發酵乳桿菌CECT 5716產胞外多糖培養基成分優化及抗氧化活性研究
2022-12-11 08:12:56李佳佳耿偉濤賈龍剛陳鐵濤王艷萍
中國釀造 2022年11期
方 偉,李佳佳,耿偉濤,賈龍剛,陳鐵濤,王艷萍*
(1.天津科技大學 食品科學與工程學院,天津 300457;2.百施(上海)生物科技有限公司,上海 200436)
乳酸菌(lactic acid bacteria,LAB)是一類能利用碳水化合物產生大量乳酸的革蘭氏陽性細菌的統稱,廣泛存在于人和動物的腸道以及傳統發酵制品等中[1-2]。發酵乳桿菌(Lactobacillus fermentum)CECT 5716是一株分離自健康人乳的乳酸菌,具有極高的安全性[3]。2016年國家衛生和計劃生育委員會發布公告(2016年第6號),將發酵乳桿菌CECT 5716列入《可用于嬰幼兒食品的菌種名單》,為該菌株在母嬰產品中的應用提供了依據。目前,國內外研究學者已對發酵乳桿菌CECT 5716的益生功能開展了大量的研究工作,研究發現,該菌株具有緩解哺乳期婦女乳腺炎[4]、免疫調節[5]、降血壓和降血脂[6]等益生功能。
乳酸菌胞外多糖(exopolysaccharides,EPS)是乳酸菌在生長代謝過程中分泌到細胞外的高分子質量碳水化合物聚合物[7]。產EPS的乳酸菌主要有乳桿菌屬(Lactobacillus)、明串珠菌屬(Trichococcus)和鏈球菌屬(Streptococcus)等[8]。國內外相關研究表明,EPS除具有增黏性、保水性和抗剪切性等理化特性外,還具有多種生理活性,如抗腫瘤、調節腸道菌群和降低血清膽固醇等[9-10]。而發酵乳桿菌CECT 5716擁有的益生功能是否與其EPS相關,其作用的關鍵分子尚需進行深入研究與揭示,但目前對其EPS的系統研究鮮見報道,因此迫切需要對其EPS展開研究,而較低的產量是其研究受限的關鍵因素。
本研究采用單因素及正交試驗優化了發酵乳桿菌CECT 5716產EPS的培養基成分,并對優化后獲得的EPS進行抗氧化活性研究,以期為該菌株EPS的大規模制備及開發其作為天然抗氧化劑提供參考。
1 材料與方法
1.1 材料與試劑
1.1.1 菌種
發酵乳桿菌(Lactobacillus fermentum)CECT 5716:百施(上海)生物科技有限公司保藏。
1.1.2 培養基
