草魚β-防御素1的趨化活性及免疫佐劑效應研究
2022-12-21 07:24:38周建成劉小玲蘇建國袁改玲
水生生物學報 2022年12期
關鍵詞:檢測
況 睿 周建成 劉小玲 蘇建國 袁改玲
(1.華中農業大學水產學院, 武漢 430070; 2.武漢大北農水產科技有限公司, 武漢 430090)
草魚(Ctenopharyngodon idella)是我國淡水養殖中最主要的養殖品種之一, 2021年中國漁業年鑒統計結果顯示, 2020年我國草魚產量高達557.1083萬噸。隨著我國水產養殖業的迅速發展, 水產養殖的病害問題日漸突出。當前用于治療魚類細菌性疾病的主要是抗生素類化學藥物, 但抗生素污染問題已經成為目前影響我國水產養殖業可持續健康發展的重大科技問題。尋找能夠替代抗生素的環保型飼料添加劑, 研制出無抗生素的環境友好型飼料, 是我國水產養殖業健康發展的迫切需求[1,2]。
抗菌肽廣泛存在于各種生物體中, 是機體先天性免疫的重要組成部分[3]。抗菌肽對細菌、真菌和病毒等多種病原具有生物活性且不易產生耐藥性[3,4],同時還具有非常好的免疫調節活性, 被認為是開發抗菌藥物的新方向。Ganz等[5]將在人和兔白細胞中首次發現的一類結構相似的抗菌肽命名為“防御素”, 之后研究發現防御素幾乎廣泛分布在所有的生物體中, 根據防御素六個保守的半胱氨酸的位置及形成二硫鍵的連接方式不同可將動物防御素分為α、β和θ三種[6], β-防御素在所有脊椎動物中廣泛分布, α-防御素僅存在于哺乳動物中, θ-防御素僅在恒河猴和狒狒中被發現[7]。α-和θ-防御素被認為是從古老的β-防御素進化而來[8]。
近年來研究發現, β-防御素除了具有強大的直接抗病原微生物活性以外, 還可以在天然免疫中發揮趨化等調節作用[9], 而巨噬細胞和中性粒細胞等白細胞定向遷移到病原入侵部位對宿主防御和誘發免疫反應至關重要[10]。了解防御素對魚類白細胞的趨化作用及其免疫佐劑效應可以為魚類防治微生物感染及疫苗提供新的策略, 為魚類β-防御素在生產實踐中的應用提供理論依據。
1 材料與方法
1.1 實驗草魚和細菌
實驗草魚[(1000±100)g 和(30±5)g]購自湖北省仙桃市排湖風景區漁場, 在華中農業大學水產學院循環水養殖系統中暫養2周。水溫控制在(26±2)℃, 每日投喂魚體重量2%的商品化飼料(武漢大北農集團)。實驗魚通過華中農業大學實驗動物中心動物倫理審核(倫理號: HZAUFI-2021-0018), 一切操作均嚴格遵守實驗動物福利倫理原則。金黃色葡萄球菌(S.aureus, ATCC 25923)、無乳鏈球菌(S.agalactiae, ATCC 13813)、大腸桿菌(E.coli,ATCC 25922)和嗜水氣單胞菌(A.hydrophila, ATCC 7966)分別從美國標準菌庫(American Type Culture Collection, ATCC)中獲得, 由本實驗室擴大后自主保管。
