲的發育時間漸縮短(圖6)。2.5×104個/mL組的Ⅵ期無節幼蟲發育時間長達9 d,15×104個/mL組和 20×104個/mL組發育最快只需6 d。金星幼蟲發育時間10×104個/mL組的較長需10 d,20×104個/mL組的最短僅8 d。
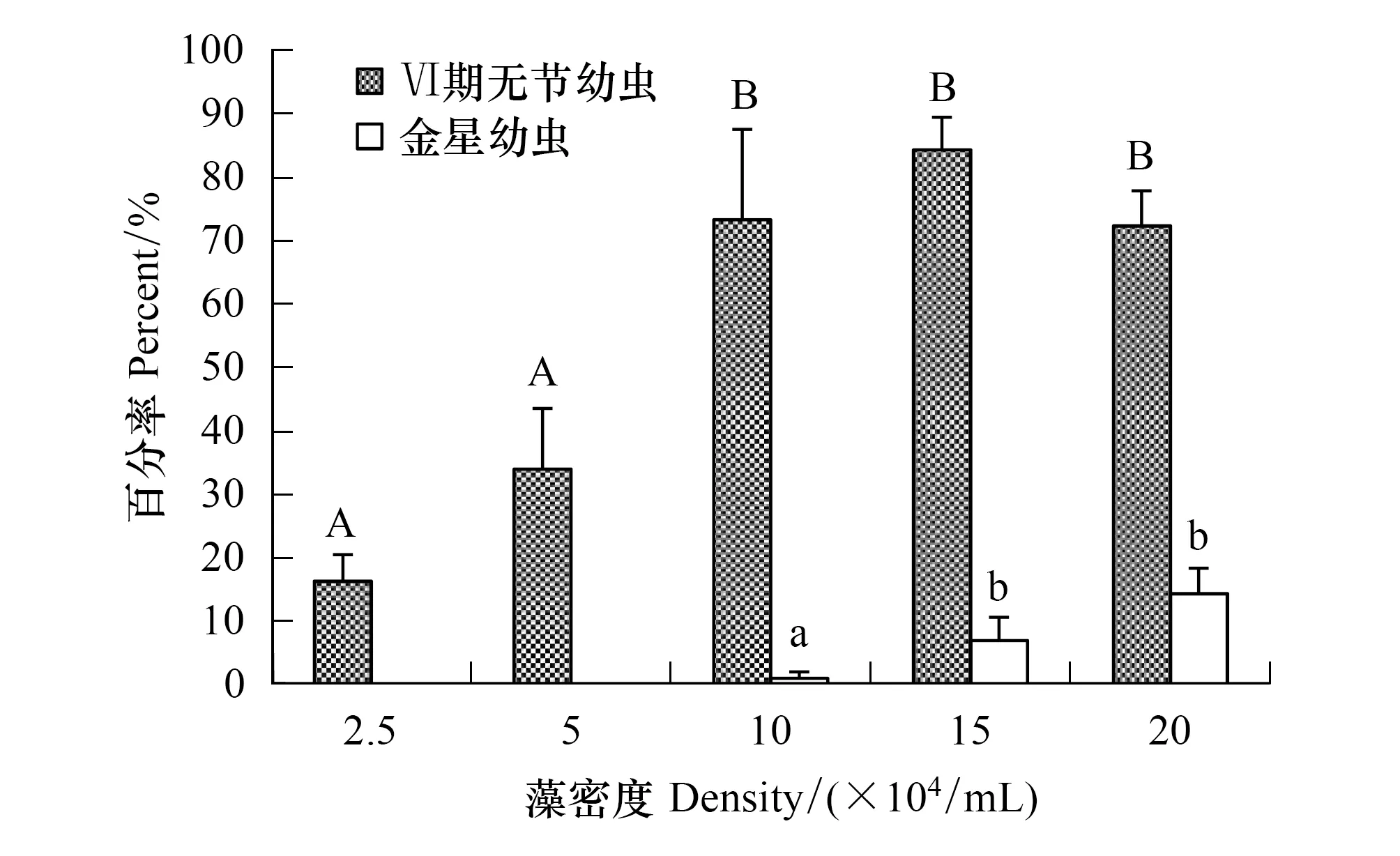
圖5 亞心形扁藻密度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲存活率的影響
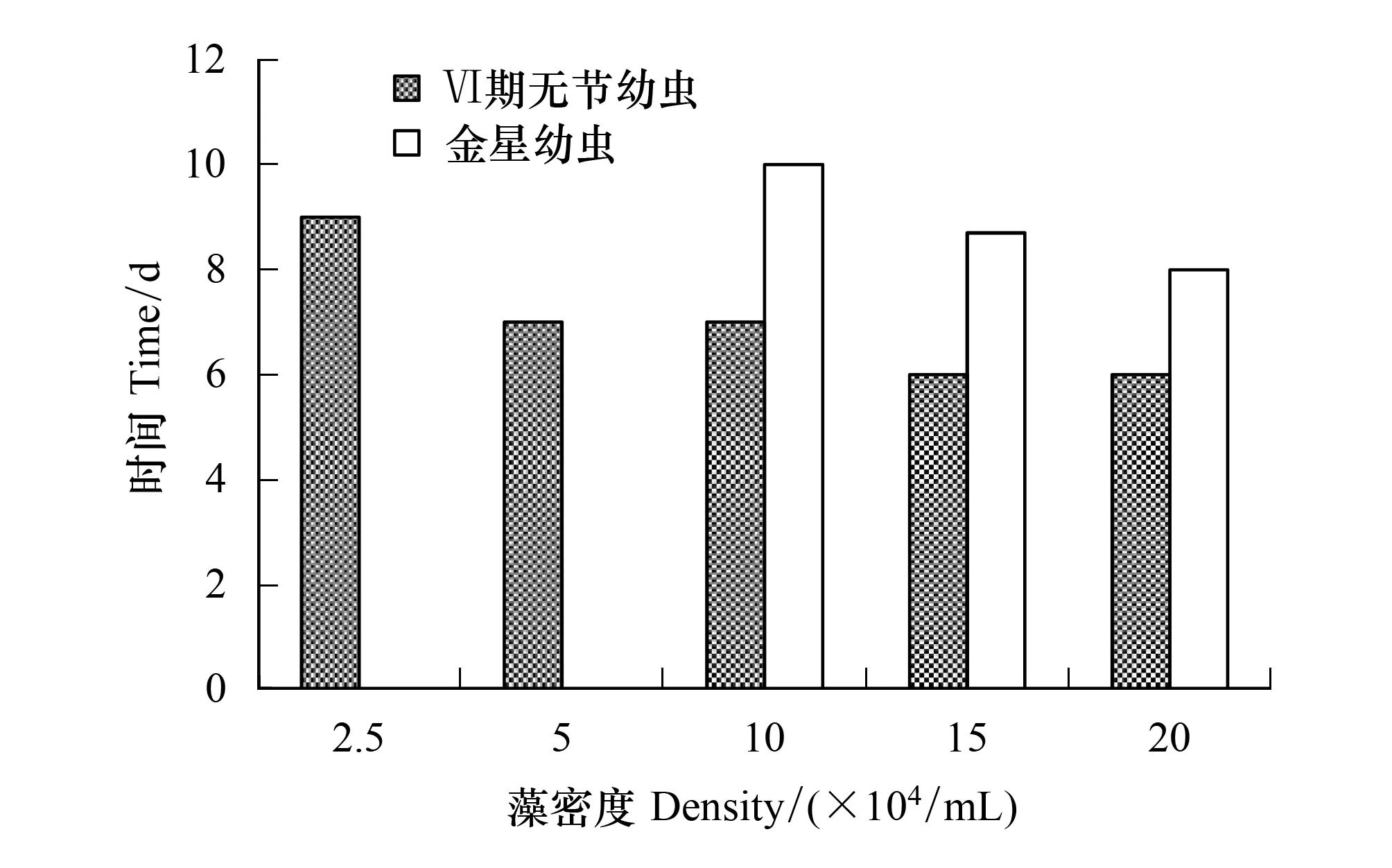
圖6 亞心形扁藻密度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲發育時間的影響
扁藻密度對刺巨藤壺金星幼蟲體長的影響見表3。經單因素方差分析表明扁藻密度對金星幼蟲體長沒有影響(P>0.05),各組間的體長沒有明顯差異。總體而言,用扁藻培養的刺巨藤壺金星幼蟲個體較小、存活率低,亞心形扁藻不是培養刺巨藤壺幼蟲的適宜餌料。

表3 亞心形扁藻密度對刺巨藤壺金星幼蟲體長的影響
2.2 鹽度對刺巨藤壺幼蟲發育的影響
刺巨藤壺幼蟲除了18鹽度組只能發育到Ⅳ期無節幼蟲外,從22至42各鹽度組均能發育到金星幼蟲階段(圖7)。Ⅵ期無節幼蟲存活率以34鹽度組最高為70.33%,22鹽度組最低為30.67%。金星幼蟲存活率以30鹽度組最高為35.67%,42鹽度組最低為9.67%。經過單因素方差分析表明,鹽度對Ⅵ期無節幼蟲存活率(F=76.143,P=0.000)和金星幼蟲存活率(F=7.022,P=0.003)均有極顯著影響。經過多重比較,22和42鹽度組的Ⅵ期無節幼蟲存活率與其它各組間均有顯著差異(P< 0.05)。22、38和42鹽度組間的金星幼蟲存活率沒有顯著差異(P≥ 0.268),22、26、30、34和38鹽度組之間的也無顯著差異(P≥ 0.129),但42鹽度組與26、30、34鹽度組間有極顯著差異(P≤ 0.01)。
鹽度對Ⅵ期無節幼蟲和金星幼蟲發育時間的影響不明顯(圖8)。各鹽度組Ⅵ期無節幼蟲發育時間均為6 d,金星幼蟲的發育時間在8 d和8.7 d之間。