MRS液體培養基[11]:牛肉膏1.0%,酵母膏0.5%,蛋白胨1.0%,葡萄糖2.0%,磷酸氫二鉀0.2%,無水乙酸鈉0.5%,硫酸鎂0.02%,硫酸錳0.005%,檸檬酸氫二胺0.2%,吐溫80 0.1%,pH值6.2,115 ℃高壓蒸汽滅菌15 min。固體MRS培養基中添加瓊脂2%。
1.1.3 試劑
無水乙醇、三氯乙酸、硫酸、苯酚(重蒸餾)、鉬酸銨、硫酸鈉、鐵氰化鉀、三氯化鐵、過硫酸鉀、維生素C(vitamin C,VC)(均為分析純):江天化工技術股份有限公司;透析袋(截留分子質量8 000 Da):鼎國生物技術有限責任公司;1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-trinitrophenylhy drazyl,DPPH)、2,2-聯氮-二(3-乙基-苯并噻唑啉-6-磺酸)二銨鹽(2,2'-Azino-bis(3-ethyl-benzothiazoline-6-sulfonic acid)diammonium salt,ABTS):國藥集團化學試劑有限公司;其他試劑均為國產分析純。
1.2 儀器與設備
UV-9600紫外可見分光光度計:北京瑞利分析儀器有限公司;Avanti J-E大容量冷凍離心機:美國Beckman公司;ALPHA 1-2 LD冷凍干燥機:德國CHRIST公司;Anaerobox IV型厭氧培養箱:美國GeneScience公司;Thermo全波長酶標儀:美國Thermo Fisher公司。
1.3 試驗方法
1.3.1 發酵乳桿菌CECT 5716的培養條件
將保藏于-80 ℃的發酵乳桿菌CECT 5716劃線接種于MRS固體培養基上,37 ℃靜置培養48 h,挑取單菌落,再重復進行2次平板劃線,挑取單菌落接種于MRS液體培養基中,37℃靜置培養12 h,并傳代兩次以使菌株的活力最大化。將活化后的菌株以2%(V/V)的接種量接種于MRS液體培養基中,37 ℃靜置培養16 h。
1.3.2 發酵乳桿菌CECT 5716產EPS培養基優化單因素試驗
碳源種類、復配比例及其添加量的優化:以MRS液體培養基(對照)為基礎,使用不同的碳源(麥芽糖、蔗糖、乳糖和果糖)替代MRS培養基中的碳源(葡萄糖),以確定最佳碳源的種類。在最佳碳源種類的基礎上,對麥芽糖和蔗糖進行了復配,復配比例為3∶1、2∶1、1∶1、1∶2、1∶3。在最佳復配比(麥芽糖∶蔗糖=2∶1)的基礎上進行碳源添加量(1.00%、1.50%、2.00%、2.50%、3.00%、3.50%、4.00%)的優化。