1.2 生物信息學分析
從NCBI(https://www.ncbi.nlm.nih.gov/)中下載得到人類(Homo sapiens)、小鼠(Mus musculus)、原雞(Gallus gallus)和包含草魚在內的多種魚類β-防御素的氨基酸序列。將包含所有序列信息的文檔上傳到Clustal Οmega在線程序(https://www.genome.jp/tools-bin/clustalw)完成各物種β-防御素的多序列比對, 并通過MEGA7軟件構建NJ系統發育樹。
利用在線網站https://services.healthtech.dtu.dk/service.php?SignalP-5.0預測CiBD1成熟肽, 將成熟肽序列通過在線同源建模系統SWISS-MODEL(http://swissmodel.expasy.org/)預測CiBD1三級結構。之后將其三級結構導入PyMOL軟件(PyMOL Molecular Graphics System, Version 1.3, 2011)中進行蛋白質三級結構成像, 進行可視化分析。最后, 利用軟件直觀展現蛋白質三級結構, 分析CiBD1的α螺旋與β折疊結構和CiBD1的表面凈電荷分布。
1.3 引物合成
根據CiBD1(GenBank登錄號: KX906958.1)基因核苷酸序列, 利用Primer 5.0軟件設計特異性引物, 在引物上游引入KpnⅠ限制性內切酶和腸激酶(Enterokinase, EK)酶切位點, 在引物下游引入XhoⅠ限制性酶切位點。實驗中用于擴增CiBD1基因及實時熒光定量PCR(Quantitative Real-time PCR,qRT-PCR)使用的引物序列見表1, 由武漢擎科生物工程有限公司合成。
1.4 A.hydrophila感染前后草魚各組織中CiBD1的轉錄分析
隨機取5尾健康草魚[(30±5)g]用MS222麻醉,先進行尾靜脈放血然后解剖取各組織(脾臟、皮膚、鰾、肝胰腺、鰓、后腸、腦、眼、頭腎和體腎)混樣后用TRIzol試劑盒(中國北京, Aidlab)提取總RNA。使用NanoDrop 2000分光光度計測定濃度并根據260和280 nm下的吸光度比值判斷RNA質量,A260/A280值在1.8—2.0為佳。