“雙碳”政策背景下厭氧消化產氫產乙酸過程控制的研究進展*
2022-12-13 20:15:55萬思卓袁林江
環境污染與防治 2022年10期
丁 聰 萬思卓 王 茹 袁林江
(西安建筑科技大學環境與市政工程學院,陜西 西安 710055)
能源是國民經濟的重要物質基礎,因此能源的開發和有效利用至關重要,當前能源緊缺問題已成為各個國家和社會關注的重點問題[1]。隨著經濟社會的發展,人們生活水平的逐漸提高,生產生活中產生大量的生活及工業廢棄物,這些廢棄物的高效處理成為亟待解決的問題。厭氧消化技術是指在厭氧條件下有機物質被厭氧菌分解產生甲烷和CO2等氣體的過程,是一種國內外常用的廢水、污泥以及工業廢物的穩定處理方法[2]。由于厭氧消化技術可以在處理廢物的同時回收資源作為能源,因此被大眾認可并廣泛應用。
厭氧消化過程分為水解酸化、產氫產乙酸以及產甲烷3個階段。傳統的厭氧消化技術追逐高效能的甲烷回收,而與甲烷相比,氫氣作為未來能源,以無污染、清潔無碳、能量密度高等特點被視為極具潛力的可再生替代能源[3],是實現國家“雙碳”目標的重要支撐[4]。氫氣是厭氧消化產氫產乙酸階段的主要產物之一,而該階段另一產物——乙酸也是重要的化工原料,可用作酸度調節劑、酸化劑、腌漬劑、增味劑、香料、抗微生物劑等[5],具有較大的回收利用價值。較之于以甲烷為終端產物的傳統厭氧消化技術,氫氣和乙酸的形式更加綠色清潔。因此,若采用可行的方法將厭氧消化控制在產氫產乙酸階段并回收此階段產生的乙酸和氫氣,避免廢棄物中的碳源向甲烷或CO2等溫室氣體的轉化,可有效緩解清潔能源緊缺和廢棄物污染治理壓力,滿足國家“雙碳”政策要求,為傳統厭氧消化技術的改進提供新思路。
鑒于此,本研究從厭氧消化的原理概述出發,聚焦厭氧消化的產氫產乙酸階段,探討產氫產乙酸階段的強化措施,闡述目前可行的乙酸、氫氣回收方法,以期以乙酸和氫氣的形式回收有機固體廢棄物中的資源,為廢棄物“減排”處理和資源化利用提供理論依據。
1 厭氧消化原理概述
1.1 厭氧消化過程
厭氧消化過程主要由厭氧微生物完成,因此也稱為厭氧生物處理技術。20世紀70年代以來,大量學者對厭氧消化過程和相關微生物進行了深入探究,在厭氧消化二段理論的基礎上提出了三段理論,即將厭氧消化的過程分為水解酸化、產氫產乙酸和產甲烷3個階段[6]。其中,水解酸化階段由細菌、真菌、原生動物等分泌蛋白酶,將大分子有機物分解為小分子有機物;產氫產乙酸階段由產氫產乙酸菌(HPA)將水解酸化后期產生的不能被甲烷菌直接利用的有機物轉化為乙酸等揮發性脂肪酸(VFA)、氫氣和CO2,同時同型產乙酸菌利用氫氣和CO2產生乙酸;最終,在產甲烷階段,由產甲烷菌利用乙酸、氫氣和CO2等生成甲烷。參與產氫產乙酸階段的細菌主要包括HPA和同型產乙酸菌。
1.2 產氫產乙酸階段優勢菌
HPA是一種嚴格厭氧的細菌,與產甲烷菌相比,HPA世代周期短,十幾分鐘到幾個小時內即可繁殖一代,其參與的反應難以自發進行,需要和其他菌群(產甲烷菌、同型產乙酸菌、反硝化菌、硫酸鹽還原菌等)協同作用來完成厭氧消化,也稱為共生聯合或互營聯合作用。此外,HPA菌群在整個厭氧消化的過程中處于中間位,起到了承上啟下的重要作用[7]30。
同型產乙酸菌是一類既能利用多種有機底物異養生長,又能利用氫氣和CO2或CO自養生長的厭氧微生物類群。同型產乙酸菌可消耗氫氣,進而減輕由于氫氣積累造成的產氫產乙酸階段抑制,同時還可互營協助HPA產生酸類物質[8-9]。當產甲烷菌被抑制后,同型產乙酸菌便會代替產甲烷菌進行耗氫作用。研究報道,添加氯仿可抑制同型產乙酸菌,進而得到較高的產氫量;但若抑制了同型產乙酸菌,則不能利用其對產酸的促進作用來得到較高的產酸量,因此對同型產乙酸菌的調控至關重要。