氮源種類及其添加量的優化:以MRS液體培養基(對照)為基礎,使用不同氮源(蛋白胨、牛肉膏、酵母膏、胰蛋白胨、3072乳清蛋白水解物和3065乳清蛋白水解物)替代MRS培養基中的氮源(蛋白胨、牛肉膏和酵母膏),以確定最佳氮源的種類。在最佳氮源種類的基礎上進行氮源添加量(2.00%、2.50%、3.00%、3.50%、4.00%、4.50%)的優化。
L-半胱氨酸鹽酸鹽添加量的優化:以MRS液體培養基(對照)為基礎,考察L-半胱氨酸鹽酸鹽不同添加量(0.05%、0.10%、0.15%、0.20%、0.25%、0.30%)對發酵乳桿菌CECT 5716 EPS產量的影響。
1.3.3 發酵乳桿菌CECT 5716產EPS培養基優化正交試驗
根據單因素試驗結果,篩選出顯著影響EPS產量的3個因素復合碳源(麥芽糖∶蔗糖=2∶1)添加量(A)、酵母膏添加量(B)、L-半胱氨酸鹽酸鹽添加量(C)。以EPS產量(Y)為考察指標,設計3因素3水平L9(33)正交試驗,優化培養基組成。正交試驗因素與水平見表1。
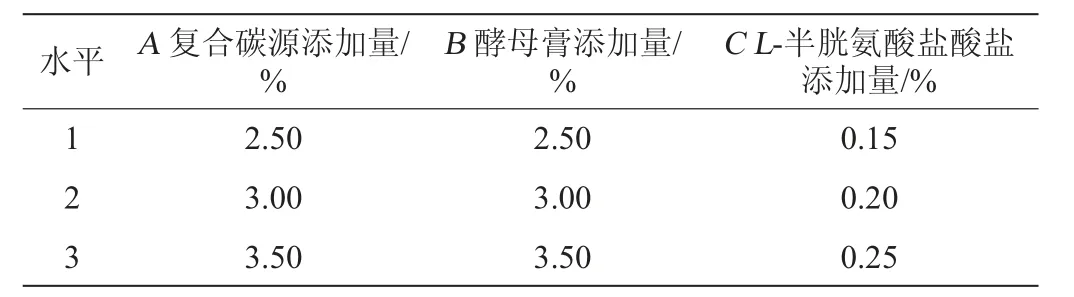
表1 發酵乳桿菌CECT 5716產胞外多糖培養基優化正交試驗因素與水平Table 1 Factors and levels of orthogonal experiments for medium optimization of exopolysaccharide production by Lactobacillus fermentum CECT 5716
1.3.4 EPS的制備及含量的測定
根據BENGOA A A等[12]方法并略作修改制備EPS。取發酵液在100 ℃加熱30 min,冷卻后離心(4 ℃、8 000 r/min、20 min)。取上清,加入2倍體積的無水乙醇,4 ℃靜置過夜后,離心(4 ℃、8 000 r/min、20 min)。將沉淀復溶于適量蒸餾水中,加入10%三氯乙酸,置于磁力攪拌器上冰浴4 h,離心(4 ℃、8 000 r/min、20 min),取上清,將其pH值調至7.0,加2倍體積的無水乙醇進行復醇沉,4 ℃靜置過夜后離心(4 ℃、8 000 r/min、20 min),沉淀復溶于少量蒸餾水中,透析(截留分子質量8 000 Da)2~3 d,定容,采用苯酚-硫酸法[13]測定透析液中EPS的含量,將透析液冷凍干燥得EPS。
1.3.5 EPS純度的測定