鹽度對刺巨藤壺金星幼蟲體長的影響見表4。22鹽度組的金星幼蟲體長最長,38鹽度組的最短。經方差分析表明鹽度對金星幼蟲體長有顯著影響(F=17.994,P=0.000)。經過多重比較,22和26鹽度組的金星幼蟲體長沒有顯著差異 (P=0.392),30、34、38、42鹽度組間的體長也沒有顯著差異(P≥ 0.199)。低鹽培養下,金星幼蟲的體長相對較長。
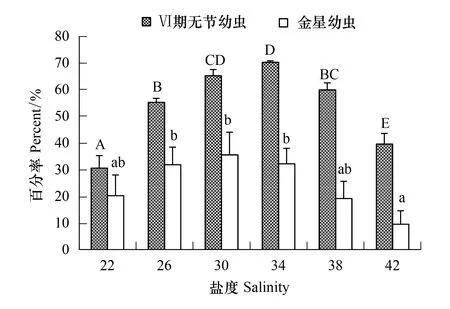
圖7 鹽度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲存活率的影響
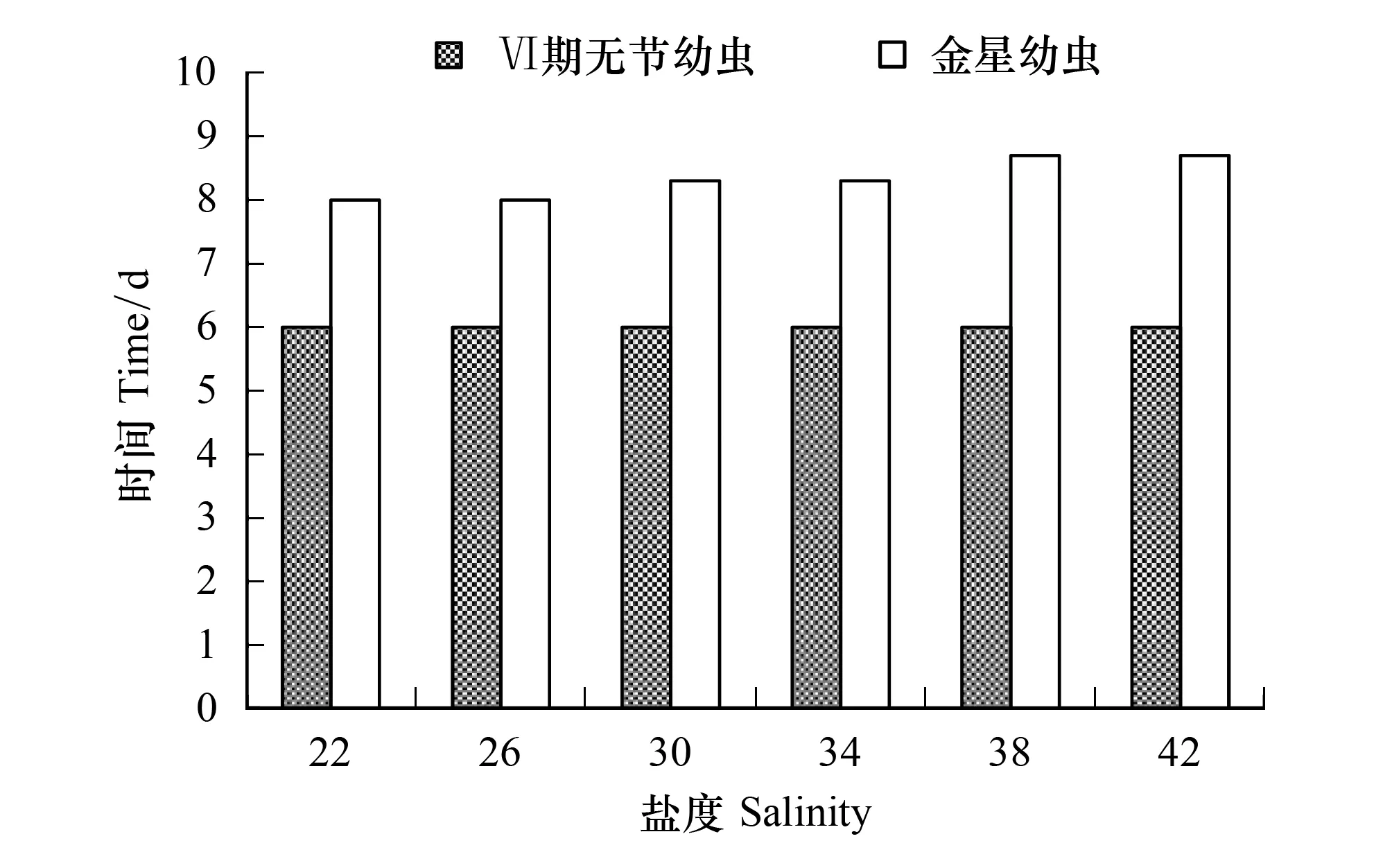
圖8 鹽度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲發育時間的影響

表4 鹽度對刺巨藤壺金星幼蟲體長的影響
2.3 溫度對刺巨藤壺幼蟲發育的影響
14℃組中的刺巨藤壺幼蟲只能發育到Ⅲ期無節幼蟲階段,34℃組僅發育到Ⅴ期無節幼蟲階段,18—30℃各溫度組無節幼蟲均能發育至金星幼蟲階段(圖9)。各溫度組Ⅵ期無節幼蟲的存活率均達到70.00%以上,以22℃的存活率最高為86.67%。金星幼蟲的存活率22℃時最高為50.67%,30℃時的最低僅9.67%。經過方差分析表明,溫度對Ⅵ期無節幼蟲(F=4.401,P=0.042)和金星幼蟲存活率(F=13.945,P=0.002)均有顯著影響。經過多重比較,18℃和22℃組間的Ⅵ期無節幼蟲存活率有顯著差異,18℃、26℃、30℃各組間的沒有明顯差異(P≥0.473),22℃、26℃、30℃各組間的也沒有明顯差異(P≥0.090)。30℃組金星幼蟲存活率與其它各組間的均有顯著差異,其它3個溫度組間的存活率無顯著差異(P≥0.134)。
隨著溫度的提高,Ⅵ期無節幼蟲和金星幼蟲發育時間均縮短(圖10)。以18℃組的幼蟲發育最慢,Ⅵ期無節幼蟲和金星幼蟲發育時間分別需要9 d和14 d;以30℃組的幼蟲發育最快,Ⅵ期無節幼蟲和金星幼蟲發育時間分別需要5 d和7.3 d。
