地下水埋深和季節性干旱對古爾班通古特沙漠南緣梭梭生理和生長的影響
2022-11-26 08:18:34劉深思徐貴青米曉軍陳圖強
生態學報 2022年21期
劉深思,徐貴青,*,米曉軍,陳圖強,李 彥
1 中國科學院新疆生態與地理研究所,荒漠與綠洲生態國家重點實驗室,烏魯木齊 830011
2 中國科學院阜康荒漠生態國家野外科學觀測研究站,阜康 831505
3 中國科學院大學,北京 100049
4 烏魯木齊城北再生水有限公司,烏魯木齊 830013
干旱荒漠區因降水稀少、蒸發強烈、缺少地表徑流,荒漠植被部分或全部依賴地下水生存。地下水是決定荒漠區植被生長、分布和種群演替的主導因素之一[1],而且在干旱區生態水文過程和循環中發揮著不可替代的作用[2]。在全球變化的背景下,極端干旱事件頻發與氣候變暖加劇了荒漠生態系統的干旱程度[3]。近年來,因工業和農業的生產需求,地下水被大量開采,導致地下水儲量減少、地下水位下降和地下水質量變差。與降水相比,地下水是植物群落持續且穩定的水源。然而,地下水埋深的變化會改變生態系統的水分平衡,引起植被群落組成、水分利用策略和功能性狀進行調整[4—5]。研究地下水埋深增加引起的地下水水文干旱與植物之間的相互作用對干旱區地下水的可持續利用和生態系統的健康發展至關重要[6]。
水力性狀的調節是植物抵御干旱脅迫的重要方面。植物的水分調節包括葉水勢、水力導度、水分利用效率、水力安全閾值等[7]。地下水埋深增加引起的水文干旱,造成水分有效性降低,促使植物進行水分調整。但不同的植物對水文干旱的敏感性和適應性具有較大的差異。例如,胡楊(Populuseuphratica)通過提高水分利用效率[8]和葉片最大導水率、降低飽和膨壓和膨壓消失點滲透勢以適應水文干旱[9];古丁氏柳(Salixgooddingii)以較低的黎明前葉水勢應對水文干旱,而檉柳(Tamarixchinensis)的水分有效性則沒有改變[10]。盡管已有研究揭示梭梭(Haloxylonammodendron)的同化枝水勢與地下水埋深呈顯著的負相關[11—12],但荒漠植物水力性狀在生長季的不同階段會表現出不同的變化趨勢[13],因此深入研究梭梭的水分調節對策仍具有重要意義。