通過2%瓊脂糖凝膠電泳檢測RNA完整性。使用HiScript?Ⅱ Q RT SuperMix for qPCR with gDNA wiper(中國南京,Vazyme)試劑盒按說明書操作進行反轉錄得到cDNA。qRT-PCR反應體系(20 μL)為: AceQ?qPCR SYBR Green Master Mix (中國南京, Vazyme)10 μL, 上下游引物(表1)各0.4 μL, cDNA 1.0 μL, ddH2O 8.2 μL。反應程序為: 94℃預變性3min; 95℃變性10s; 60℃退火30s; 72℃延伸20s; 重復40個循環。溶解曲線:95℃ 15s, 60℃ 1min , 95℃ 15s。以草魚β-actin作為內參基因, 每個樣品設置3個重復。使用2–??Ct方法計算各組織CiBD1相對轉錄水平, 將感染前脾臟轉錄水平設為1, 其余組織是此基準的相對值。

表1 本研究所用到的引物Tab.1 Primers used in this study
取5×107CFU/mL嗜水氣單胞菌懸液以200 μL/尾注射20尾草魚, 72h后進行取樣。之后操作步驟同感染前草魚樣品的處理一致。
1.5 原核表達載體構建與CiBD1重組蛋白的獲得
根據預測得到的成熟肽對應的核苷酸序列設計特異性引物, 原核表達載體選擇pET-32a。以草魚后腸cDNA為模板, 采用PCR擴增目的片段, 用1.5%瓊脂糖凝膠進行電泳檢測。使用Gel Extraction kit 試劑盒(中國杭州, Axygen), 參照說明書純化回收CiBD1的PCR擴增產物。用限制性內切酶KpnⅠ(中國北京, TaKaRa)和XhoⅠ(中國北京, TaKaRa)對CiBD1的PCR擴增片段和pET-32a載體進行雙酶切, 37℃水浴酶切30min, 將酶切產物回收后進行連接。采用大腸桿菌DH5α感受態細胞進行重組載體的轉化和陽性克隆篩選, 以單個克隆的菌液為模版,用CiBD1擴增引物進行 PCR擴增, 選擇 PCR擴增檢測陽性的菌落送武漢擎科生物科技有限公司進行測序確認。確認無誤后擴大培養菌液并提取質粒,轉化到大腸桿菌表達菌株 BL21(DE3)pLysS中進行重組表達, 轉化和鑒定方法同上。經確認的重組載體命名為pET-32a-CiBD1。