1.3 產氫產乙酸階段的影響因素
1.3.1 pH
pH是生物處理以及厭氧代謝過程中的重要參數,參與厭氧消化的微生物對生長環境條件十分敏感,pH的變化不僅會影響HPA,也會對產甲烷菌的增殖速度和代謝途徑造成影響。HPA的最適pH為5.5~7.2。有研究表明,在不同初始pH下,厭氧污泥對丁酸、乙酸、氫氣等基質的轉化速率均有所差異[7]29。
1.3.2 無機鹽類
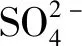
1.3.3 氨基酸
厭氧發酵離不開有機物的作用,而氨基酸是構成生命體中蛋白質的小分子單位,也是厭氧消化的中間代謝產物。國內外許多學者探究了氨基酸對厭氧發酵微生物的影響以及對厭氧消化第2、3階段產物的影響。SHARMA等[11]的研究結果表明,投加丙氨酸、半胱氨酸等不僅促進了厭氧消化產酸,也使產氫量大大提升。
根據三段4菌群理論,厭氧消化過程涉及的微生物包括水解產酸菌、HPA、同型產乙酸菌和產甲烷菌。大多數水解產酸菌和HPA對溫度、pH、有機酸濃度、氧氣濃度等環境條件適應性強,而產甲烷菌對上述環境要求較為苛刻,所以一般情況下只要滿足了產甲烷菌的生長要求,水解產酸菌和HPA便能正常生長。在厭氧消化體系中,水解產酸菌提供了丙酸、乙醇等產物,再經HPA作用生成乙酸、氫氣和CO2。作為厭氧消化的最后環節,產甲烷菌利用乙酸、氫氣和CO2生成甲烷。水解產酸菌和HPA產生的乙酸和氫氣積累到一定程度后會抑制產氫產乙酸過程的進行,產甲烷菌為水解產酸菌和HAP的生化反應解除了反饋抑制。因此水解產酸菌、HAP和產甲烷菌互為彼此創造合適的生存環境。此外,溫度,氨氮,VFA濃度等也會影響產氫產乙酸階段的進程[12-13]。
2 產氫產乙酸階段強化措施
2.1 構建優勢菌群
構建優勢菌群即通過向自然菌群中投加特殊的微生物,以強化某菌種對環境或污染物的反應。近年來有學者通過構建優勢菌群實現了厭氧消化某一階段的強化。不過由于HPA的互營聯合作用[14],即它總是與產甲烷菌、同型產乙酸菌、硫酸鹽還原菌等共生,只有少數HPA能被分離純化,較難通過HPA優勢菌群實現產氫產乙酸階段的強化。
張國華等[15]研究表明,pH為4.5~5.5時有利于產氫產乙酸的進行,并且氫氣濃度較高;而pH為7.0~7.5時有利于產甲烷階段的進行。根據厭氧消化3類菌群pH生態位的不同,可調控出適合HPA的pH生境,從而強化厭氧消化保持在產氫產乙酸階段。李建政等[16]研究證實,pH在6.0~6.5時,雖然厭氧消化系統的甲烷生成量明顯降低,但系統內氫氣占比仍然不高;直至pH降至4.0~5.0時,氫氣才便于收集,產氫得到實質性的強化。王碩[17]通過調節碳源、溫度、pH、鹽類等多種因子實現了產氫產乙酸階段優勢菌群的構建,提高了厭氧消化系統氫氣產率。王祥錕等[18]研究顯示,HPA的pH生態幅較窄,且會影響乙酸的轉化速率。唐澤雨[19]發現不斷提高的總酸對產甲烷菌有一定影響,從而會影響乙酸和氫氣的濃度和產量。
綜上可見,通過調節厭氧消化系統pH等方式創造適宜菌群生活的環境來強化產氫產乙酸過程,從而提高產氫產乙酸量是許多學者研究的重點。雖然僅調節pH即可實現乙酸和氫氣的累積,但累積量的增效及回收可行性還有待進一步研究[20-22]。此外,在對pH進行調節時,需要注意整個厭氧消化過程中的限速步驟和各菌群之間的相互關系。
2.2 抑制產甲烷過程
近年來,多數學者將目光聚焦于抑制產甲烷過程來使厭氧消化控制在產氫產乙酸階段,從而達到強化產氫產乙酸的目的。而相較于構建優勢菌群,產甲烷抑制技術則較為成熟。目前的產甲烷抑制技術可粗略分為內源抑制、外源抑制和生物抑制[23]。
