溫帶草原7種針茅植物根系特征及其對環境因子變化的適應
2022-11-26 03:00:18袁野梅柳雋瑤高秀麗王仁忠
生態學報 2022年21期
袁野梅,柳雋瑤,高秀麗,薛 璟,4,王仁忠,4,*
1 廣東培正學院統計學系,廣州 510830
2 陜西省林業科學院黃土高原水土保持與生態修復國家林草局重點實驗室,西安 710082
3 中國科學院植物研究所,北京 100093
4 中國科學院大學,北京 100049
在陸地生態系統中,根系不僅是植物獲取水分、養分及同化物分配的重要器官,也是植物適應環境變化的重要構件,因此受到廣泛關注[1—2]。根系作為水分吸收器官對干旱極為敏感,不僅根系外部形態性狀因干旱而產生變化[3—4],根系解剖結構[5—6]和生理調節物積累也表現出一定的變化[7—9],從而形成應對環境變化的綜合適應機制。但以往的研究多集中在生態學、解剖學或生理生態學等某一特定層次上,缺乏多學科多層次的系統性研究[10],因此在根系生物量、解剖結構和生理調節等多層次上系統研究植物對環境變化的綜合適應機制對探討未來氣候變化下植物適應策略及分布區域變化等有著重要的理論和實踐意義[11]。
針茅屬植物廣泛分布于我國西部干旱和半干旱區的天然草地,是荒漠草原區主要牧草資源。關于針茅屬植物的研究報道很多[12—13],主要集中在特定區域或某一種針茅植物的特征[14—17]、草地管理方式對針茅植物根系的影響[18—19]等方面,關于大尺度梯度下多種針茅植物根系性狀的比較、與區域環境因子的關系及未來氣候變化下適應策略和分布區域變化等尚不明確。本研究以溫帶草原8個針茅草地的7種針茅植物根系為研究對象,系統研究了針茅屬植物根系生物量、根冠比(R/S)、解剖結構和生理調節物積累等性狀的變化規律,并分析了根系性狀與環境變化的關系,旨在為探究未來氣候變化下針茅植物適應策略及不同針茅植物分布區更替趨勢等提供科學依據。
1 材料和方法
1.1 研究區自然概況
研究區域涵蓋了我國溫帶主要針茅草原(106°88′—120°12′E,38°02′—49°35′N),從東北部的海拉爾到西南部的鹽池高沙窩鎮(約1600 km),東西貫穿經度約14°,南北跨越緯度約10°(表1)。海拔高度呈現由東北至西南增加的趨勢,位于東北端的海拉爾(HL)平均海拔為679 m,錫林郭勒白音錫勒牧場(BY)和多倫(DL)海拔高度逐漸升高至1000 m以上,達茂旗召河(ZH)和烏拉特后旗(UR)地勢隆升,海拔高于1600 m,烏海四合木保護區(WH)、鄂爾多斯草原(OR)及寧夏鹽池高沙窩鎮(GS)海拔在1300—1400 m之間。
該區域屬溫帶大陸性季風氣候,夏季風弱、冬季風強,氣候干燥,日照豐富。近50年(1970—2018年)氣象數據統計表明區域年平均氣溫在-0.8—8.5℃之間,最低溫多出現在1月,最高溫均出現在7月,年均氣溫總體呈現由東向西及由北至南逐次遞增的趨勢(表1)。降水量少而不均是限制該區域草原植物生長的首要因子,年降水量分布東多西少,北高南低,西南部烏拉特后旗和烏海年降水量小于150 mm,東北部海拉爾、多倫、白音錫勒牧場年降水量高于350 mm,是西南部樣點年降水量的2倍多。春季和初夏(4—6月)會經歷較為明顯的季節性干旱(1—6月降水量約占全年的30%),降水量峰值出現在7—8月(降水量約占全年的50%)。降水和氣溫的梯度變化導致植被分布呈現出明顯地帶性格局,由東北到西南依次為草甸草原、典型草原和荒漠草原或荒漠,土壤主要為鈣土或暗栗鈣土、栗鈣土、風沙土、灰鈣土、漠鈣土和棕漠土等,主要針茅植物依次為貝加爾針茅、大針茅、克氏針茅、本氏針茅、短花針茅、戈壁針茅和沙生針茅。
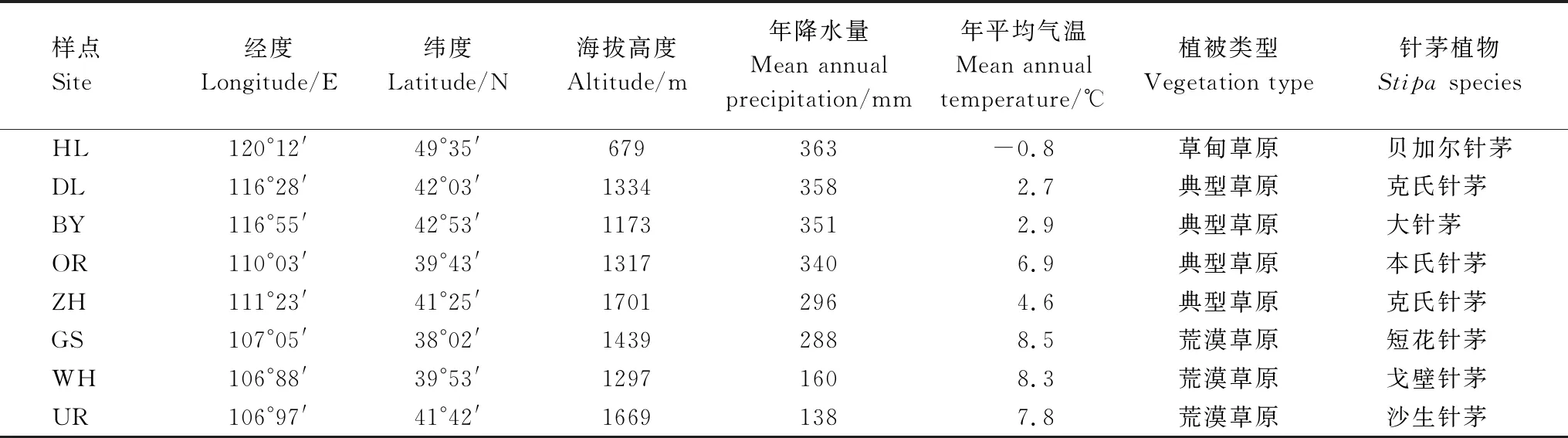
表1 大尺度梯度下實驗樣點地理位置、氣候、地帶性植被和主要針茅植物種類.