形態調節是植物在干旱脅迫下維持生存的重要生態策略。在器官水平上,根、莖、葉的表型可塑性是植物適應干旱的重要組成部分。在荒漠生態系統中,地下水埋深增加引起的水文干旱是導致植物發生形態調整的關鍵環境因子。駱駝刺(Alhagisparsifoli)幼苗增加根系生長速率[14]、胡楊降低展葉效率[15]、檉柳增加冠層枯死率[10]的策略來響應地下水埋深的增加。部分旱生植物的葉片為了減少蒸騰面積而退化為膜質鱗片,由同化枝代替葉進行同化作用[16]。此外,安富博等[17]研究表明,地下水埋深增加造成民勤綠洲人工梭梭林當年新枝生長量急劇減少。然而,在極端干旱的條件下,關于荒漠植物葉片生長速率與地下水埋深關系的研究還少有報道。
積累滲透調節物質是植物抗旱的一種重要生理適應機制[18]。有機溶質和無機離子是植物體內兩種重要的滲透調節物質,它包括可溶性糖、脯氨酸、鈉離子、氯離子等一系列滲透活性分子/離子[19]。發揮滲透調節作用的物質因物種和環境脅迫而不同。梭梭通過積累鈉離子以抵御干旱脅迫;白梭梭(Haloxylonpersicum)主要依靠積累可溶性糖來抵抗鹽堿的危害[20];胡楊通過積累可溶性糖、蔗糖、脯氨酸、甜菜堿和鉀離子等多種滲透調節物質來適應地下水埋深增加引起的水文干旱[21]。從時間尺度上來看,植物體內滲透調節物質不僅具有日變化[22],而且因物候[23]、樹齡[24]發生改變。然而,荒漠植物體內滲透調節物質和非結構性碳水化合物對地下水埋深增加的響應特征,尚不清楚。
梭梭,屬藜科梭梭屬灌木或小喬木,是古爾班通古特沙漠南緣荒漠灌叢植物的建群種之一。它具有耐貧瘠、耐干旱、耐鹽堿、抗風沙等優良特性,是荒漠地區防風固沙、保護草場、改善沙區氣候的重要植物,在維護區域生態系統結構和功能方面具有很高的價值[11]。早期研究認為梭梭應對水分脅迫(降水及其轉化而來的表層土壤水引起的水分虧缺)的一個重要策略是穩定的氣孔開度與有效的形態調節[25]。在水分條件較好時,具有高的光合速率、氣孔導度和蒸騰速率;水分匱乏時則選擇落葉并進入休眠,生長減緩或停滯[26],并認為梭梭主要依賴降水,是雨養旱生植物。但進一步研究發現,在干旱少雨甚至無雨的生長季后期,地下水是主要水源甚至唯一水源[27]。此外梭梭水分吸收策略存在年際尺度的調整,相對于干旱年份,梭梭增加了濕潤年份對表層土壤水的利用比例,但表層土壤水對植株水分來源的總體貢獻度仍不及深層土壤水和地下水[28]。由此可見梭梭主要依賴地下水,可以稱之為兼性潛水植物。當前,地下水埋深增加對梭梭生存造成了嚴重破壞,而且發現梭梭在不同區域存在大面積的退化死亡現象[29]。本文沿地下水位梯度對古爾班通古特沙漠梭梭的生理生態特征進行研究,旨在闡釋梭梭的水分調節對策、形態結構調整和滲透調節過程,這對于充分認識荒漠梭梭林的抗旱生存維持機制具有重要意義,同時有助于荒漠梭梭林的管理和保育。