內源抑制是針對剩余污泥本身包含的產甲烷抑制物抑制產甲烷菌,剩余污泥中的腐殖質、氨氮均可抑制產甲烷菌活性。
外源抑制即通過外部投加產甲烷抑制劑達到抑制產甲烷的目的,在實際應用中效果較好。典型的產甲烷抑制劑有二溴乙烷基磺酸鈉(BES)、氯仿、抗生素等。研究表明,BES對污泥厭氧消化過程中產甲烷菌的抑制作用十分明顯,投加0.06 mol/L的BES可使甲烷產量降低近98%[24]。然而BES對水解產酸過程也有明顯的抑制作用,在污泥中投加BES后,污泥的酸化程度有所降低。此外,氯仿也可作為產甲烷抑制劑,但以氯仿作為抑制劑時所得的乙酸濃度比以BES作為抑制劑時所得的乙酸濃度低。鑒于BES對水解產酸過程也存在抑制作用,采用投加BES抑制產甲烷過程從而實現乙酸積累的方法還需進一步斟酌。此外,抑制革蘭氏陰性菌的活性也能達到抑制產甲烷的目的,LI等[25]在研究畜禽養殖廢水厭氧消化過程中發現抗生素通過抑制革蘭氏陰性菌抑制了產甲烷過程。WANG等[26]還發現了羅紅霉素對產甲烷的抑制作用。
生物抑制是指通過改變電子受體,使電子流向除產甲烷菌以外的其他微生物來實現產甲烷階段的抑制。HPA和產甲烷菌的本質區別在于HPA是細菌,而產甲烷菌是古菌,具有古菌的特殊組成成分和產甲烷代謝功能。產甲烷菌的細胞壁由雜多糖和蛋白質亞基等組成,而HPA的細胞壁主要成分則是胞壁酸;產甲烷菌細胞膜上的磷脂雙分子層主要由醚鍵連接,而HPA的磷脂雙分子層主要由酯鍵連接;產甲烷菌細胞內有獨特的輔助因子——各類輔酶參與甲烷的合成。針對上述特征,若能從細胞組成的角度考慮抑制產甲烷或是阻礙輔助因子發揮作用,將會是一個值得深入的研究方向和有意義的途徑。
2.3 聯合作用強化體系
由于HPA的生態位處于水解產酸菌群和產甲烷菌群之間,起到承上啟下的作用,并且在生理上更接近產甲烷菌,想要抑制厭氧消化反應的第3階段,淘汰產甲烷菌,可以使反應體系中的優勢菌群傾向于水解產酸菌和HPA。如上所述,僅靠調節pH等方式較難實現產甲烷階段的抑制,因此通過多種方式聯合作用強化厭氧消化產氫產乙酸階段更加現實。
蘇曉煜[27]15-17通過不斷下降厭氧連續攪拌反應器(CSTR)系統中的pH使堿度最終維持在360 mg/L左右,發現當pH降至4.2~4.5時未能實現HPA的積累;當調節pH至中性,同時連續投加BES和氯仿,系統的產氫能力提高了9倍。調節pH和投加產甲烷抑制劑的聯合作用可初步實現乙酸和氫氣的積累。
許科偉[28]研究發現,氯仿對厭氧消化系統中的所有微生物存在抑制作用,包括HPA和同型產乙酸菌。因此在使用氯仿作為產甲烷抑制劑時,還需結合其他手段最終實現氫氣和乙酸的積累。而考慮到同型產乙酸菌的作用,可采用產氫產乙酸和同型產乙酸耦合體系,在產氫產乙酸階段生成氫氣、乙酸和CO2等的同時,利用同型產乙酸菌將生成的CO2固定化為乙酸。
2.4 其 他
除上述產氫產乙酸階段強化措施之外,在暗發酵生物制氫技術中,常采用物理、化學等各種預處理方法強化生物產氫或產乙酸,如熱處理、酸堿處理、微波超聲、紫外處理等方式。在厭氧消化產氫產乙酸小試研究過程中也可采用這些預處理方式,以此強化產氫產乙酸量,但鑒于工程運營成本的考慮,在此不做贅述。
3 乙酸和氫氣的回收
3.1 乙酸回收
厭氧消化過程中的乙酸作為中間產物很快被產甲烷菌消耗,想要收集乙酸必須強化產氫產乙酸過程,抑制產甲烷過程;但是當乙酸濃度較高時,又抑制HPA和同型產乙酸菌的進一步反應,乙酸難以大量積累,因此乙酸的回收必須及時且連續。