精確稱取100.00 mg EPS,用適量蒸餾水溶解,轉移至100 mL容量瓶中定容,混勻,得樣品溶液,采用苯酚-硫酸法[13]測定樣品中的多糖含量,計算EPS的純度,其計算公式如下:
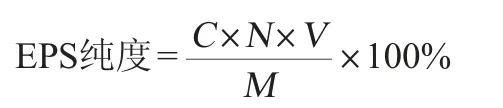
式中:C為稀釋后樣品的多糖含量,mg/L;N為稀釋倍數;V為樣品溶液的體積,L;M為樣品的質量,mg。
1.3.6 EPS的體外抗氧化活性測定
DPPH自由基清除活性的測定:參考ZHANG Q等[14]的方法;ABTS自由基清除活性的測定:參考XU Y M等[15]的方法;總抗氧化能力的測定:參考SHANKAR T等[16]的方法;Fe3+總還原力的測定:參考WANG K等[17]的方法。
1.3.7 數據處理
使用Graphpad Prism 8.0軟件進行數據分析和繪圖。所有試驗均重復3次,結果表示為“平均值±標準差”。
2 結果與分析
2.1 發酵乳桿菌CECT 5716產EPS培養基優化單因素試驗結果
2.1.1 碳源種類、復配比例及添加量的優化
碳源是微生物生長代謝過程中的必需物質,對乳酸菌EPS的產量及其相對分子質量、單糖組成和摩爾比都有較大的影響[18]。不同碳源對EPS產量的影響見圖1A。由圖1A可知,當碳源為麥芽糖時,EPS的產量最高,為(305.67±9.71)mg/L,是對照組的1.25倍;其余依次為蔗糖、乳糖和果糖。不同碳源導致EPS產量的差異,可能是因為該菌對各種碳源的利用能力不同,導致菌株代謝速率等方面的差異,從側面也反映出EPS合成途徑的調節可能依賴于碳水化合物的不同[19]。
綜合考慮EPS產量與降低成本的因素,選擇將麥芽糖和蔗糖進行復配作為碳源,考察其復配比例對EPS產量的影響,結果見圖1B。由圖1B可知,當麥芽糖∶蔗糖=2∶1時,EPS的產量最高,為(364.65±13.89)mg/L。因此,選用麥芽糖和蔗糖以2∶1的比例復配作為碳源。
碳源濃度過低或過高都不利于菌體的正常生長及其代謝產物的合成,故碳源添加量對EPS的產量也有顯著影響[20]。不同復合碳源添加量對EPS產量的影響,結果見圖1C。由圖1C可知,隨著復合碳源添加量的升高,EPS產量呈現先上升后下降的趨勢。當復合碳源添加量為3%時,EPS產量最高,為(421.11±6.78)mg/L。復合碳源添加量過高導致EPS產量下降,分析原因可能是由于發酵過程中產生了“葡萄糖效應”,導致菌株代謝異常,乳酸等初級代謝產物大量積累,次級代謝受到抑制從而影響了EPS的合成[8]。因此,確定最優復合碳源添加量為3%。