1.2 實驗方法
在研究區內選擇8個優勢種為針茅屬植物且人為干擾較少的天然草地為固定采樣點(表1),2018年4月26日—6月11日(旱季)開花期和7月28日—9月5日(雨季)果后期分別進行野外采樣。為減少不同樣點植物生長節律差異的影響,旱季由南至北(西南部返青早于東北部),雨季由北至南進行調查取樣(北部枯死期早于南部)。在各樣點選取3個天然針茅草地作為采樣區(面積大于1 hm2,且土壤條件基本一致),每個采樣區隨機設置5個1 m×1 m樣方,在樣方內取1個包含中等大小針茅株叢的20 cm×20 cm×20 cm土柱,將土柱分成0—10 cm和10—20 cm2層(涵蓋了針茅植物90%以上根系生物量),分別裝入塑料袋帶回實驗室。地上部分分成針茅和其他植物,分別裝入紙袋,烘干稱重。
用1 mm孔徑篩子篩出每層土樣中的土粒,將植物根裝入90目尼龍網袋,用清水沖洗干凈。去除雜物和其他植物根系,得到針茅根系樣品,用吸水紙將植物根表面的水分吸干。剪下針茅根蘗,稱根系樣品鮮重。依據顏色、彈性、韌性等將其分為活根和死根,活根再分為吸收根(主要是當年新生1—3序級細根)和運輸根(主要是大于3序級的往年細根)[20—21],用信封分裝,放入烘箱,105℃殺青1 h,65℃烘48 h,稱干重。根冠比(R/S)為地下根系生物量與地上生物量之比。
在每個土柱中取2—3段針茅植物活根(從根尖開始5 cm長,主要是當年生的吸收根)放入FAA (冰醋酸∶甲醛∶70%乙醇=1∶1∶18)固定液固定。采用石蠟切片法[5],經過軟化、抽真空、脫水、透明、浸蠟以及石蠟包埋,用切片機(Leica,RM2235,Germany)切成8—10 μm切片,最后染色制片。利用NIS-Elements(Nikon,Japan)圖像分析軟件測定根切片的導管(Vea)和中柱(Stea)直徑,表皮厚度(Ept)、皮層厚度(Cort)、內皮層厚度(Ent)、內皮層徑向壁厚度(Rawt)及根直徑(Rd)等[10]。
烘干稱量后的各層活根樣品經球磨儀粉碎過100目篩,粉末樣品用于測定抗旱生理指標脯氨酸(Pro)和可溶性糖(SS)含量。取0.1—0.5 g樣品粉末加入磺基水楊酸(10 mL,3%)進行提取,經濾紙過濾后取上清液2 mL,茚三酮法測定Pro含量[10]。取50 mg樣品粉末置于80%乙醇萃取液,85℃靜置1 h。然后高速離心(3000轉)10 min,如此反復用80%乙醇萃取三次,合并上清液,加入活性炭脫色,在真空干燥器里干燥,最后用蒸餾水定容,蒽酮比色法測定SS含量[10]。
在各樣方中采集0—20 cm土樣,風干后研磨粉碎過0.15 mm篩。取0.1—1.0 g土樣,重鉻酸鉀滴定法測土壤碳含量。取0.1—1.0 g樣品,凱氏定氮儀(FOSS2200,Denmark)測定土壤全氮含量。
1.3 數據處理