縱觀國內外,從厭氧消化產氫產酸階段回收乙酸的研究甚少,大多數研究聚焦在從工業廢水中回收乙酸。張歡歡等[29]對工業廢水中乙酸的回收方法做了整理,當廢水中乙酸質量分數低于1%時可以用電滲析和滲透法收集乙酸;乙酸質量分數在1%~10%時可以采用吸附法和反應萃取法收集,質量分數在10%~30%時則多借助精餾分離回收。王元元等[30]在對厭氧消化系統中有機酸進行研究時,采用吸附法成功從污泥中回收了有機酸,其中乙酸占比在60%左右。若控制厭氧消化過程在產氫產乙酸階段,且使得體系中乙酸積累到一定量時,可以嘗試采用吸附、萃取等方法對乙酸進行回收。
3.2 氫氣回收
氫氣是厭氧消化的一種重要中間代謝產物[31],由于厭氧消化小試系統產生的氫氣量較少,回收往往采用排水或排氣法。工業生產中產生的氫氣總是與甲烷等混合在一起,因此回收方法以膜分離法為主,也有采用吸附、吸收、冷凝等方法回收。郝小明[32]對工業氣體混合物中的氫氣進行回收,在原有基礎上增設了一臺膜分離裝置,使氫氣回收率大大提升;穆正德等[33]利用帶有滲透膜的裝置使氫氣與其他氣體組分分離,裝置的氫氣回收率≥92%,再通過氣液分離器實現氣液的分離。產氫產乙酸階段中的氣態產物主要為氫氣和CO2,采用膜分離法時要注意兩者的特性和其他分離影響因素。
目前,國內外對于抑制產甲烷階段來回收氫氣的做法還缺乏大量可靠的實驗支撐,但已有學者通過兩相厭氧以及優化厭氧反應器的方法探究產氫階段氫氣的產量[34]。KHONGKLIANG等[35]研究了不同濃度的木薯淀粉和玉米淀粉廢水的中溫兩相厭氧產氫產甲烷,其最大氫氣產量可達到81.5 L/kg;因此,未來通過產氫產乙酸階段的強化提高氫產氣量,再通過膜處理、吸附或吸收的方式進行回收是有望實現的,但采用膜分離法回收氫氣可能會面臨價格昂貴和收集、運輸不安全等問題。
4 技術可行性
基于國家“雙碳”戰略,提出了厭氧消化過程控制技術減少固體廢棄物處理過程的碳排放,期望以乙酸和氫氣的形式實現固體廢棄物的“減排”和“資源回收”。傳統污水處理的模式和觀念也逐漸由“污染去除”向“資源回收”轉變[36-37]。許多學者針對厭氧消化產物氫氣及乙酸等的回收進行了實踐,潘欣榮[38]通過調控pH實現了厭氧消化過程的連續、高選擇性產乙酸;姜錦玉[39]利用酵母廢水厭氧消化制氫得到了較好的效果;蘇曉煜[27]31-32則通過調節pH和投加產甲烷抑制劑的聯合作用初步實現了乙酸和氫氣的積累;劉楊等[40]從厭氧廢水中收集了足量氫能;陳瑞紅[41]在生物油加氫水相中成功回收了乙酸。可見,以乙酸和氫氣的形式回收厭氧消化中的有機物是技術允許且經濟可行的。
以氫氣和乙酸的形式回收固體廢棄物中的資源,同時實現厭氧消化過程的“減排”,是“雙碳”政策下厭氧消化的最佳出路[42-44],雖然目前該技術的應用化、規模化仍有挑戰和難度,但仍具可行性。
5 結 語
采用物理、化學、生物手段把厭氧消化系統控制在產氫產乙酸階段,以乙酸和氫氣的形式回收固體廢棄物中的資源,是一個值得深入探究的話題和值得重點關注的方向。從經濟效益來看,產氫產乙酸階段所產生的氫氣較甲烷具有熱值高、價值高、無污染等優點,另一產物乙酸可用于工業生產,尤其是隨著“雙碳”政策的施行,零碳排放將為企業帶來巨大收益;從社會效益來看,阻斷厭氧消化進程,避免CO2、甲烷等溫室氣體的排放,有利于全球生態環境保護。
目前國內外對于強化產氫產乙酸階段的措施主要包括培養優勢菌群、采用產甲烷抑制劑、多種方式聯合作用等。各項研究均處于初步探索階段,對于具體過程的優化、強化階段的各指標參數、HPA的性狀和產氫效率等問題仍然缺乏可靠的實驗數據支撐。如何規避或利用好菌群之間的協同作用,是否可以從生物角度抑制產甲烷過程,怎樣維持較高的、穩定的產氫產乙酸速率都是未來亟需研究和解決的難題。