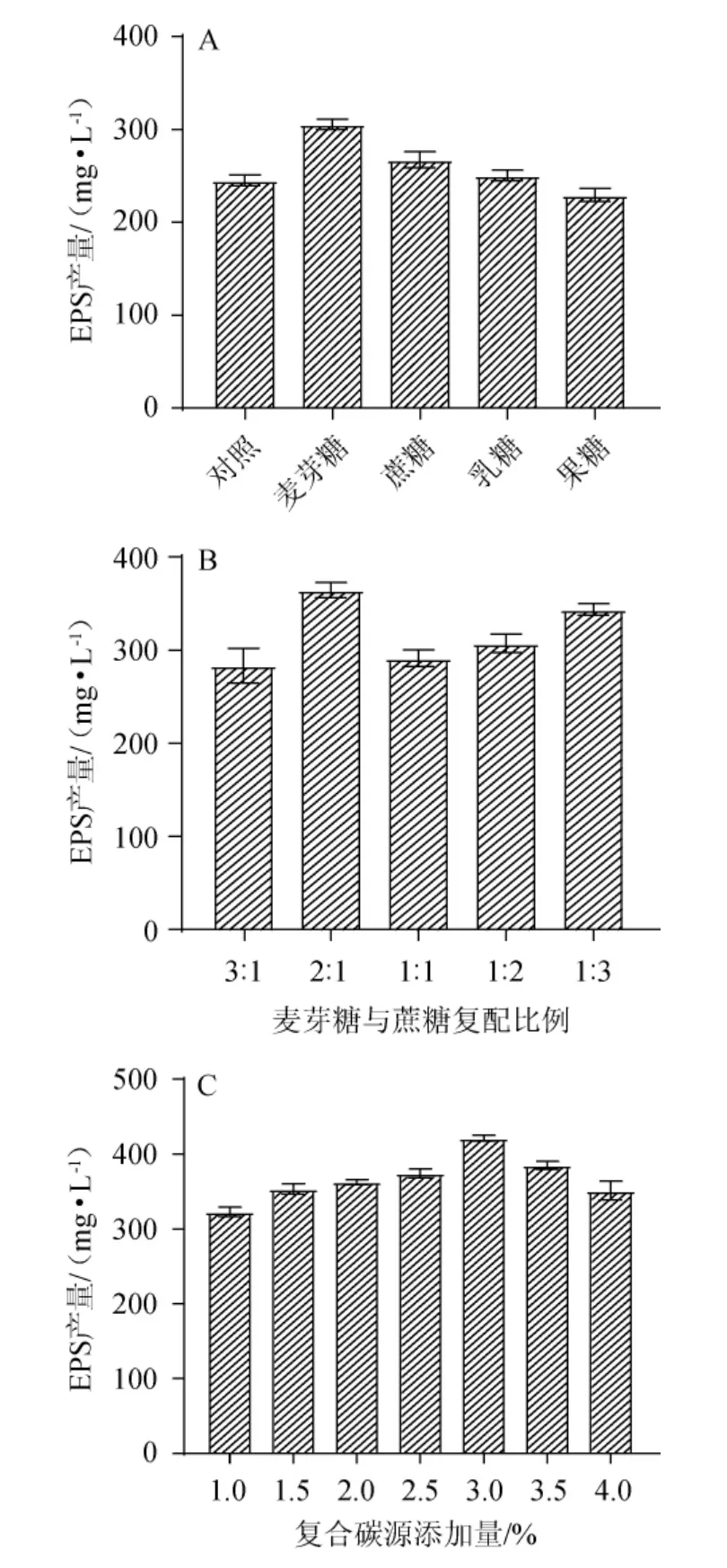
圖1 碳源種類(A)、復配比例(B)及復合碳源添加量(C)對發酵乳桿菌CECT 5716產胞外多糖的影響Fig.1 Effects of carbon source type(A),compound proportion(B)and compound carbon source addition(C)on exopolysaccharide production by Lactobacillus fermentum CECT 5716
2.1.2 氮源種類及添加量的優化
氮源在乳酸菌的生長及其代謝產物的合成中,發揮著重要的作用[21]。不同氮源對EPS產量的影響見圖2A。由圖2A可知,不同氮源對EPS產量有較大的影響,當氮源為酵母膏時,EPS產量最高,為(740.15±19.64)mg/L,是對照組的3.02倍,顯著高于其他氮源。這可能是因為酵母膏富含蛋白質、氨基酸和維生素,可被菌體更好地利用[22]。因此,確定最優氮源為酵母膏。酵母膏添加量對發酵乳桿菌CECT 5716 EPS產量的影響見圖2B。由圖2B可知,酵母膏添加量對發酵乳桿菌CECT5716 EPS產量的影響顯著,隨著酵母膏添加量的升高,EPS產量呈先升高后下降的趨勢,這可能是由于氮源可作為能源物質而被菌株利用,促進其代謝并合成EPS,而氮源添加量過高,導致菌株產生過量的銨、硝態氮,從而抑制了其EPS的積累[23]。當酵母膏添加量為3%時,EPS產量最高,為(1 077.68±20.67)mg/L。因此,確定酵母膏最優添加量為3%。
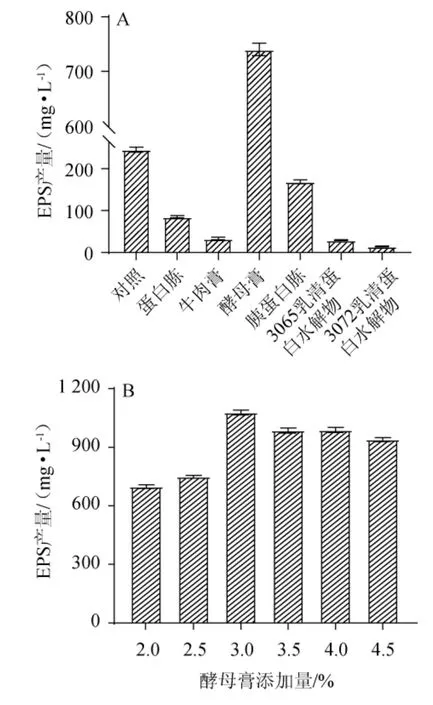
圖2 氮源種類(A)及酵母膏添加量(B)對發酵乳桿菌CECT 5716產胞外多糖的影響Fig.2 Effects of nitrogen source type (A) and yeast extract addition(B) on exopolysaccharide production by Lactobacillus fermentum CECT 5716
2.1.3L-半胱氨酸鹽酸鹽添加量的優化
L-半胱氨酸鹽酸鹽常作為一種還原劑加入培養基中,使培養基保持一個相對厭氧的環境[24]。L-半胱氨酸鹽酸鹽不同添加量對EPS產量的影響見圖3。
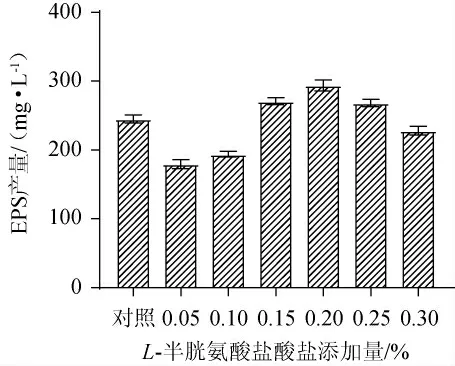