1 材料與方法
1.1 研究區概況
研究區域位于古爾班通古特沙漠南緣,中國科學院阜康荒漠生態系統國家站沙漠試驗區。該區域屬典型的溫帶大陸性沙漠氣候,晝夜溫差大,夏季炎熱,冬季寒冷,平均積雪深度為20—30 cm,年均積雪覆蓋時間為100—150 d[30]。年平均氣溫為6.6℃,年平均降水量約為160 mm,年平均蒸發量約為2000 mm[25]。地勢呈南高北低,海拔370—500 m,地貌類型以固定與半固定沙丘、沙壟為主。沙丘頂部是以白梭梭植物群落為代表的荒漠植物群落,蓋度為15%—30%;壟間則以梭梭群落為主,蓋度為20%—50%[31]。物種豐富度相對較高,群落常伴生有沙漠絹蒿(Seriphidiumsantolinum)、尖喙牻牛兒苗(Erodiumoxyrrhynchum)、條葉庭薺(Alyssumlinifolim)等草本植物。
1.2 實驗樣地設置
在北沙窩野外試驗區,選擇5個實驗樣點組成地下水埋深變化梯度,實驗樣點分別記為S1、S2、S3、S4和S5(表1)。根據土壤水分狀況的季節性差異[11],將所有的實驗測量分為兩個階段:生長季前期(4—6月)和生長季后期(7—9月)。生長季前期土壤受到冬季冰雪融水和春季降水的補給;生長季后期氣溫升高,蒸發加劇,土壤水分虧缺得不到及時補充[32],淺層土壤含水量(0—100 cm)偏低(圖1)。在每個樣點選擇5棵長勢良好、株高冠幅相近的成年梭梭測量其生理指標。利用GPS測量儀(ZL180,卓林科技,安徽合肥)測量樣點的地理位置和海拔,利用激光測距儀(DL4168,得力,浙江)測量樣點地下水埋深信息。
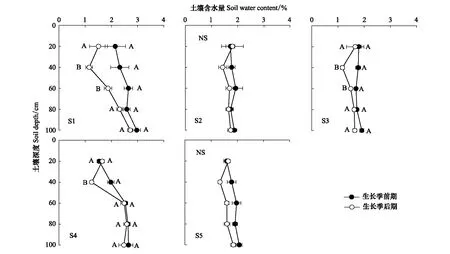
圖1 生長季前期和后期土壤含水量的變化
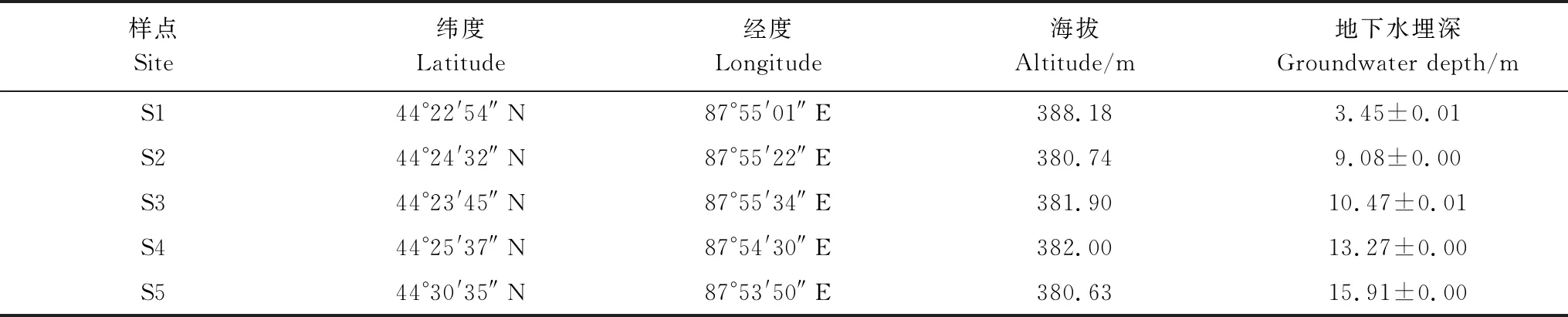
表1 樣點地理位置、海拔與地下水埋深
1.3 試驗方法
1.3.1 土壤含水量
利用土鉆在植物采樣點附近采集淺層土壤樣品,深度為1 m,土層深度分別為0—20 cm,20—40 cm,40—60 cm,60—80 cm,80—100 cm。采集的土壤樣品裝入鋁盒,用烘干法測量土壤含水量。每個實驗樣點3個重復。