采用Excel 2016和R 3.4.1軟件對各樣點旱季和雨季針茅植物地上地下生物量、R/S、Vea、Stea、Ept、Cort、Ent、Rawt、Pro和SS含量等數據進行統計分析,方差分析比較各樣點間及同一樣點旱雨季間針茅根系地上地下生物量、R/S、Vea、Stea、Ept、Cort、Ent、Rawt,Pro和SS含量的差異。利用一元線性回歸分析了針茅根系生物量、R/S、Ept/Rd、Cort/Rd、Ent/Rawt、Vea/Stea、Pro和SS含量與旱季和雨季降水量和氣溫間的關系(旱季和雨季降水量和氣溫分別指1—6月和1—8月降水量和平均氣溫,涵蓋了所有時段降水和氣溫對植物性狀的影響)。采用獨立性較高的旱季和雨季平均氣溫、降水量、海拔作為環境變量對針茅植物上述性狀進行偏相關分析(控制其中兩個因子,分析各指標與第3個因子的關系)。本研究氣象數據來源于中國氣象數據網(http://www.worldclim.org) 1970—2018年地面常規觀測資料。
2 結果
2.1 根系生物量和根冠比
無論是旱季還是雨季,除BY樣點的大針茅具有較高的根系生物量外,在由東北到西南的大尺度區域內針茅植物根系生物量基本呈現增加的趨勢(圖1)。除了旱季OR與ZH、雨季DL、OR與ZH相鄰樣點間外,其他各樣點0—10 cm土層根系生物量差異均達到顯著水平(P<0.05),旱季DL和ZH樣點的克氏針茅0—10 cm土層根系生物量差異也達到了顯著水平。旱季和雨季,HL和DL樣點0—10 cm土層根系生物量僅為UR和BY樣點的7%和17%。8個樣點10—20 cm土層針茅植物根系生物量變化基本與表層一致,BY樣點根系生物量最大,顯著高于其他樣點(P<0.01),旱季約為最低樣點(HL)的6.1倍、雨季約為最低樣點(DL) 5.3倍。旱季OR與UR、ZH與GS間及雨季GS與WH間10—20 cm土層根系生物量差異不顯著(P>0.05)。旱季和雨季各樣點0—10 cm土層針茅根系生物量均顯著大于10—20 cm土層(P<0.05),且表層平均根系生物量為總根系生物量的85%。在研究區域內,針茅植物R/S差異顯著,HL和DL樣點R/S最低,旱季由BY至WH逐漸降低,最低值(DL)僅為BY和UR樣點的20%和13%,UR樣點針茅R/S最大。雨季由東北到西南R/S基本呈增加的趨勢,UR樣點的R/S約為HL樣點的8倍。
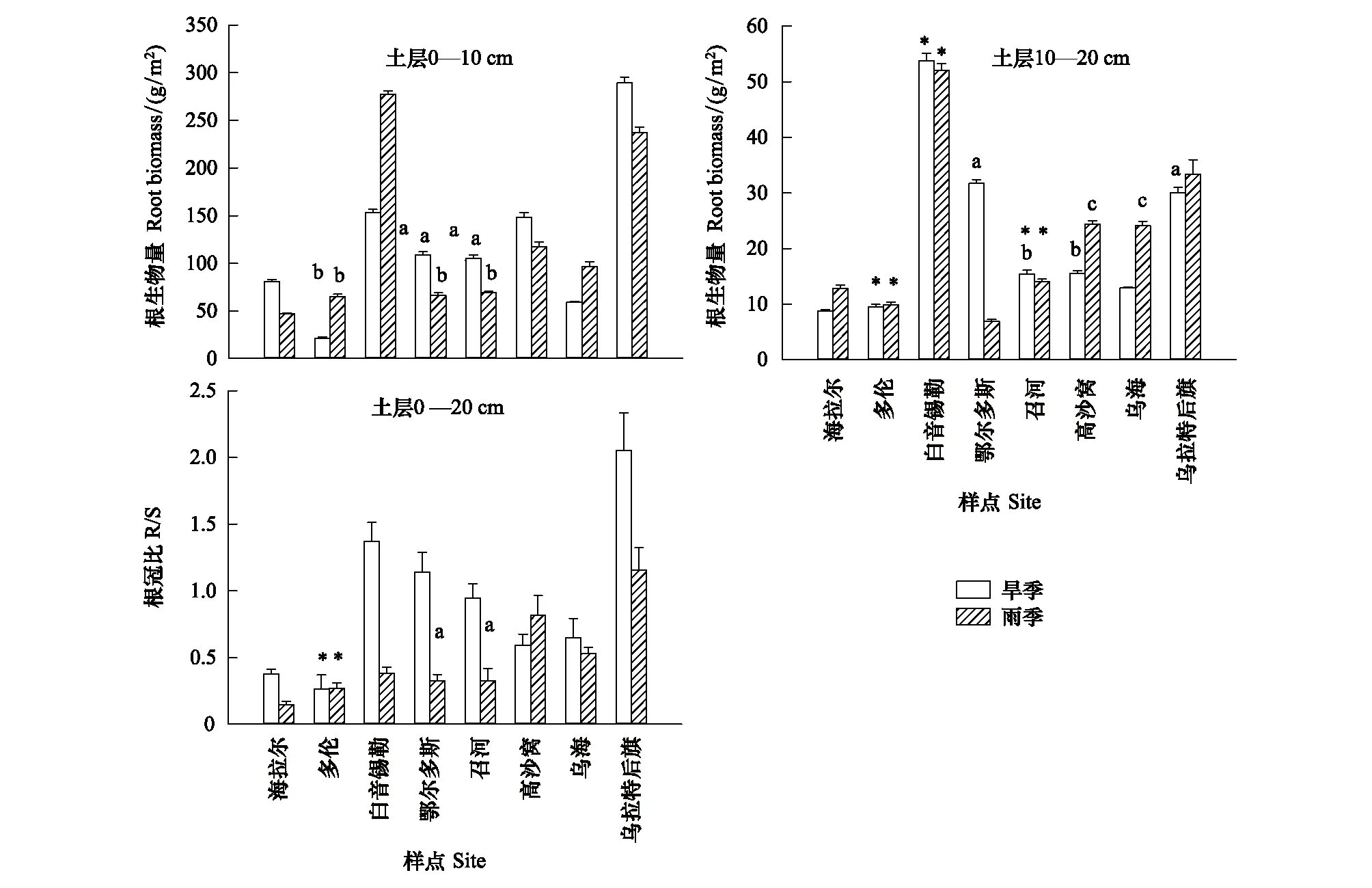
圖1 大尺度梯度下旱季(DS)和雨季(RS)各取樣點0—10 cm和10—20 cm土層針茅植物根系生物量和根冠比的比較(平均值±標準誤)
2.2 運輸根和吸收根
旱季和雨季,各針茅植物0—10 cm土層根系中運輸根占比最大,平均為64.4%和65.4%,且由東北至西南呈逐漸增大的趨勢(圖2),吸收根平均占比分別為20.7%和17.4%,其變化趨勢與運輸根相反。在10—20 cm土層根系中,運輸根平均占比下降(圖2),旱季和雨季分別為31.0%和38.3%,二者差異顯著(P<0.05),吸收根比例分別增大到49.0%和36.2%,且旱季顯著大于雨季(P<0.01)。雨季下層土壤中死根比例有所增加,達到25.4%,是旱季的3倍。旱季和雨季,BY、ZH、OR和WH樣點針茅植物0—10 cm的吸收根與運輸根比(Ra/Rt)差異顯著(P<0.05),BY樣點大針茅旱季Ra/Rt約是雨季的2倍。GS和HL樣點10—20 cm土層中Ra/Rt旱季和雨季差異顯著,GS樣點短花針茅旱季是雨季近3倍。