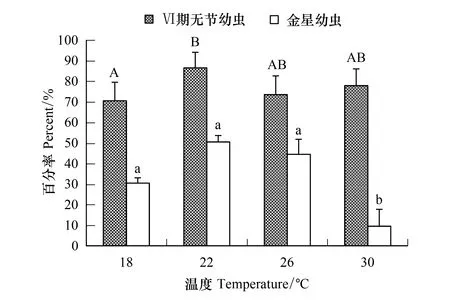
圖9 溫度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲存活率的影響
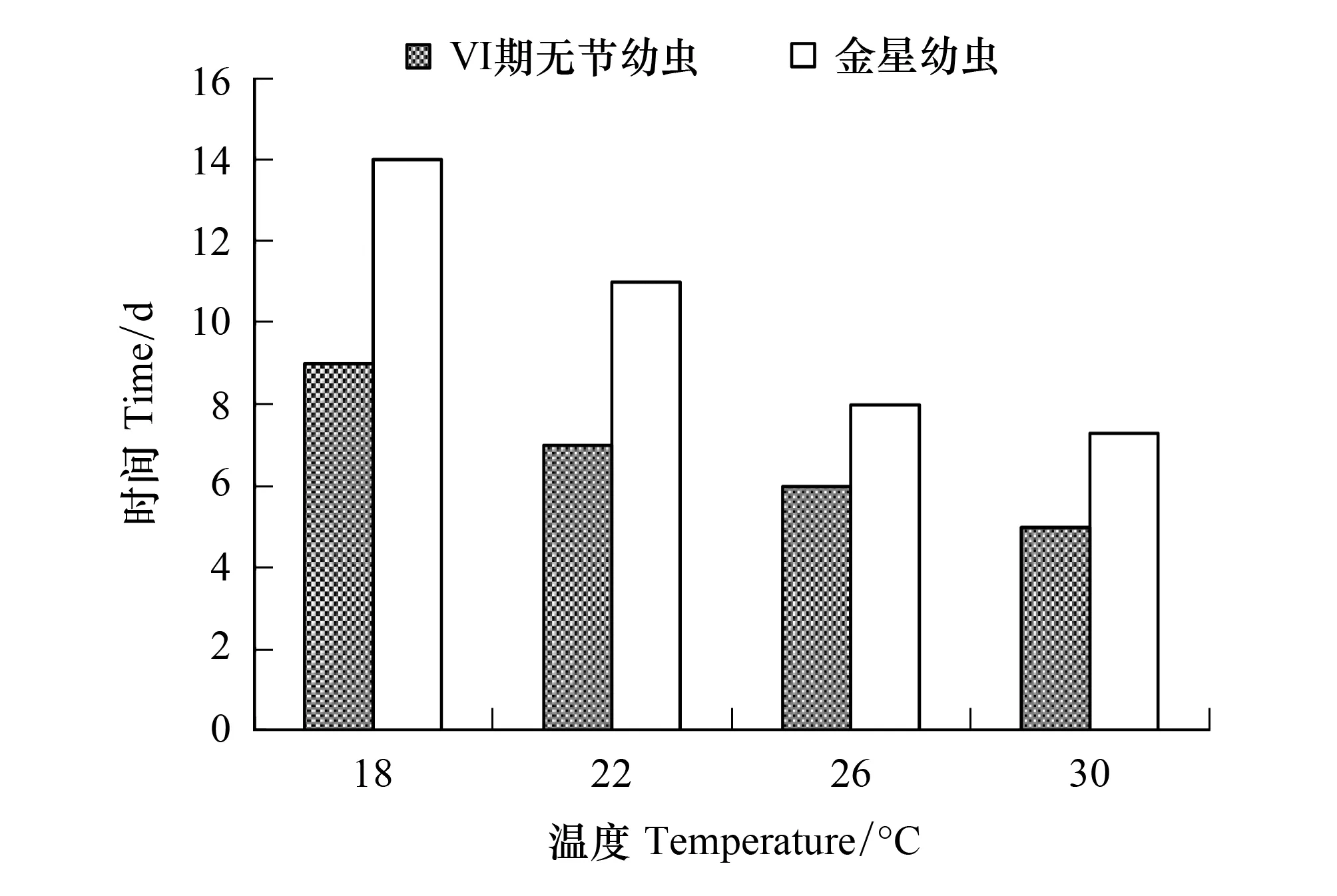
圖10 溫度對刺巨藤壺Ⅵ期無節幼蟲和金星幼蟲發育時間的影響
溫度對刺巨藤壺金星幼蟲體長的影響見表5。18℃組的金星幼蟲體長最長,30℃組的最短。經單因素方差分析表明鹽度對金星幼蟲體長有極顯著影響(F=18.751,P=0.000)。經過多重比較,18℃組和其它各溫度組間的金星幼蟲體長有極顯著差異 (P< 0.01),其它3個溫度組間的沒有顯著差異(P≥0.178)。低溫培養下,金星幼蟲的體長相對較長。

表5 溫度對刺巨藤壺金星幼蟲體長的影響
3 討論
3.1 餌料種類和密度對刺巨藤壺幼蟲發育的影響
餌料、鹽度和溫度等多種生態因子影響海洋無脊椎動物幼蟲的生長發育和變態,其中最重要的因子是餌料。合適的餌料是動物培養的關鍵,只有找到合適的餌料才有可能在室內進行動物的人工培養[4,7—8]。蔓足類的生活史包括無節幼蟲和金星幼蟲2個幼蟲階段,金星幼蟲階段是不攝食的,因而無節幼蟲的攝食經歷決定了金星幼蟲的能量儲備以及隨后的募集成敗[2,9]。在實踐中,培養各種蔓足類幼蟲適宜的微藻不同。曹文浩等認為旺盛期的亞心形扁藻是培養網紋藤壺幼蟲的適宜餌料[11]。馮丹青等用牟氏角毛藻可將白脊藤壺 (B.albicostatus)幼蟲培養至金星幼蟲階段[22]。Desai等用角毛藻(C.calcitrans)培養紋藤壺幼蟲的效果比中肋骨條藻的好[7]。Nasrolahi等用角毛藻(C.calcitrans)和小球藻可以培養致密紋藤壺(A.improvisus)幼蟲到金星幼蟲階段;而四尾柵藻 (Scenedesmusquadricauda) 不合適培養致密紋藤壺幼蟲,可能是因為它表面有刺的形態妨礙了幼蟲的攝食[8]。Nunes等用5種微藻培養M.azoricus的無節幼蟲,結果表明等鞭金藻(T-Isochrysissp.)