1.3.2 同化枝水勢
利用植物水勢壓力室(Model 3500, PMS Instrument Company, Albany, USA) 測量梭梭同化枝的黎明前和正午水勢。黎明前水勢(Ψpd)在日出前30 min測定,正午水勢(Ψm)在13:00—14:00測定,每個實驗樣點8個重復。
1.3.3 同化枝含水量
同化枝含水量采用烘干稱重法測定。清晨剪取同化枝樣品,將其迅速放入隨身攜帶的冰盒中,帶回實驗室稱取鮮重,然后放入烘箱在75℃下干燥48 h,稱其干重,3個重復。同化枝含水量(ABWC)的計算公式為:(同化枝鮮重-同化枝干重)/同化枝鮮重×100%。
1.3.4 同化枝生長速率
梭梭同化枝生長速率在4—8月進行測定。每棵梭梭標記兩個同化枝。以白板為底,白板上帶有特定面積的黑塊,使用相機(EOS550D, Canon Inc., Japan)定期拍攝照片,拍照間隔為10 d或11 d。利用圖像處理軟件(Adobe Photoshop CS6,USA)去除照片中除實驗對象之外的其它枝條。使用CI- 400 CIAS軟件(Computer Imaging Analysis Software)批量計算出黑塊與同化枝的面積,通過黑塊實際面積與計算面積的比例關系計算出同化枝的實際面積[25]。同化枝生長速率(ABGR)計算公式,如下:
式中,St為t時刻的同化枝面積,St+1為t+1時刻的同化枝面積,Δt為兩次拍照的時間間隔。
1.3.5 胡伯爾值
在每棵梭梭上剪取一個南面生長的三級枝條。使用掃描儀(Epson Perfection 2400 Photo, Seiko Epson, Japan)對每個枝條上所有的同化枝面積進行掃描。用品紅溶液對枝條橫截面染色,用游標卡尺測量枝條的邊材和心材直徑,計算枝條橫截面的導水面積。胡伯爾值,即為枝條導水面積與葉片總面積的比值。
1.3.6 滲透調節物質和非結構性碳水化合物
采集當年新生的同化枝,帶回實驗室。將樣品置于65℃烘箱烘干48h后,使用混合震蕩型研磨儀研磨成粉。測定指標包括鈉離子(Na+)、氯離子(Cl-)、脯氨酸(Pro)、可溶性糖(SS)和淀粉。Na+采用火焰光度計(FP640,上海精科)測定;Cl-采用以明膠-乙醇水溶液為穩定劑的氯化銀沉淀法測定[33];脯氨酸采用茚三酮比色法測定[34];可溶性糖和淀粉采用苯酚硫酸法測定[35],非結構性碳水化合物為可溶性糖和淀粉之和。每個指標的測定均為3個重復。