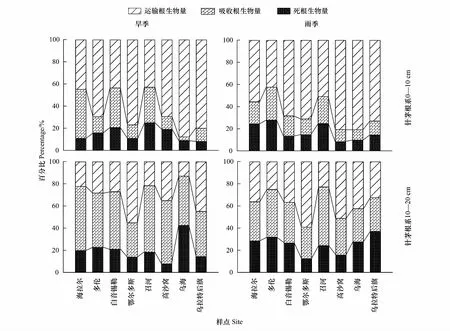
圖2 旱季(DS)和雨季(RS)針茅根系0—10 cm和10—20 cm土層運輸根(Rt)、吸收根(Ra)和死根(Rd)生物量百分比堆疊圖
2.3 根解剖結構
在實驗區域內,旱季和雨季針茅植物根直徑(Rd)、表皮厚度(Ept)和皮層厚度(Cort)表現出不同的變化規律(表2)。旱季針茅Rd、Ept和Cort基本表現為由東北至西南呈下降趨勢,但在最西部的UR各指標均出現明顯上升,而雨季總體呈由東北到西南的上升趨勢。旱季HL樣點Rd比雨季高45% (P<0.01),而其他樣點均為雨季高于旱季。旱季東北部3個樣點的Ept和Cort均顯著高于雨季(P<0.05),而西南部5個樣點則低于雨季。
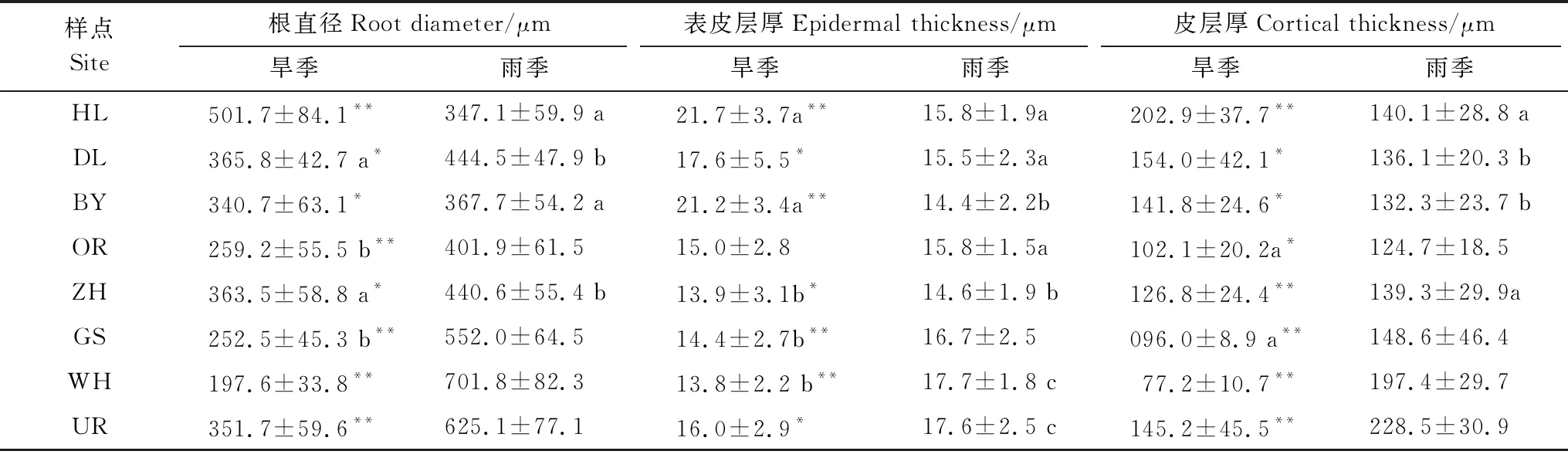
表2 旱季和雨季針茅根直徑、表皮厚度及皮層厚度 (平均值±標準誤)
各針茅植物根系的內皮層均有不同程度的馬蹄形加厚,旱季針茅根系內皮層(Ent)和徑向壁厚度(Rawt)由東北至西南有下降趨勢,但在UR樣點顯著增加(P<0.01),而雨季無明顯趨勢(表3)。旱季東北部3個樣點的Ent和Rawt均顯著高于雨季(P<0.05),其余5個樣點的Ent則是雨季高于旱季。8個樣點中,針茅植物中柱(維管束)直徑(Stea)的種間差異顯著,但無明顯規律。導管直徑(Vea)的變化與Ent相似,但雨季大體由東北至西南逐漸上升(表3)。除東北部3個樣點外,其余5個樣點的Stea和Vea基本為雨季高于旱季。
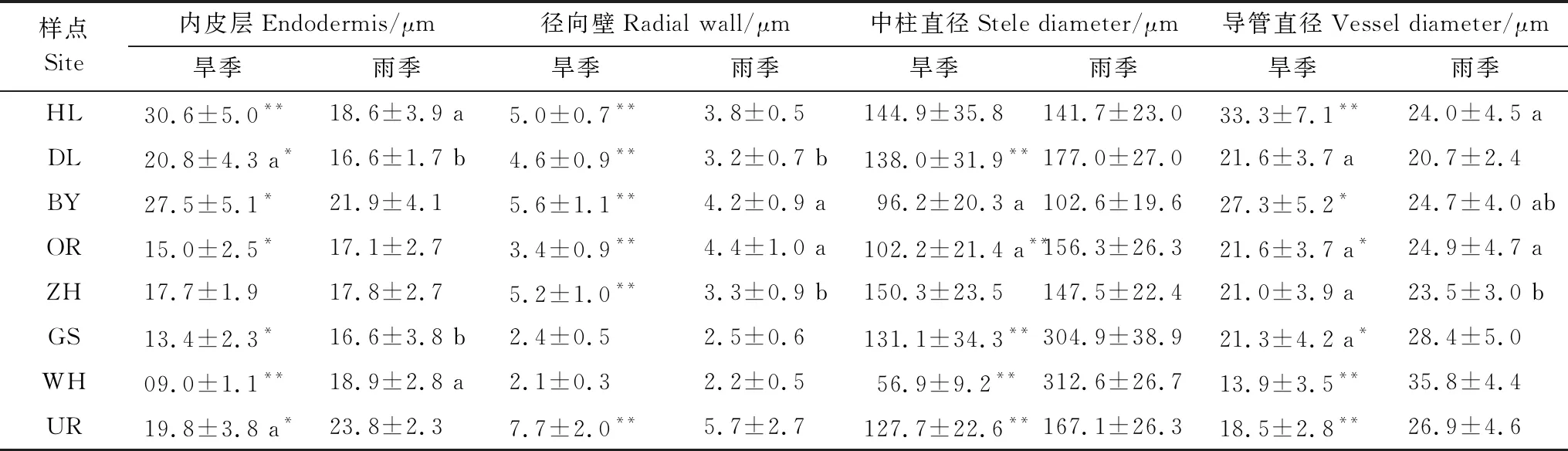
表3 旱季和雨季針茅根內皮層、徑向壁厚度、中柱和導管直徑變化(平均值±標準誤)
根表皮厚度和皮層厚度與根直徑比例(Ept/Rd、Cort/Rd)可以直觀地反映植物根對表皮的物質投入比例及其適應功能。各樣點針茅植物Ept/Rd和Cort/Rd基本為旱季大于雨季(ZH樣點Cort/Rd除外),WH樣點戈壁針茅旱雨季差異最大,旱季Ept/Rd約為雨季的3倍(P<0.01),貝加爾針茅(HL)、克氏針茅(DL,ZH)和本氏針茅(OR)季節差異不顯著(P>0.05)(表4)。旱季各植物Ept/Rd處于波動狀態,種間差異顯著。雨季Ept/Rd總體呈現由東北向西南逐漸下降的趨勢,最大值為HL樣點貝加爾針茅,是最小值WH樣點戈壁針茅的1.6倍(P<0.05)。
內皮層與徑向壁厚度比(Ent/Rawt)在一定程度上反映了根保水能力的強弱。各樣點針茅根Ent/Rawt基本是雨季大于旱季(表4),且差異顯著(P<0.01)(GS樣點除外)。無論旱季還是雨季GS和WH樣點的Ent/Rawt均顯著高于其他樣點。導管面積與中柱面積比(Vea/Stea)體現了根系的輸水效率,該值越大說明根輸水能力越強。旱季和雨季間各種針茅植物Vea/Stea差異較小,且東北部3個樣點針茅植物間差異均不顯著(P>0.05),最西部的UR樣點Vea/Stea顯著高于其他樣點(P<0.01)。