和一種骨條藻的效果較好、杜氏鹽藻(Dunaliellasp.)效果一般、而角毛藻(C.calcitrans)和擬衣藻(Chloromonassp.)效果最差[20]。Franco等的結果表明瑪氏骨條藻 (S.marinoi) 培養有柄類指茗荷 (Pollicipespollicipes) 無節幼蟲的效果較肩突四鞭藻 (Tetraselmissuecica) 好,而用球等鞭金藻 (I.galbana)培養的幼蟲不能發育到金星幼蟲階段[23]。Thiyagarajan等進一步指出不同餌料(纖細角毛藻C.gracilis、硅限制的纖細角毛藻、假微型海鏈藻Thalassiosirapseudonana和中肋骨條藻)培養的紋藤壺金星幼蟲的營養指標如三酰甘油(TAG)/DNA的比值不同,而金星幼蟲TAG/DNA的比值越高,其附著變態能力越強[9]。
綠藻、金藻和硅藻3個門的薇藻在海洋動物的培養中被廣泛使用。微藻餌料的營養價值取決于很多因素,藻類營養成分的含量和比例(脂肪酸成分、氨基酸成份、碳水化合物水平、必要維生素等)決定了微藻的食用價值[4,7,20]。而微藻的大小、形狀和藻鏈的長度均會影響它的沉降率,進而影響蔓足類幼蟲的攝食和消化效率[24-25]。隨著蔓足類幼蟲的發育,游泳附肢剛毛間的距離漸增大,晚期幼蟲比早期幼蟲可以攝食更大的餌料顆粒[25],如致密紋藤壺早期無節幼蟲用小球藻的效果比角毛藻的好,而晚期幼蟲則相反[8]。因此,不同微藻的餌料效果不同,不同的蔓足類及同一種蔓足類不同的發育階段適宜的微藻也不同。
本實驗中所用的3種微藻均能成功培養刺巨藤壺無節幼蟲發育到金星幼蟲階段,但用中肋骨條藻和牟氏角毛藻培養的刺巨藤壺金星幼蟲存活率和金星幼蟲體長都比用亞心型扁藻培養的高得多。因此,中肋骨條藻和牟氏角毛藻是培養刺巨藤壺無節幼蟲適宜的餌料,而亞心型扁藻并不是合適的餌料。3種微藻細胞大小、形態、營養價值等的不同,導致了刺巨藤壺無節幼蟲產生了選擇性的差異。