1.4 數據分析
利用SPSS 21.0軟件對數據進行統計分析, 利用單因素方差分析(One-way ANOVA)中的Duncan檢驗分析不同地下水埋深生理生態參數的顯著性差異,利用雙因素方差分析(Two-way ANOVA)確定地下水埋深、生長階段以及地下水埋深×生長階段的交互作用對梭梭生理生態參數的影響;利用獨立樣本T檢驗分析同一地下水埋深不同生長階段生理生態參數的顯著性差異;采用Pearson相關分析探討地下水埋深對梭梭同化枝生長速率的影響。數據分析之前對所有數據進行正態性和方差齊性檢驗。使用SigmaPlot 12.5軟件對數據進行繪圖。
2 結果
2.1 不同地下水埋深和生長期梭梭生理生化特征的變化
不同生長階段梭梭沿地下水埋深增加的水分關系如圖2所示。地下水埋深和生長階段對Ψpd和Ψm有顯著的影響(P<0.001)。Ψpd和Ψm隨地下水埋深的變化與生長階段有關。生長季前期Ψpd和Ψm顯著高于生長季后期(P<0.05)。隨著地下水埋深的增加,生長季前期,S1梭梭Ψpd最高,明顯高于S2—S4(P<0.05);S1和S3Ψm明顯高于其它3個樣點(P<0.05);生長季后期,梭梭Ψpd先降后升,Ψm在較淺的地下水埋深明顯高于較深的地下水埋深(P<0.05)。地下水埋深對ABWC產生了顯著的影響(P<0.001)。生長季前期和后期,梭梭ABWC先升后降。

圖2 不同地下水埋深下梭梭水分關系的變化
不同生長階段梭梭同化枝中滲透調節物質和非結構性碳水化合物沿地下水埋深梯度的變化特征如圖3所示。地下水埋深對梭梭同化枝中Cl-、SS和碳水化合物(NSC)含量的影響達到極顯著水平(P<0.01)。隨著地下水埋深的增加,梭梭同化枝中Cl-的含量先升后降;S1—S4同化枝中SS和NSC含量差異不顯著, 但在S5 SS和NSC含量顯著升高(P<0.05)。生長階段對梭梭同化枝中Na+和SS含量產生了極顯著的影響(P<0.01)。生長季前期梭梭同化枝中Na+含量顯著低于生長季后期(P<0.05)。地下水埋深對梭梭同化枝中Pro和淀粉含量的影響與生長階段有關(P<0.05)。生長季前期,S3梭梭同化枝中脯氨酸(Pro)含量最高,且明顯高于其它4個樣點(P<0.05); S1—S4同化枝中淀粉含量差異不顯著,但在S5淀粉含量明顯減少(P<0.05)。生長季后期,S4梭梭同化枝中Pro含量最低,另4個樣點Pro含量明顯高于S4(P<0.05),且不存在明顯差異;同化枝中淀粉含量相對穩定。