圖3 L-半胱氨酸鹽酸鹽添加量對發酵乳桿菌CECT 5716產胞外多糖的影響Fig.3 Effects of L-cysteine hydrochloride addition on exopolysaccharide production by Lactobacillus fermentum CECT 5716
由圖3可知,與對照組相比,添加一定量的L-半胱氨酸鹽酸鹽可以提高EPS的產量,當L-半胱氨酸鹽酸鹽添加量為0.2%時,EPS產量最高,為(293.81±13.79)mg/L,是對照組的1.20倍。這可能與其能夠改變培養基的氧化還原電位,從而更有利于發酵乳桿菌CECT 5716的生長及代謝有關[25]。因此,確定L-半胱氨酸鹽酸鹽最優添加量為0.2%。
2.2 發酵乳桿菌CECT 5716產EPS培養基優化正交試驗結果
在單因素試驗的基礎上,進一步通過正交試驗優化培養基組成,正交試驗設計及結果見表2。由表2可知,通過極差分析,L-半胱氨酸鹽酸鹽是影響EPS產量的最主要因素,麥芽糖和蔗糖添加量次之,酵母膏添加量影響最小。最佳培養基配方組合為A3B3C2,因為在A3與A2以及B3與B1水平下,EPS的產量相差不大,考慮到生產成本等因素,最終選擇A2B1C2為最佳組合,即碳源添加量3%、酵母膏2.5%、L-半胱氨酸鹽酸鹽0.2%。在此條件下,EPS產量為(1 560.15±22.64)mg/L。驗證試驗得EPS產量為(1 575±22.91)mg/L,證明該正交試驗設計成功。
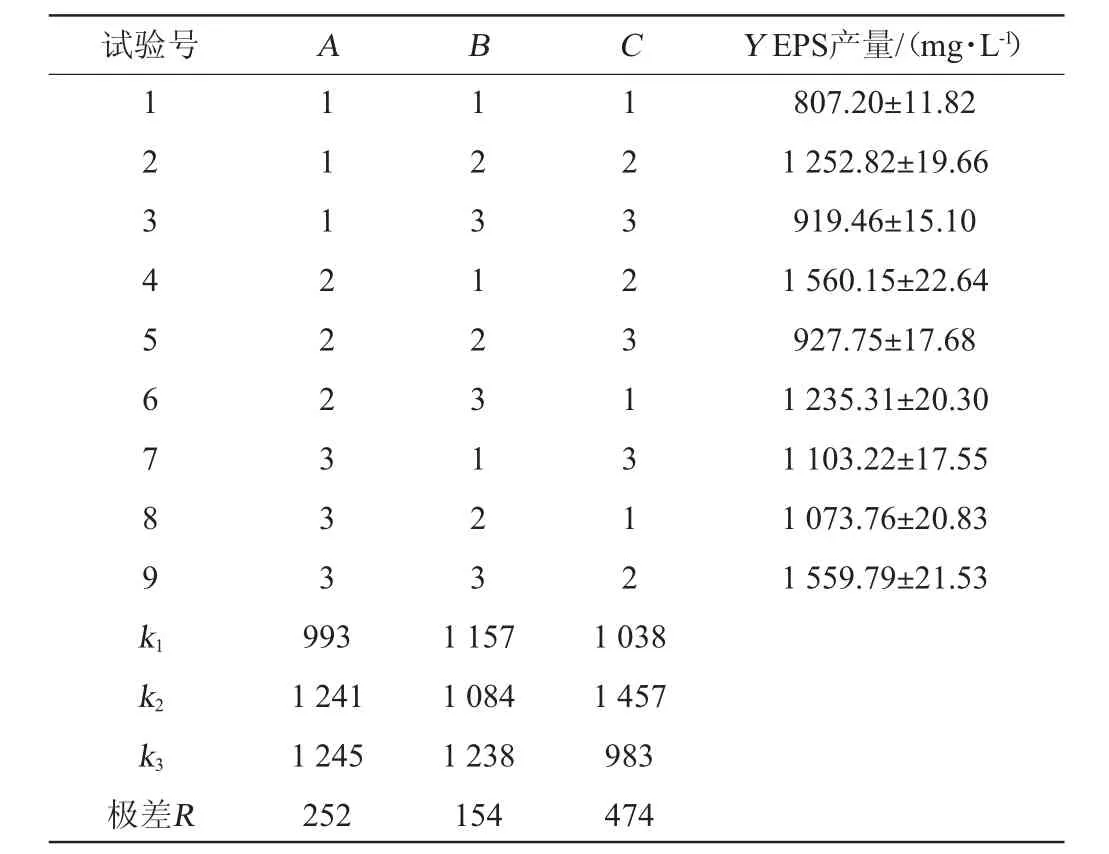
表2 發酵乳桿菌CECT 5716產EPS培養基優化正交試驗結果與分析Table 2 Results and analysis of orthogonal experiments for medium optimization of exopolysaccharide production by Lactobacillus fermentum CECT 5716
2.3 胞外多糖的純度
經測定,優化后所得EPS的純度為(92.2±1.8)%。
2.4 胞外多糖的抗氧化活性
2.4.1 DPPH自由基清除活性