牟氏角毛藻細胞小、壁薄、多數營單個生活,大小為3.5×4.6—4.6×9.2 μm,也有 2—3個細胞相連組成群體生活;中肋骨條藻其細胞大小為 4—15 μm,呈圓柱形,褐色,藻體由幾個到幾十個細胞靠殼面相連成鏈狀;扁藻為綠藻門、具鞭毛,長為11—14μm,寬7—9μm[26]。如上所述,雖然有多種微藻可作為蔓足類幼蟲的餌料,除少量報道扁藻可用于蔓足類幼蟲的培養外[11—13,27],用于蔓足類幼蟲培養的微藻普遍為硅藻類的角毛藻和骨條藻[4—6,10,28—30]。角毛藻和骨條藻同屬硅藻門,富含二十二碳六烯酸 (DHA)、二十碳五烯酸 (EPA) 等高不飽和脂肪酸,是蔓足類適宜的餌料[7,24,31]。本實驗用高密度扁藻培養的刺巨藤壺Ⅵ期無節幼蟲的存活率均高達70%以上,但金星幼蟲階段的存活率急劇下降。扁藻比2 種硅藻大,雖然藻類大小是影響幼蟲攝食效率的一個因素,但蔓足類晚期幼蟲比早期幼蟲可以攝食更大的餌料顆粒[25],藻類大小顯然不是影響刺巨藤壺幼蟲攝食扁藻的一個原因。對蔓足類來說,從Ⅵ期無節幼蟲變態為金星幼蟲是一個敏感時期[23],只有最終能培養到金星幼蟲階段的餌料才是合適的餌料。本文認為扁藻可能含有某種對幼蟲發育不利且具有累積效應的成分,或是缺少某些營養要素,最后導致Ⅵ期無節幼蟲變態為金星幼蟲的比例很低。因此,扁藻不是培養刺巨藤壺幼蟲合適的餌料。混合藻類往往具有更好的餌料效果[20,23],今后,可以在單種藻類結果的基礎上進一步進行兩種及以上混合藻類的實驗,以尋找到更加有效的餌料配比。
目前,文獻報道了另2種巨藤壺幼蟲的培養。在一種巨藤壺 (M.coccopoma) 幼蟲培養中成功使用纖細角毛藻作為餌料[32],本實驗結果與此一致。但本文結果和Nunes等的結果相比有很大不同[20],他認為等鞭金藻和骨條藻是M.azoricus幼蟲合適的餌料而角毛藻不是合適的餌料。本實驗中也曾采用湛江叉鞭金藻培養刺巨藤壺無節幼蟲(結果沒有顯示),但效果很差,幼蟲不能發育到金星幼蟲階段。Nunes分析角毛藻的大小(4×8 μm)與等鞭金藻的大小(5—6 μm)相近,但在沒有充氣的條件下角毛藻沉降速率快,不利于幼蟲的攝食;骨條藻雖與角毛藻同為硅藻,也有下沉的不利因素,但為鏈狀且較大(7×16 μm),從而不影響幼蟲的攝取[20]。