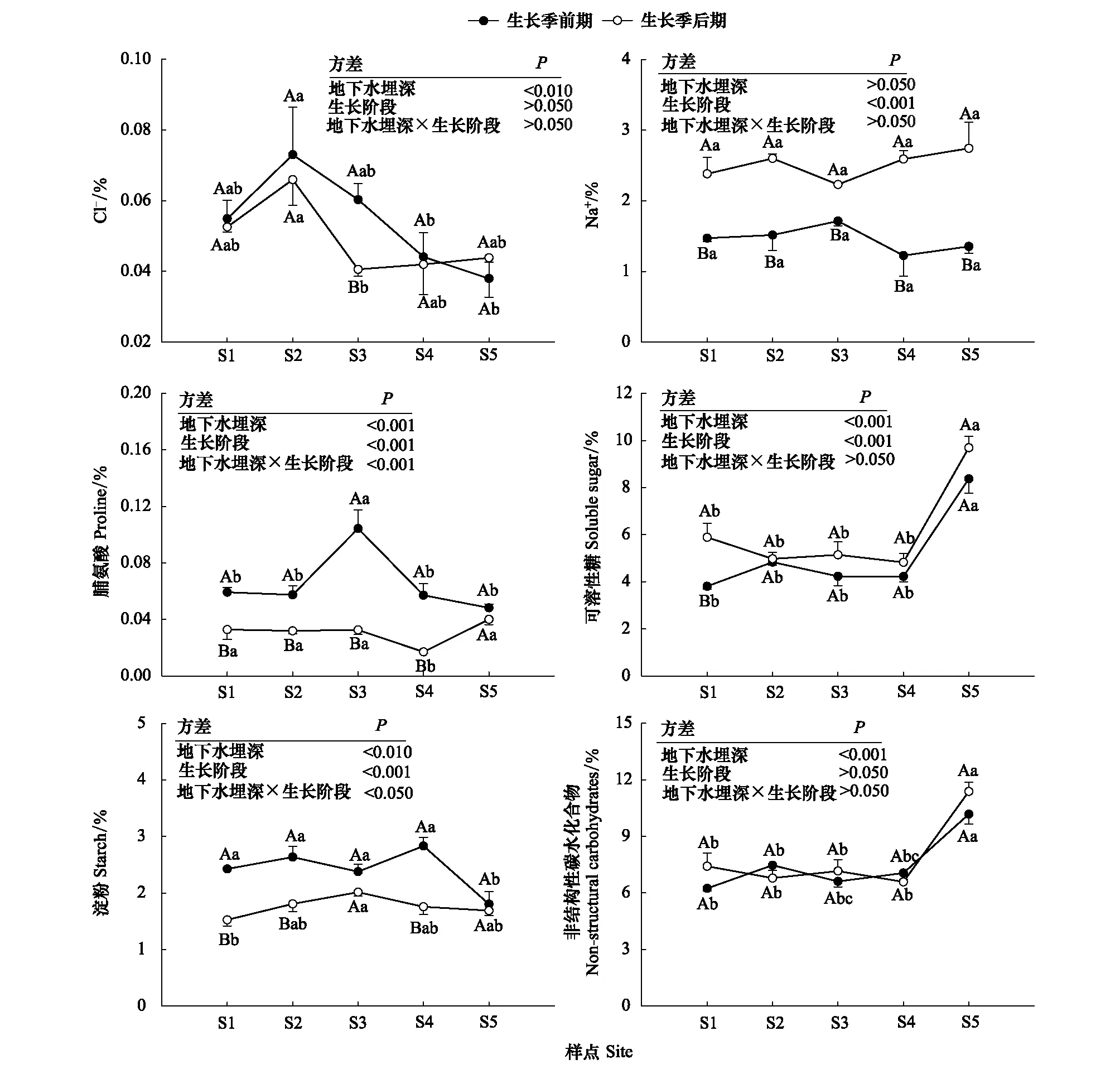
圖3 不同地下水埋深下梭梭同化枝滲透調節物質和非結構性碳水化合物的變化
2.2 不同地下水埋深和生長期梭梭生長和形態的變化
不同地下水埋深下梭梭的同化枝生長速率如圖4所示。地下水埋深對4—5月梭梭ABGR產生了顯著的影響,5月地下水埋深較淺的ABGR明顯高于地下水埋深較深的地點(P<0.05)。6—8月地下水埋深對ABGR沒有顯著的影響。不同地下水埋深梭梭胡伯爾值的變化特征如圖5所示。地下水埋深、生長階段和二者的交互作用對梭梭HV的影響均達到極顯著水平(P≤0.001)。生長季前期,在較淺的地下水埋深HV差異不顯著,在較深的地下水埋深HV顯著增大(P<0.05)。生長季后期,在地下水埋深增加的條件下,梭梭HV相對穩定。
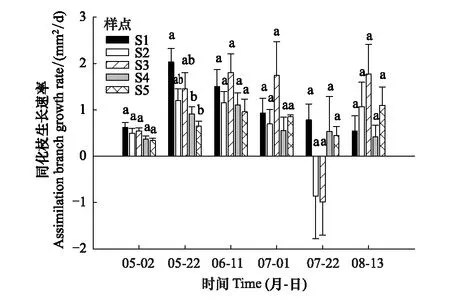
圖4 不同地下水埋下深梭梭同化枝生長速率
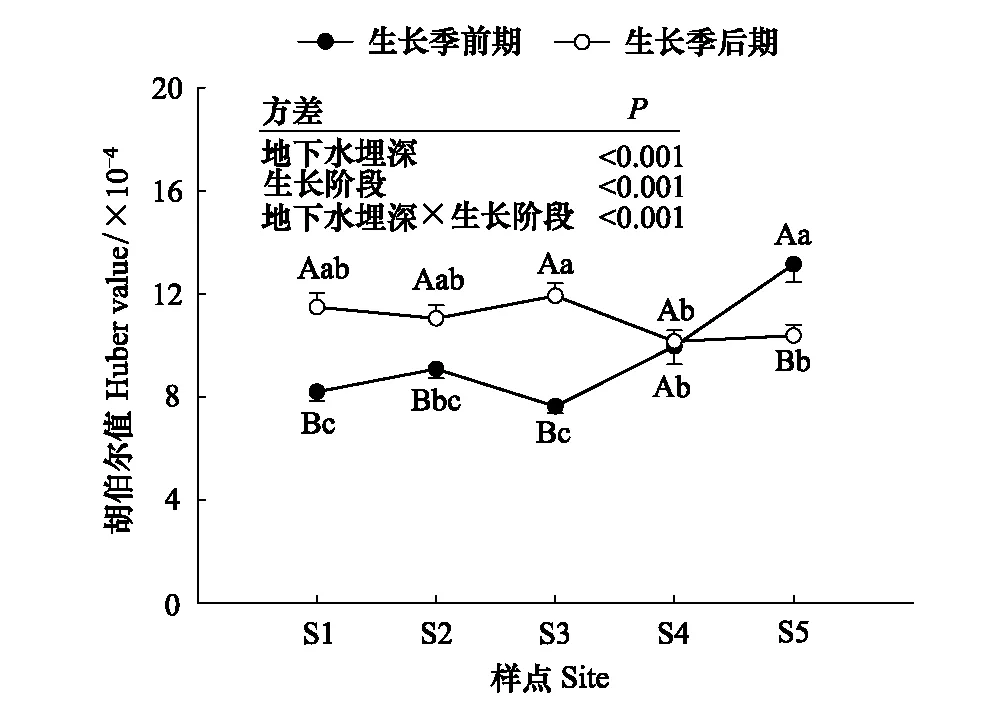
圖5 不同地下水埋深下梭梭胡伯爾值的變化
3 討論
3.1 地下水埋深和季節性干旱對梭梭生理性狀的影響
葉(同化枝)水勢和含水量不僅能反映植物的水分狀態,也是植物進行水分調節的重要表征。馮樹林等[36]研究發現,隨著干旱脅迫的加劇,荒漠灌木葉水勢呈下降趨勢。然而,也有學者認為,荒漠灌木的水力行為高度動態,在不同的水分供應狀態和溫度條件下,其水力調節對策可從部分等水勢轉變為極端非等水勢[13]。本研究中,隨著地下水埋深的增加,生長季前期和后期梭梭Ψpd均呈現先降低后升高的變化趨勢(圖2),表明梭梭Ψpd受到地下水埋深增加引起的水文干旱脅迫,但地下水有效性降低促使梭梭水分利用效率提高、氣孔導度降低以及蒸騰速率下降[11—12],這在一定程度上補償了地下水埋深持續增加對其同化枝水勢的負面影響。與較淺的地下水埋深相比,較深的地下水埋深下Ψm明顯降低(圖2),這與前人[11]研究的結果一致。此外,與生長季前期相比,除S2的Ψm外,生長季后期Ψpd和Ψm均顯著降低(圖2),這表明梭梭經歷季節性大氣干旱。梭梭是荒漠環境中的旱生肉質植物,同化枝角質層厚,儲水組織發育良好[37]。因此,盡管地下水埋深在15 m以上,生長季前期和后期梭梭ABWC仍高于65%以上(圖2),這表明高ABWC是梭梭在干旱生境中長期生存的適應性特征。