表達菌株的最適誘導條件: 1.0 mmol/L IPTG,37℃, 5h。誘導結束后5000 r/min離心10min收集菌體, 使用平衡緩沖液(10 mmol/L Tris, 50 mmol/L NaCl, 20 mmol/L 咪唑, pH=7.2)重懸菌體, 使用高壓破碎儀(中國上海, 勵途, FB-110X)在850 Pa下破碎5min, 然后12000 r/min離心60min, 分別收集上清和沉淀。添加 5×SDS-PAGE上樣緩沖液后, 在沸水中變性10min后于–20℃保存樣品。用SDS-PAGE凝膠試劑盒(中國北京, Solarbio)配制12% SDSPAGE凝膠。取上清和包涵體樣品進行點樣檢測,凝膠電泳條件為 80 V 2h。在電泳完成后, 分離SDS-PAGE凝膠進行考馬斯亮藍染色1h, 用水沖洗凝膠后, 于沸水中脫色20min后拍照檢測。確定CiBD1表達于上清后, 采用Ni-NTA柱進行蛋白純化, 使用腸激酶去除標簽蛋白, 收集酶切產物。用Tris-Tricine-SDS-PAGE 凝膠試劑盒(中國北京, Solarbio)配制15%凝膠進行酶切產物檢測。使用His標簽抗體對重組蛋白進行免疫印跡(Western Blot)檢測。
1.6 CiBD1重組蛋白的體外抑菌活性檢測
本研究中檢測CiBD1重組蛋白抗菌活性的方法為 CFU平板法。將凍存的金黃色葡萄球菌(ATCC 25923)、無乳鏈球菌(ATCC 13813)、大腸桿菌(ATCC 25922)和嗜水氣單胞菌(ATCC 7966)接種到LB培養基中復蘇。將復蘇的菌液以1﹕100比例重新接種到新LB培養基中培養至對數期。使用平衡緩沖液稀釋細菌到1×106CFU/mL, 吸取50 μL菌液與含CiBD1蛋白100 μg/200 μg的蛋白溶液150 μL混合后在細菌最適生長溫度(金黃色葡萄球菌、無乳鏈球菌和大腸桿菌為37℃, 嗜水氣單胞菌為28℃)水浴1h, 吸取50 μL 涂布在無抗LB固體平板上, 培養16h后拍照計數, 以Tris-HCl緩沖液為陰性對照。
1.7 CiBD1重組蛋白的趨化活性分析
用Percoll梯度離心法分離草魚[(1000±100)g]頭腎白細胞[11]。清洗兩遍后調整細胞濃度為1×106CFU/mL。在遷移小室(Corning, 規格3 μm)中進行趨化實驗, 將500 μL含有5% FBS(Fetal Calf Serum, 胎牛血清)和不同濃度CiBD1蛋白(0、12.5、25、50、100和200 ng/mL)的DMEM培養液(美國, Life Technologies)引入外室, 在內室中引入300 μL白細胞懸液。28℃趨化2h后收集外室的細胞懸液, 采用流式細胞儀(美國, BD Biosciences)進行計數, 并基于SSC(前向角散射)和FSC(側向角散射)參數分析遷移細胞的種類。
1.8 滅活疫苗制備與個體感染實驗