本實驗因為在燒杯中進行,水體小,幼蟲的密度低,同時為避免充氣后幼蟲可能會粘附在壁上影響計數,本結果是在沒有充氣的條件下取得的。后續實驗可在充氣的條件下進行,藻類不會下沉且分布會更加均勻,更有利于幼蟲的攝食,可能會得到不同的結果。本實驗與Nunes的實驗都是在沒有充氣的條件下進行,金藻和角毛藻在兩種巨藤壺表現出不同餌料效果的原因仍需作進一步的分析。
餌料的密度也會影響幼蟲的存活率、發育速度和幼蟲的大小;一般隨著藻密度的增加,存活率上升,發育速率加快,培養的幼蟲更大,營養物質積累更多。如角毛藻和小球藻的密度對紋藤壺幼蟲的發育速度、無節幼蟲的大小和營養狀況(以RNA含量為指標)均有影響[4,8]。West等認為食物密度對象牙紋藤壺無節幼蟲的大小沒有影響但對金星幼蟲的大小仍有影響[10]。Thiyagarajan等的結果表明4種微藻除了骨條藻外紋藤壺金星幼蟲的TAG/DNA比值也與藻密度相關[9]。金星幼蟲是蔓足類由浮游生活向固著生活的轉變階段,金星幼蟲的作用是尋找、探究合適的基質并最終附著完成變態。有許多物理、化學和生物的因子影響金星幼蟲的附著和變態,其中金星幼蟲的營養狀態和能量儲備是決定其變態能否成功的關鍵因子[1—3,33]。合適的藻類和密度培養得到的金星幼蟲個體比較大,顯微鏡下可見大量油滴,運動活潑,具有很高的RNA含量或TAG/DNA含量,最終附著變態的比例顯著提高[4,9,23]。
同樣,在本實驗中,3種微藻的密度均影響刺巨藤壺幼蟲的發育速度和存活率;高密度組幼蟲的發育時間短、存活率高,低密度組幼蟲發育速率降低甚至不能完成發育。中肋骨條藻低密度10×104個/mL組金星幼蟲的發育時間需要12 d,存活率僅2.33%;高密度80×104個/mL組金星幼蟲的發育時間僅需要6 d,存活率高達41.33%。牟氏角毛藻低密度10×104個/mL組金星幼蟲的發育時間需要12 d,存活率僅5.33%;高密度80×104個/mL組金星幼蟲的發育時間僅需7 d,存活率高達43.67%。除亞心形扁藻培養的金星幼蟲大小不受藻密度影響外(比其它2種藻培養的小得多),用中肋骨條藻和牟氏角毛藻培養的金星幼蟲大小均受藻密度的影響,高密度藻培養的金星幼蟲體長比低密度的長。