滲透調節是植物在細胞水平上減輕干旱脅迫的一種適應策略。滲透調節物質以不同的方式保護植物免受脅迫,如提高細胞的滲透調節能力和保護膜的穩定性。植物在生長過程中對氯的需求量十分少,一般在0.03%—2.0%之間。梭梭同化枝中Cl-含量低,且隨地下水埋深的增加先升后降(圖3),這表明梭梭同化枝中Cl-的滲透調節能力有限[38],當水分有效性降低到一定程度時,植物體內Cl-向地上部分的遷移會減弱[39]。在模擬干旱的溫室條件[40]和極端干旱的自然環境中[20], Na+對梭梭滲透勢調節的貢獻均高達45%—50%。本文中梭梭同化枝中Na+的含量沒有受到地下水埋深的限制,而是因生長階段而異。在極端干旱的荒漠環境中,Na+是梭梭長期適應干旱的重要滲透調節物質,其含量隨干旱脅迫時間的延長而累積。梭梭同化枝中Pro含量在多個樣點差異均不顯著(圖3),說明梭梭同化枝中Pro對滲透調節的貢獻較小且有限。呂昕培等[40]在研究梭梭幼苗應對干旱脅迫的滲透調節作用時,也發現Pro對滲透調節的貢獻水平十分有限。
有研究發現,在滲透脅迫下,梭梭幼苗同化枝中的SS對滲透調節的貢獻較小[40]。但王國華等[38]研究表明,SS是種植年限為5—20年的梭梭抵御干旱脅迫的重要滲透調節物質。樹齡的增長促使梭梭體內非結構性碳水化合物(NSC)含量成倍提高[24]。在本試驗中,生長季前期與后期梭梭同化枝中SS含量較高,且在S5顯著升高(圖3)。由此表明,SS是成年梭梭抵御水文干旱的滲透調節物質之一。淀粉作為植物的儲存物質,通常在植物遭受干旱時部分轉化為SS以供植物應對環境脅迫[41]。這與本研究結果一致,生長季前期梭梭同化枝中淀粉含量在S1-S4相對穩定,但在S5顯著下降(圖3)。而且,與生長季前期相比,生長季后期同化枝中淀粉含量出現明顯的損耗(圖3)。這表明梭梭可以將同化枝中的淀粉轉化為SS以抵御地下水埋深增加的水文干旱和季節性大氣干旱。干旱會改變樹木的碳平衡,NSC是在干旱脅迫下衡量碳平衡的重要性狀[42]。有研究表明,干旱脅迫下植物NSC含量減少[43],但NSC含量累積和維持[44]的結論也有報道。本文中,地下水埋深持續增加引起的水文干旱沒有導致梭梭NSC含量耗盡,反而顯著升高(圖3)。這意味著梭梭沒有受到碳限制,在干旱脅迫下維持較高的NSC含量,這對其水力功能恢復可能十分重要[45]。