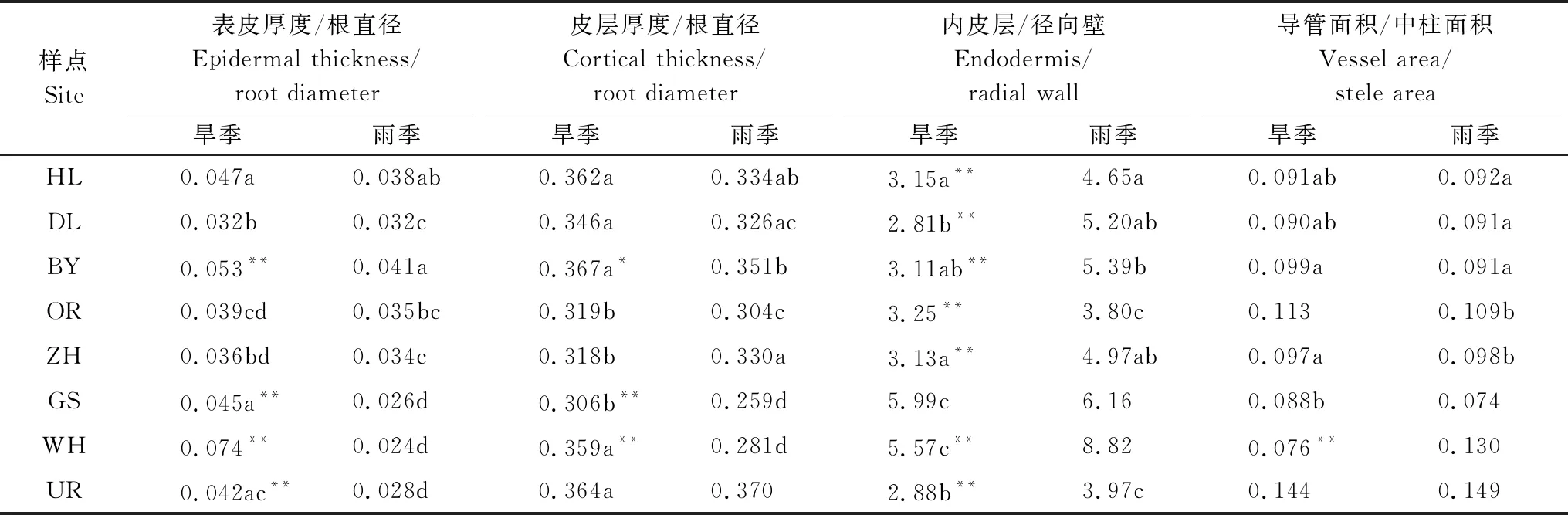
表4 旱季和雨季針茅根表皮厚度、皮層厚度與根直徑比、根內皮層與徑向壁厚度比和導管與中柱面積比
2.4 根系生理調節物
各樣點針茅根系脯氨酸含量(Pro)波動很大,由東北到西南沒有明顯的變化規律(表5),最西部的UR樣點Pro最高。旱季和雨季僅有不到一半樣點各土層根系Pro差異顯著,然而,無論是旱季還是雨季,各樣點10—20 cm土層根系Pro含量均高于0—10 cm土層(除最西部的UR樣點外)。3/4樣點10—20 cm土層根系Pro旱季高于雨季。與Pro變化相似,由東北到西南針茅植物根系可溶性糖含量(SS)沒有呈現明顯的增減趨勢(表5)。除OR樣點外,各樣點根系SS呈現顯著的季節性差異(P<0.05),西南部3個樣點旱季0—10 cm土層根系SS含量顯著高于雨季。除HL樣點外,10—20 cm土層根系SS含量均為旱季高于雨季。
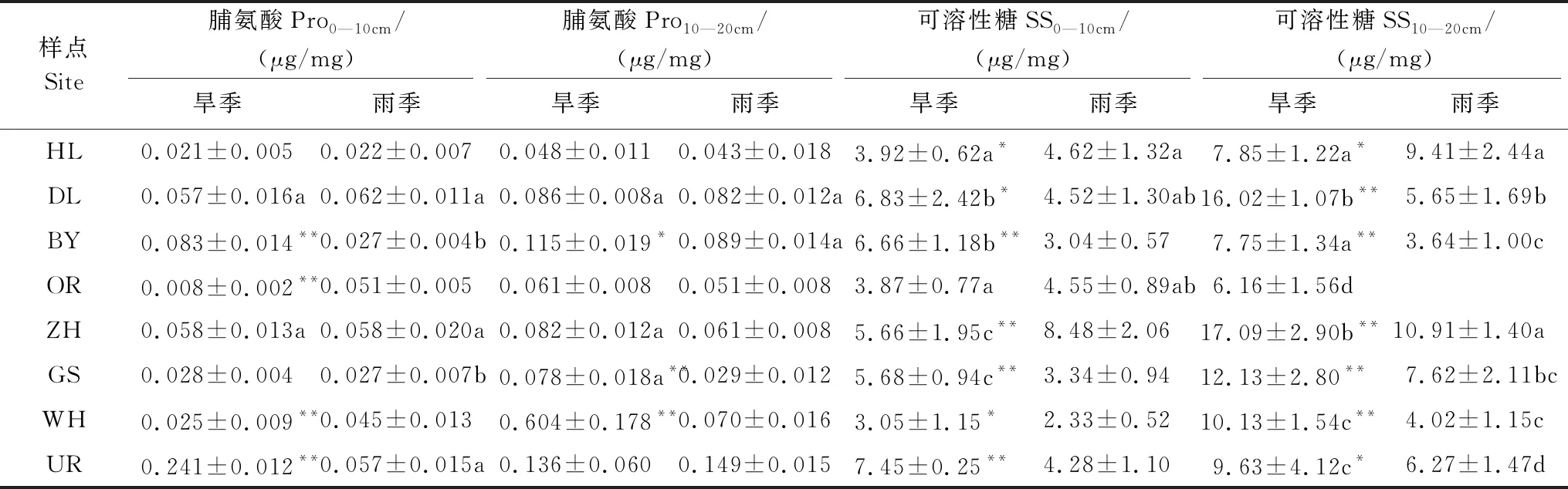
表5 旱季和雨季0—10 cm和10—20 cm土層針茅根系脯氨酸和可溶性糖含量(平均值±標準誤)
2.5 根系性狀與環境因子的相關分析
偏相關分析顯示大尺度梯度上針茅植物根系性狀與環境因子(降水量、平均氣溫和海拔高度(Alt))間存在顯著的相關性(表6)。0—10 cm土層針茅根系生物量與P1—6、P1—8、T1—6、T1—8和Alt均呈極顯著負相關關系(P<0.01); 10—20 cm土層根系生物量與P1—8和Alt呈顯著負相關關系(P<0.01),但與T1—6呈顯著正相關關系(P<0.05)。旱季針茅植物R/S僅與P1—6顯著負相關(P<0.01),但雨季R/S與P1—8和Alt均達到了極顯著負相關水平(P<0.01),且與T1—8呈正相關關系(P<0.01)。降水量、平均氣溫和海拔高度對各土層針茅運輸根生物量均呈負效應,與旱季0—10 cm土層運輸根生物量呈極顯著負相關關系(P<0.01)。旱季針茅植物吸收根生物量受到降水量和海拔高度的極顯著負效應(P<0.01),T1—6僅對表層吸收根生物量產生極顯著的正效應(P<0.01),雨季環境因子對針茅植物吸收根生物量的影響較旱季明顯減弱。

表6 大尺度梯度上根系生物量、根冠比、吸收根和運輸根生物量與氣候和海拔高度(Alt)等因子的偏相關分析
環境因子對針茅植物根系解剖性狀有顯著影響(表7),P1—6對Ept/Rd、Cort/Rd及P1—8對Cort/Rd、Ent/Rawt、Vea/Stea均有顯著負效應(P<0.01),溫度(T1—6、T1—8)對上述各指標的影響不盡相同。降水量顯著影響旱季各土層和雨季10—20 cm土層Pro含量(P<0.05),溫度對各土層根系SS有極顯著負效應(P<0.01)。海拔高度對表層根系Pro和SS含量有顯著正效應(P<0.01)。