綜合幾個觀察指標,中肋骨條藻和牟氏角毛藻培養刺巨藤壺幼蟲的適宜密度是40×104個/mL和80×104個/mL。本實驗用適宜密度的中肋骨條藻和牟氏角毛藻培養的刺巨藤壺金星幼蟲個體大,油滴多,質量更高;用亞心形扁藻培養的金星幼蟲個體明顯小,油滴也少,質量較差。
3.2 鹽度對刺巨藤壺幼蟲發育的影響
海水鹽度是影響蔓足類分布的關鍵因素[5—6,29—30,34—36]。一些低鹽種類如泥藤壺 (B.uliginosus) 和白脊藤壺 (B.albicostatus) 廣泛分布于我國沿海河口等咸淡水交匯水域;而三角藤壺、紅巨藤壺、鐘巨藤壺 (M.tintinnabulumtintinnabulum)、日本笠藤壺 (Tetraclitajaponica) 等高鹽種類則只分布于海水鹽度較高的區域[14]。很多研究表明在室內培養蔓足類幼蟲合適的鹽度往往與成體分布海域的鹽度相吻合。如網紋紋藤壺和鱗笠藤壺 (T.squamosasquamous) 當水體中的鹽度≤18,會嚴重阻滯發育甚至導致其死亡;當鹽度≥24時,這 2 種藤壺幼蟲的成活率雖不會隨鹽度改變發生顯著變化,但鹽度為 30的水體更有利于網紋藤壺幼蟲的發育[12]。這一特性與它們分布位置不同相一致,網紋藤壺主要分布在潮間帶至潮下帶,鱗笠藤壺主要分布在無遮蔽海岸或灣口附近的潮間帶。在7—36鹽度范圍內致密紋藤壺無節幼蟲均能發育至金星幼蟲,以7—24鹽度金星幼蟲存活率更高,說明致密紋藤壺更適應低鹽的水體;這與致密紋藤壺是廣鹽性種,多適應在咸淡水域中有關[34]。
本研究在22—42鹽度范圍內刺巨藤壺無節幼蟲均能發育至金星幼蟲,各鹽度梯度幼蟲發育時間相差不大,金星幼蟲的存活率以26—34鹽度的最高。這與本種分布在低潮區或潮下帶有關,對鹽度適應的范圍相對窄些也高些。這個結論進一步說明培養蔓足類幼蟲合適的鹽度往往與成體分布海域的鹽度相吻合。在低鹽條件下(22、26)培養的刺巨藤壺金星幼蟲體長較其它高鹽度的長,而致密紋藤壺無節幼體在高鹽條件下更長[34]。兩者的結果說明在過高或過低的鹽度環境中個體的大小會增加。面對調節滲透壓的壓力,海洋動物會通過形態、行為或生理等方面作出相應的適應調整[34],致密紋藤壺是在高鹽下個體更長,而刺巨藤壺是低鹽下個體更長。