3.2 地下水埋深和季節性干旱對梭梭生長和形態的影響
植物生長對水分的變化非常敏感,尤其是荒漠植被。地下水埋深增加引起的水文干旱會改變植物的表型可塑性[46]。有研究表明,干旱脅迫會造成荒漠植物生長緩慢、生物量減少、總葉面積和比葉面積減小[47—48]。早期研究發現,夏季降水的減少會限制梭梭的生長[25]。本研究中,相關性分析表明,4—5月梭梭ABGR與地下水埋深呈顯著負相關(P<0.05)。蔣菊芳[49]、陳玲[50]等研究表明,梭梭萌芽期為4月12日,展葉期為4月30日,新枝形成期為5月22日。這意味著在新枝形成期,地下水埋深增加會導致ABGR顯著降低。由此可見,梭梭可以通過改變同化枝生長速率來響應地下水水文干旱。研究結果與彭守蘭等[14]的結論一致,他們發現疏葉駱駝刺隨著潛水埋深的增加,生長緩慢且生物量累積減少,并且認為這種形態的可塑性是荒漠灌木長期適應干旱貧瘠環境的生態策略。
葉片的形態和結構與植物的抗旱機制密切相關,胡伯爾值(HV)被認為是評價植物抗旱性的重要功能性狀[51]。在干旱脅迫下,植物通常具有較高的HV以維持葉片水分關系的穩定[52]。在阿塔卡馬沙漠牧豆樹(Prosopistamarugo)[53]和塔克拉瑪干沙漠胡楊[15]的研究中表明,地下水埋深增加會促使HV顯著增大。這與本研究結果一致,生長季前期梭梭HV隨地下水埋深的增加呈現增大的趨勢(圖5),這增強了單位葉面積的供水能力;生長季后期梭梭采取有效的形態調整,維持較高的HV值以保證水分運輸的有效性(圖5)。
4 結論
地下水埋深、生長階段及其交互作用對古爾班通古特沙漠梭梭的生理生化和生長與形態產生了不同的影響。主要結論為:(1)在生理調節上,梭梭通過先降低后升高Ψpd和積累NSC響應地下水埋深的增加。(2)在生長和形態調整上,梭梭在生長季前期采取降低新枝形成期的ABGR和增大HV的策略響應地下水埋深的增加。(3)與生長季前期相比,梭梭采取降低Ψpd、維持較高的HV、積累Na+和消耗淀粉的策略抵御季節性干旱。(4)在地下水埋深和生長階段的交互作用下,降低Ψm、維持較高的ABWC和積累SS是梭梭適應大氣和地下水水文干旱的結果。由此可見,梭梭生理和形態性狀對地下水水文干旱和季節性大氣干旱及其交互作用間的響應,既存在一致性,又存在差異性。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