DPPH自由基可與自由基清除劑(如EPS、多酚和維生素E等)反應,其溶液吸光度值的變化可用于評估自由基清除劑的抗氧化能力[26]。不同質量濃度的EPS對DPPH自由基的清除能力見圖4。由圖4可知,當質量濃度為0~8 mg/mL時,EPS和VC對DPPH自由基的清除能力隨著質量濃度的增加而增強,但EPS對DPPH自由基的清除能力低于相同質量濃度的VC,EPS對DPPH自由基的半抑制濃度(50%inhibition concentration,IC50)值為0.24 mg/mL。當EPS質量濃度為8 mg/mL時,EPS、VC對DPPH自由基的清除率分別為(84.17±1.30)%、(89.62±1.55)%。EPS清除自由基的機制可能是由于EPS與自由基結合并形成了穩定的分子條件,從而終止了自由基的進一步反應[27]。
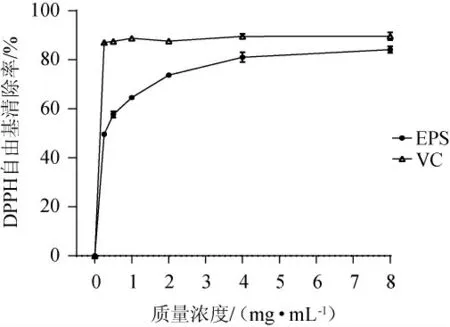
圖4 胞外多糖對DPPH自由基的清除活性Fig.4 Scavenging activity of exopolysaccharide on DPPH radical
2.4.2 ABTS自由基清除活性
不同質量濃度的EPS對ABTS自由基的清除能力見圖5。由圖5可知,當質量濃度為0~8 mg/mL,EPS和VC對ABTS自由基的清除能力隨著質量濃度的增加而增強,但EPS對ABTS自由基的清除能力低于相同質量濃度的VC的清除能力,EPS對ABTS自由基的IC50值為14.81 mg/mL。當EPS的質量濃度為8 mg/mL時,EPS對ABTS自由基的清除率為(35.37±1.24)%。EPS具有較高的ABTS自由基清除能力可能是由于其能夠將自由基由活性狀態轉化為穩定狀態,并通過向自由基提供電子來終止自由基鏈式反應[28]。
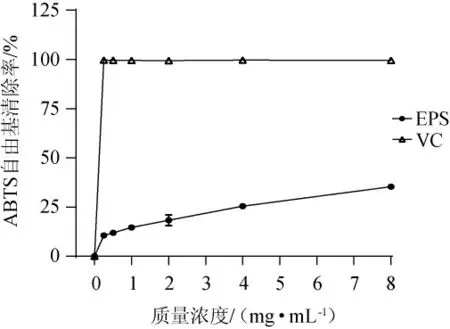
圖5 胞外多糖對ABTS自由基的清除活性Fig.5 Scavenging activity of exopolysaccharide on ABTS radical
2.4.3 總抗氧化能力
總抗氧化能力反映的是非酶促抗氧化的能力[29],不同質量濃度的EPS的總抗氧化能力見圖6。由圖6可知,當質量濃度為0~8 mg/mL時,EPS和VC的總抗氧化能力隨著其質量濃度的增加而增強,且在同一質量濃度下,EPS的總抗氧化能力低于VC,當EPS質量濃度為8 mg/mL時,溶液的吸光度值為0.31±0.004。
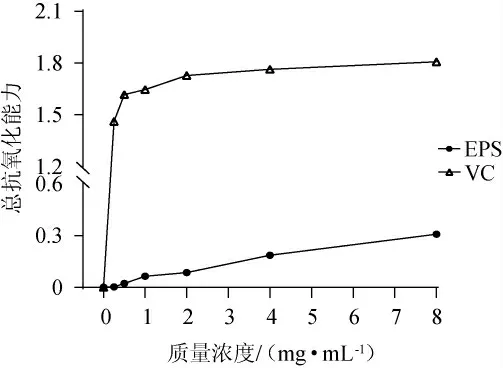
圖6 胞外多糖的總抗氧化能力Fig.6 Total antioxidant capacity of exopolysaccharide