3.3 溫度對刺巨藤壺幼蟲發育的影響
除鹽度外,溫度也是影響蔓足類生長發育、繁殖及其分布狀況的關鍵因素。溫度降低會延緩幼蟲發育,降低變態率,類似現象在三角藤壺[6]、籬藤壺(Elminiusmodestus)[29]、紋藤壺[6,30]、象牙紋藤壺[37]和巨藤壺 (M.coccopoma)[32]等均有報道。
網紋紋藤壺幼蟲發育及附著的適宜溫度為18—30℃,12℃低溫對其發育影響不大但不能附著[13]。網紋紋藤壺為熱帶亞熱帶暖水種,調查表明在網紋紋藤壺終年均可繁殖附著的湛江港海域其全年水溫約為17—30℃[38],這與上述結果顯示的網紋紋藤壺金星幼蟲附著所需的溫度狀況基本一致。隨著溫度的上升,龜足(Capitulummitella)幼蟲發育時間從24℃的11 d縮短為30℃的7 d;幼蟲可在 21—33℃發育,而適宜的溫度是24—30℃[39—40]。這與龜足分布海區繁殖期的水溫是19.5—27.2℃、3次繁殖高峰期水溫均高于25℃相吻合[41]。溫度也顯著影響指茗荷幼蟲的發育速度,隨著溫度提高,發育時間縮短;高溫和低溫條件下金星幼蟲存活率較低,存活率以15℃和20℃的較高,這與分布海域其繁殖季節的溫度相一致[23]。因此,室內培養蔓足類幼蟲合適的溫度往往與成體分布海域繁殖季節的溫度相一致。
在本實驗中溫度對刺巨藤壺幼蟲發育時間和存活率有顯著影響。在一定的溫度范圍內,隨溫度的上升發育時間縮短,18℃時金星幼蟲發育時間長達14 d,而30℃時僅需7.3 d。在過低(14℃)或過高(34℃)的溫度幼蟲均不能完成發育,在18—30℃無節幼蟲均能培養至金星幼蟲,但以22℃和26℃的金星幼蟲存活率最高。溫度同樣會影響個體的大小。在20℃培養的紋藤壺金星幼蟲比30℃的金星幼蟲小[4],來自Helgoland島籬藤壺的Ⅵ期無節幼蟲個體在低溫培養下更大[29],而本結果同樣在低溫(18℃)培養的刺巨藤壺金星幼蟲體長更長。綜合而言,刺巨藤壺幼蟲培養的適宜溫度是22—26℃。這也與其分布在熱帶和亞熱帶海域低潮區或潮下帶相關。