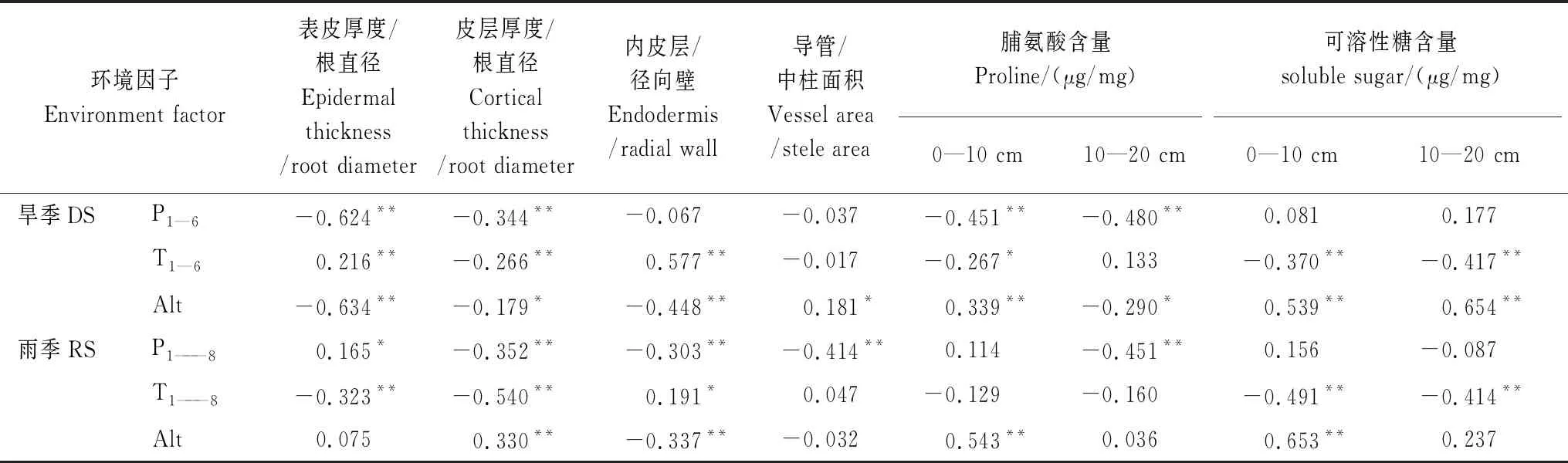
表7 旱季和雨季針茅根系解剖和生理調節特征與降水、溫度和海拔的偏相關分析
3 討論
3.1 針茅植物根系性狀差異及其適應策略變化
植物應對干旱的適應策略雖然復雜多樣,但主要包括以下兩種途徑,一是通過改變根系性狀(如增加R/S、減小內皮層厚度等)最大程度增加吸水和保水能力,二是改變植物葉片性狀(如減少氣孔密度、增加比葉重等)減少水分散失[9]。針茅植物根系對干旱極為敏感,研究其耐旱性狀變化對揭示針茅植物的適應策略和機制具有重要的理論和實踐意義。在本研究的8個樣點中,由東北至西南針茅根系生物量呈現波動增長趨勢(圖1),即體現了針茅植物的種間差異,也反映了它們對干旱的趨同適應性。這一趨勢與其他報道基本一致,Gao等對分布于不同沙丘部位兩種沙蒿根系的比較發現生長在沙丘頂部(干旱貧瘠)的白沙蒿(Artemisiasphaerocephala)比生長在沙丘底部(水分和養分相對較好)的褐沙蒿(A.intramongolica)具有更發達的根系,不僅分布較深(140 cm以上),而且生物量也高[4]。Wang等的研究顯示在典型草原和荒漠草原區隨干旱增加羊草根系生物量呈現增加的趨勢[22]。干旱區生態系統中發達的根系增強了植物吸水和保水能力,進而增強了植物適應干旱的能力[3—4]。多數樣點針茅植物在旱季維持較高的表層(0—10 cm)根系生物量,這主要是由于表層運輸根占比高(約65%,圖2),其木質素含量高周轉率低[2]。針茅植物深層(10—20 cm)根系生物量旱季和雨季差異不顯著,其原因是深層根系中周轉率較高的吸收根(或細根)占比大(圖2),相對穩定的土壤環境(主要是溫度和水分)導致吸收根新生或死亡的波動較小[4]。深層根系對降水的響應較弱表明針茅有著較強的耐旱性,其深層根系的相對穩定維持了群落生物量的穩定[23]。
Ma等的研究顯示內蒙古草原群落R/S與降水量及溫度沒有顯著相關性[13],但在本研究區域內,各針茅植物R/S普遍是旱季大于雨季,且旱季和雨季R/S與降水量均呈極顯著負相關,這與Schenk等[3]、Mokany等[24]和Wang等[22]的結果基本一致。旱季較高的R/S增強了植物吸水、保水及碳水化合物貯藏能力,有助于植物度過干旱逆境[25]。與以往報道不同的是在由東北至西南的樣帶上,旱季針茅植物R/S并沒有隨年均降水量的減少而增加(圖1),而是由BY樣點至WH樣點呈逐漸下降的趨勢,這可能與該樣帶溫度變化有關,BY樣點至WH樣點年均溫逐漸增加,升溫導致根系呼吸增強,降低生物量積累,同時升溫有助于植物生長和地上生物量增加,結果導致這些樣點針茅R/S下降。升溫與干旱脅迫對植物根系交互影響比較復雜,今后需要增加控制實驗才能準確甄別。
3.2 針茅植物根系功能和解剖結構及其對環境的適應