采用福爾馬林滅活法制備嗜水氣單胞菌滅活疫苗, 首先將嗜水氣單胞菌復壯培養24h后加入福爾馬林溶液(分析純, 國藥集團化學試劑有限公司),使終濃度為0.2%, 37℃滅活24h。將滅活后的嗜水氣單胞菌于5000 r/min 離心10min 沉淀菌體, 棄置上清后用PBS 懸浮再離心以去除殘余的甲醛, 重復3次。用PBS稀釋調整菌的濃度至1.0×108CFU/mL, 即為嗜水氣單胞菌滅活疫苗。將滅活疫苗與Montanide ISA206(法國, Seppic)佐劑按1﹕1(質量比)準備, 然后分別加熱至(30±2)℃, 將Montanide ISA206佐劑倒入組織勻漿機中, 以200 r/min攪拌,并將滅活疫苗緩慢加入, 待滅活疫苗加完后, 提高攪拌速度至2000 r/min, 保持在30℃攪拌10min后逐漸降低攪拌速將苗液冷卻至15℃停機, 在15℃以下靜置24h后完成制備, 于4℃保存備用。CiBD1的佐劑組是先將CiBD1重組蛋白與嗜水氣單胞菌滅活疫苗混合后再按上述方法與Montanide ISA206佐劑混合完成制備。吸取100 μL 疫苗涂布在無抗LB固體平板上, 37℃培養72h, 期間沒有細菌或真菌生長,完成疫苗的無菌性檢測。以100 μL/尾的劑量, 將制備好的各疫苗組注射到20尾健康草魚體內, 以PBS作為陰性對照。觀察7d后, 草魚未出現死亡即完成疫苗的安全性檢測。
將暫養兩周的健康草魚[(30±5)g]隨機分為對照組, 疫苗組和疫苗+CiBD1組置于3個單獨的400 L養殖水缸中。每組3個平行, 每個平行30尾魚, 每組的3個平行在同一個大缸中并用特制網箱分隔開。每尾魚腹腔注射100 μL PBS/疫苗/疫苗+CiBD1進行免疫, 控制水溫為(28±2)℃, 免疫14d后用半致死濃度的嗜水氣單胞菌(5×107CFU/mL)進行腹腔注射感染, 每尾注射量為200 μL。在感染前及感染后14d分別進行1次取樣, 包括血清、頭腎和后腸。注射感染后每8h監測1次, 統計14d的死亡率。
1.9 血清酶活測定和免疫相關基因qRT-PCR檢測
采用尾椎靜脈抽取草魚[(30±5)g]血液后在4℃靜置12h, 在4℃, 3000×g下離心15min, 收集血清。血清總超氧化物歧化酶(Total superoxide dismutase,T-SOD)、溶菌酶活性(Lysozyme Activity, LA)和補體C3酶活性的測定采用南京建成生物工程研究所生產的試劑盒。免疫相關基因采用qRT-PCR檢測方法同1.4。
1.10 數據處理與統計學分析