2.4.4 Fe3+總還原力
天然活性物質的鐵還原能力可作為其電子給體活性的重要指標,常用來評價抗氧化劑的抗氧化能力[30]。為了解EPS將Fe3+還原為其亞鐵形式的能力[31],測定不同質量濃度EPS的Fe3+總還原力,結果見圖7。由圖7可知,當質量濃度為0~8 mg/mL時,EPS和VC的Fe3+總還原力隨著其質量濃度的增加而增強,且在同一質量濃度下,EPS的Fe3+總還原力低于VC。當EPS的質量濃度為8 mg/mL時,EPS的Fe3+總還原力為0.55±0.01。
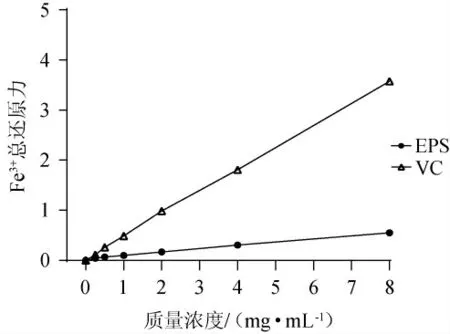
圖7 胞外多糖的Fe3+總還原力Fig.7 Fe3+total reducing power of exopolysaccharide
綜上結果表明,該EPS具有一定的抗氧化能力,抗氧化效果與其質量濃度呈正相關。但EPS在各種反應體系中顯示出不同的抗氧化能力,可能是由于EPS的單糖組成、羥基含量以及供氫能力不同等因素導致的[32]。
3 結論
通過單因素和正交試驗優化得到發酵乳桿菌CECT 5716產EPS的最優培養基組成為:麥芽糖2.0%、蔗糖1.0%、酵母膏2.5%、L-半胱氨酸鹽酸鹽0.2%、乙酸鈉0.5%、磷酸氫二鉀0.2%、硫酸鎂0.02%、硫酸錳0.005%、檸檬酸氫二胺0.2%、吐溫80 0.1%。在此條件下,EPS產量達到(1 575±22.91)mg/L,是優化前的6.38倍。EPS的抗氧化活性與其質量濃度成正比,當EPS的質量濃度為8 mg/mL時,對DPPH自由基的清除率為(84.17±1.30)%、對ABTS自由基的清除率為(35.37±1.24)%、總抗氧化能力為0.31±0.01和Fe3+總還原力為0.55±0.01。本研究優化了發酵乳桿菌CECT 5716產EPS的培養基成分,并研究了該菌株產生的EPS的抗氧化特性,為進一步揭示發酵乳桿菌CECT 5716抗氧化的功能及作用機制的研究奠定了基礎,也為發酵乳桿菌CECT 5716的大規模制備及其結構的研究提供了理論依據。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國生殖健康(2019年2期)2019-08-23 08:12:08
聚氯乙烯(2018年9期)2018-02-18 01:11:34
產品可靠性報告(2017年7期)2017-09-05 09:49:12