干旱不僅改變植物R/S,也改變根系的功能結構。本研究根據文獻[20—21]、根序級及解剖結構將針茅根系分為吸收根和運輸根兩部分,劃分雖然簡單,卻能較好地區分不同等級根系在適應干旱過程中的功能作用。表層根系中運輸根占比較大,深層吸收根比例較高,且旱季大于雨季(圖2),表明二者在應對干旱逆境時其結構和功能發生了分異。旱季吸收根生物量與降水量呈顯著負相關關系也證明了植物根系功能組成和適應策略的差異(表6)。Wang等關于干旱脅迫增加深層羊草新生根(即吸收根)生物量的報道也支持上述結論[25]。但Gao等報道在本實驗區域內兩種沙蒿植物細根生物量分布與土壤水分沒有顯著的相關性[4],因為沙蒿根系很深,主要利用深層地下水,受土壤干旱的影響較小。目前,關于植物粗根和細根的研究主要集中在碳周轉率方面[2,21],而關于根系性狀與水分關系的報道也多集中于作物研究[1,7],因而關于草原和荒漠等干旱區不同植物類群根系功能分異的系統研究仍需加強。
解剖結構的變化是植物長期生活在特定環境下而形成的穩定性適應策略,在本研究區域內針茅植物根表皮厚度(Ept)和皮層厚度(Cort)的變化與根直徑一致(表2),但是,旱季和雨季變化趨勢不同。旱季由東北至西南隨降水量的減少Ept和Cort變薄,Chimungu等研究顯示Ept和Cort變薄、細胞增大等有利吸收土壤水分,并橫向輸送至導管[26]。在最干旱的UR樣點針茅根系Ept和Cort均維持較高水平,Ept和Cort增厚雖然降低水分橫向輸導,但防止了根內水分由中柱倒流回皮層,增強了保水能力[27]。雨季由東北至西南Ept和Cort則呈增加趨勢,Ept和Cort增加,防止根內水分和養分倒流[27]。以往研究主要涉及特定時期(或一次取樣)植物解剖結構,關于解剖結構的季節性差異報道甚少[10]。本研究發現旱季和雨季不同氣候區域針茅植物根系Ept和Cort呈現不同的變化規律,東北部3個樣點Ept和Cort旱季大于雨季(表2),因為在這些樣點區域春季和夏初具有明顯干旱脅迫現象,增加Ept和Cort厚度可以起到保水作用,雨季這些區域降水量充沛,Ept和Cort厚度減少可以增加吸水的能力。西北部5個樣點Ept和Cort雨季大于旱季,這些區域常年處于干旱缺水狀態,即使雨季降水量顯著增加,水分仍然匱乏,雨季增加Ept和Cort厚度可以增強根系保水能力,有效應對干旱。旱季針茅根Ept/Rd和Cort/Rd與降水量呈顯著負相關也證明了這一點。
內皮層(Ent)和徑向壁(Rawt)調控皮層和中柱間水分和養分的運輸,控制水分和養分進入中柱,也防止中柱內溶質倒流回皮層[27]。在由東北至西南的樣帶上,針茅植物根系內皮層(Ent)和徑向壁(Rawt)厚度變化與Ept和Cort基本相似(表3),其功能作用也是類似的,這里不再贅述。WH樣點戈壁針茅和GS樣點短花針茅的Ent/Rawt顯著高于其他針茅植物(表4),體現出了較強的耐旱保水能力。水分通過橫向運輸到達中柱導管進行軸向運輸,Vea/Stea的大小在一定程度上反映了根系軸向水分運輸能力[5—6]。