采用Microsoft Office Excel 2010和GraphPad Prism 7.0軟件進行數據分析、統計分析和圖形展示。結果以平均數±標準差(SD)的方式表示。用單因素變量方差分析法(ANOVA)對抑菌、趨化等只有單一變量的實驗進行統計學分析, 兩組間采用非配對雙尾Student’st檢驗判斷是否有統計學差異,用Mantel-Cox檢驗對保護率數據進行分析。P<0.05被認為具有顯著差異, 并用“*”標記;P<0.01用“**”表示;P<0.001用“***”表示。
2 結果
2.1 CiBD1的生物信息學分析
參與比對的所有β-防御素均具有6個高度保守的半胱氨酸, 但是, 魚類β-防御素高度保守的第4號半胱氨酸與人、鼠和原雞的第4號半胱氨酸位置存在差異。圖1展現了人類、小鼠、原雞和多種魚類共39條β-防御素氨基酸序列的NJ系統發育樹 。使用Neighbor-Joining聚類法, 在bootstrap檢驗中, 進行 500次重復, 使用p-distance模型計算進化距離,在任何位置都允許少于50%的比對間隙、缺失數據和模糊的堿基。在進化關系上, 草魚β-防御素與鯉、團頭魴和斑馬魚更為相近。通過在線網站預測得到的CiBD1成熟肽序列為: ASFPWTCASLSGVCRQGVCLPSELYFGSLGCGKGFLCCVSH FG。使用PyMOL軟件對CiBD1成熟肽進行可視化分析, CiBD1的二級結構中C端有一個α螺旋, 另外最為明顯的特征是3個反向平行的β折疊, 這正是其高度保守的6個半胱氨酸兩兩連接組成, 是β-防御素最為典型的結構特征之一。當展現CiBD1靜電勢表面時, 其表面包含疏水基團與親水基團, 同時正電荷分布大于負電荷, 由此可見CiBD1為陽離子兩親性蛋白。
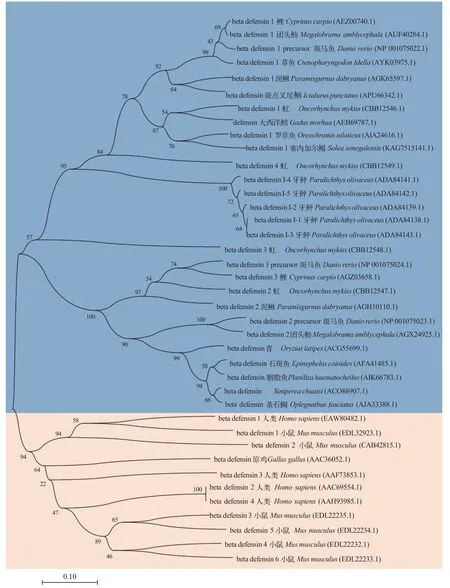
圖1 β-防御素氨基酸序列NJ系統發育樹Fig.1 NJ phylogenetic tree of β-defensin amino acid sequence
2.2 嗜水氣單胞菌感染前后CiBD1的組織轉錄水平
為了探究CiBD1的生理學作用, 我們研究了CiBD1基因在正常生理條件下草魚不同組織中的轉錄水平(圖2), 在感染前的各組織中, 皮膚和眼是CiBD1轉錄水平最高的兩個組織。嗜水氣單胞菌感染72h后的草魚各組織中CiBD1轉錄水平與感染前相比有明顯差異, 表現為與黏膜免疫相關的組織,如皮膚、鰾、后腸和眼中的CiBD1的轉錄水平較為顯著。
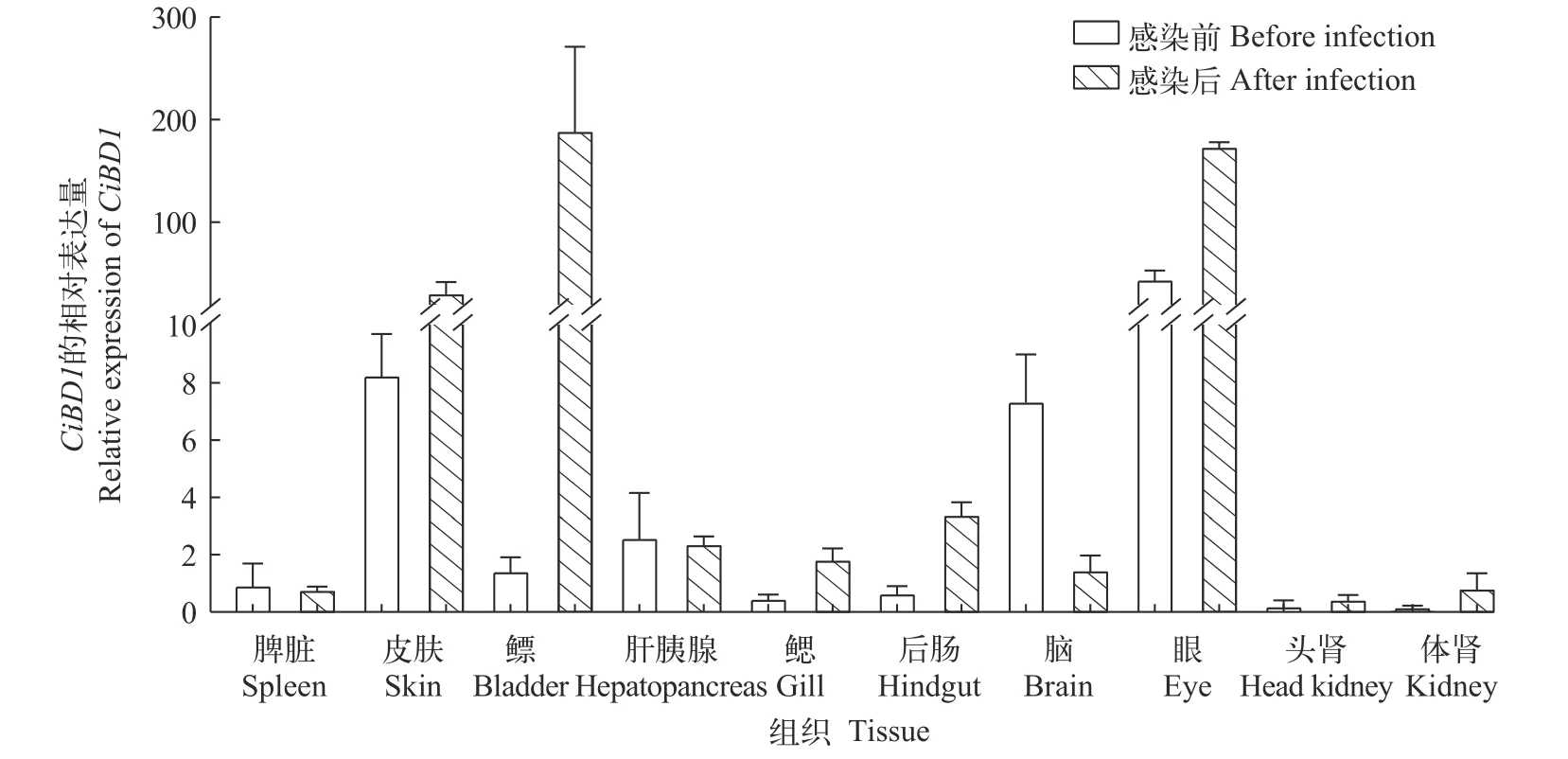
圖2 嗜水氣單胞菌感染前后CiBD1在各組織中的轉錄水平Fig.2 Constitutive levels of CiBD1 transcription in different tissues before and after A.hydrophila infection
2.3 獲得CiBD1重組蛋白
將構建的重組質粒轉化到表達菌株中進行菌液PCR后用瓊脂糖凝膠電泳檢測, 結果(圖3A)顯示在159 bp處有一條特異性條帶, 大小與預測的目的條帶一致。采用pET-32a質粒, 選擇KpnⅠ和XhoⅠ兩個酶切位點間插入目的基因片段。12%的SDSPAGE凝膠電泳結果顯示, IPTG誘導后在22.5 kD大小處出現特異性條帶(圖3B), 與重組融合蛋白預測大小一致。使用His標簽進行融合蛋白免疫印跡檢測, 結果顯示在22.5 kD大小處出現與預測大小一致的特異性條帶(圖3C)。腸激酶切后用15%的Tris-Tricine-SDS-PAGE凝膠檢測結果顯示在4.7 kD大小處出現特異性條帶(圖3D), 與目的蛋白大小一致。
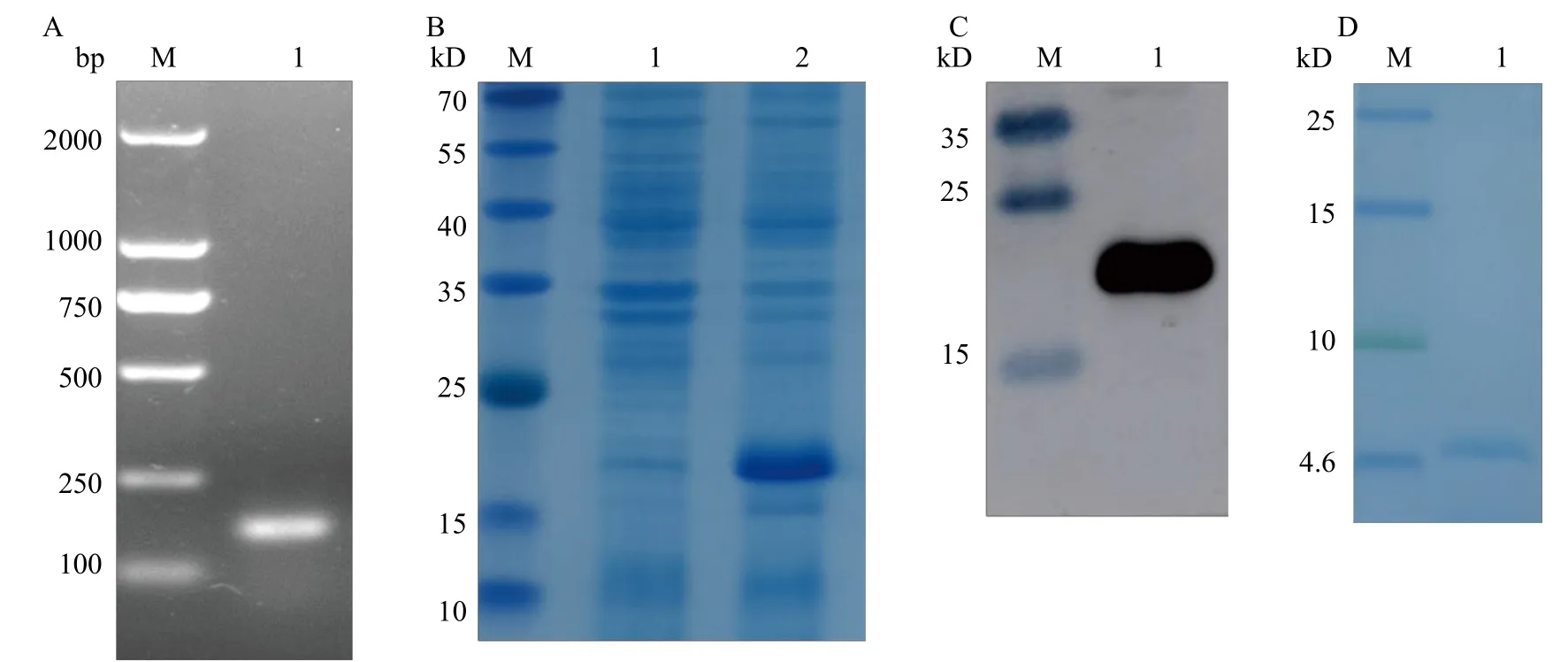
圖3 重組質粒構建與蛋白表達純化及檢測Fig.3 Construction of recombinant plasmid and expression, purification and detection of protein
2.4 CiBD1蛋白具有體外抑菌活性
CFU平板法檢測CiBD1蛋白的抗菌活性。選擇兩種革蘭氏陰性菌(嗜水氣單胞菌和大腸桿菌)和兩種革蘭氏陽性菌(無乳鏈球菌和金黃色葡萄球菌)進行蛋白抑菌實驗。結果顯示CiBD1蛋白對兩種G–和G+均具有抑制活性(圖4A)。對細菌平板拍照后用ImageJ軟件進行計數, 將數據轉換成柱形圖直觀展示, 并進行顯著性分析(圖4B)。

圖4 CiBD1的抗菌活性分析Fig.4 Antibacterial activity analysis of CiBD1
2.5 CiBD1蛋白對草魚原代白細胞具有趨化功能
使用Percoll梯度離心法分離草魚頭腎白細胞,吸取100 μL制備涂片后通過吉姆薩染色并進行掃描電鏡, 參照相關研究報道[12]對淋巴細胞、單核巨噬細胞和粒細胞進行了形態學觀察與鑒定(圖5A)。
采用遷移小室進行白細胞趨化實驗, 收集趨化后的外室細胞用流式細胞儀進行分群與計數(圖5B)。將各組細胞數轉換成柱形圖的方式進行展現, 淋巴系細胞與髓系細胞分開展現(圖5C和5D)。以5%FBS作為陽性對照, 結果表明在一定范圍內CiBD1蛋白對草魚白細胞的趨化遷移能力與蛋白濃度正相關。CiBD1蛋白在50 ng/mL時對草魚頭腎淋巴系細胞和髓系細胞均表現出較強的趨化活性。