本研究中,針茅植物中柱直徑和維管束直徑與內皮層具有相似的變化趨勢,旱季由東北向西南逐漸下降,而雨季逐漸增加(表3),且多數樣點,旱季中柱(Stea)和導管(Vea)直徑小于雨季,干旱條件下針茅根導管直徑減小有助于根內水分貯存,為地上部分生長提供保障。Richards等[28]的研究支持這個觀點,該研究表明春小麥種子根部軸向水分運輸阻力由于導管直徑減小而增大,節約了小麥生長早期對水分的消耗,有利于其之后的生長并實現豐產。偏相關分析(表7)表明雨季針茅根Vea/Stea與降水呈負相關關系(P<0.05),這也證明在干旱區隨水分條件的改善針茅植物會選擇增大導管直徑的方式來提高導管占中柱的相對面積,增強根系水分運輸效率。中柱和維管束直徑大小與植物耐旱性關系存在爭議,其在不同植物類群和不同生長期的功能作用也不同[10],因而今后對不同植物類群及不同生長期的Stea和Vea的比較研究是必要的。
3.3 針茅植物生理調節機制差異
脯氨酸(Pro)是理想的滲透調節物質,相較于可溶性糖(SS)更加靈活,可以在脅迫發生時迅速累積[8]。本研究中,各針茅植物間Pro和SS含量均處于較大的波動狀態,最干旱的UR樣點具有相對較高的Pro和SS含量(表5),表明各針茅物種根系滲透調節物質及其功能存在明顯的種間差異。Guo等的研究顯示大尺度水分梯度上植物Pro和SS含量與干旱指數(SPEI)顯著相關,且不同植物功能群間差異顯著[10]。本研究發現針茅植物深層根系Pro和SS含量顯著高于表層(表5),這主要是深層根系基本為新生吸收根,生理活性強,是滲透調節的主要參與者。與Munns的報道相反,本研究中多數樣點針茅根系Pro旱雨季差異不顯著,而SS差異顯著(表5),說明SS在針茅植物應對干旱過程中的功能作用可能比Pro更加活躍或二者對不同環境因子的響應存在差異。偏相關分析結果顯示針茅植物根系Pro含量與降水量呈顯著負相關,而SS含量卻與均溫呈顯著的負相關關系(表7),表明針茅植物對各環境因子的響應機制不盡相同。Guo等研究表明在干旱草原區喬木Pro和SS存在著權衡關系,即在應對干旱時二者的功能是不一致的[10]。Pro和SS作為快速響應逆境脅迫的重要調節物對環境因子(干旱、升溫等)變化反應迅速,這也增加了其不穩定性,因此,要明確自然條件下他們的功能作用及相互關系需要做嚴格的控制實驗。
4 結論
在由東北至西南1600 km的研究區域內,針茅屬植物呈現明顯的更替分布(間或鑲嵌分布)狀態,總體表現為隨降水量的逐漸下降針茅植物的抗旱特征增強或適應策略趨于復雜(采用多種抗旱策略),旱季和雨季不同針茅植物根系有著不同的適應策略。針茅根系各性狀與環境因子的偏相關分析表明貝加爾針茅、大針茅及克氏針茅受干旱制約顯著,克氏針茅和大針茅對降水高度敏感,雨季降水能夠促進其快速生長。沙生針茅、短花針茅、戈壁針茅、本氏針茅通過增大根冠比、加大根系保水和吸水能力、增加滲透調節物質累積等途徑適應干旱,這些變化可能為預測未來氣候變化[29]下針茅植物分布區域變化及防風固沙能力等提供參考依據[30—31]。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