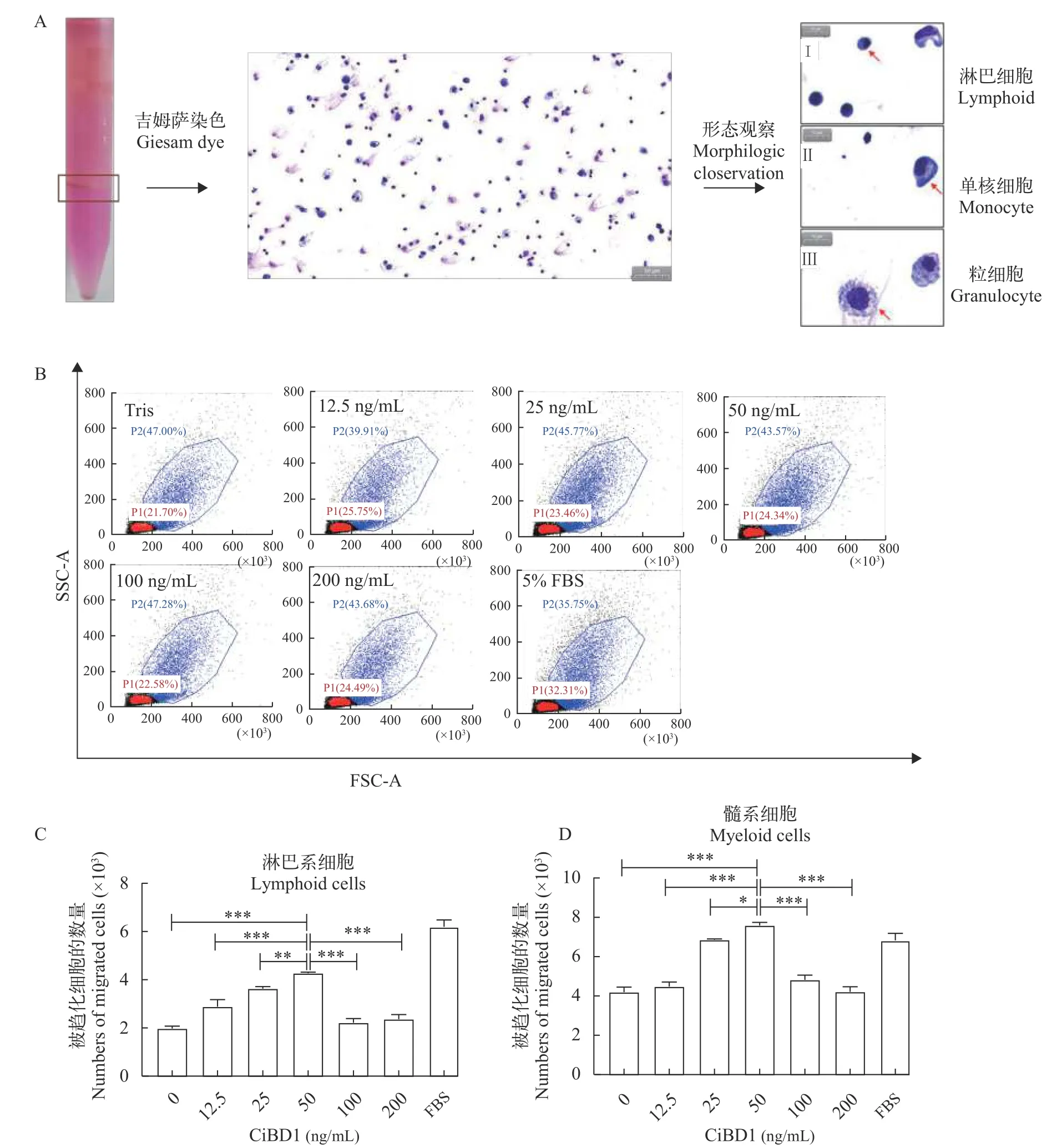
圖5 CiBD1對草魚白細胞的趨化活性Fig.5 Chemotactic activity of CiBD1 on grass carp leukocyte
2.6 CiBD1蛋白能保護草魚抵抗嗜水氣單胞菌感染
依據草魚免疫應答產生規律[13], 設置免疫和感染時間節點如圖6A所示。感染后14d的存活率曲線(圖6B)顯示, 疫苗+CiBD1組草魚存活率最高, 且與疫苗組和對照組存在顯著性差異。
用分光光度法對血清T-SOD、LA和C3進行測定(圖6C—E)。免疫后14d疫苗組和疫苗+CiBD1組的T-SOD、LA和C3均顯著高于對照組, 且疫苗+CiBD1組的T-SOD和LA顯著高于疫苗組。感染后14d疫苗+CiBD1組的T-SOD、LA和C3均顯著高于對照組和疫苗組。
免疫前、免疫后14d(感染前)和感染后14d分別檢測后腸和頭腎中免疫基因IL-1β、IL-6、MHCII、IgM和IgZ的表達水平(圖6F1—F4)。結果顯示,免疫后14d, 疫苗+CiBD1組頭腎中IL-1β和IgM表達水平顯著高于對照組及疫苗組,IL-6和IgZ的表達水平沒有顯著性差異(圖6F1); 疫苗+CiBD1組和疫苗組后腸中IL-1β、IgM和IgZ的表達水平較對照組顯著提高, 但疫苗+CiBD1組和疫苗組之間未表現出顯著性差異(圖6F2)。感染后14d, 疫苗+CiBD1組頭腎中IgM和MHC II表達水平顯著高于對照組和疫苗組, 疫苗+CiBD1組和疫苗組IL-6表達水平顯著低于對照組(圖6F3); 疫苗+CiBD1組和疫苗組后腸中IL-1β表達水平顯著低于對照組, 疫苗+CiBD1組MHC II表達水平顯著高于對照組和疫苗組, 疫苗組IgZ表達水平顯著高于對照組, 而疫苗+CiBD1組IgZ表達水平顯著低于對照組(圖6F4)。
3 討論
3.1 CiBD1的生物信息學與組織分布分析
防御素是一類廣泛存在于動植物界中的小型陽離子抗菌肽, 但在魚類中僅存在β-防御素[14]。對CiBD1多肽的生物信息學分析發現, 其具有β-防御素的典型結構, 即6個高度保守的半胱氨酸組成的3個β折疊結構。本研究檢測CiBD1在嗜水氣單胞菌感染前后的組織表達水平結果表明, 感染前CiBD1呈組成型分布在草魚各組織并高表達于一些黏膜組織, 而細菌感染后一些與外周環境接觸密切的組織中的CiBD1轉錄水平上升顯著。CiBD1高表達于皮膚、眼和腸道等黏膜組織的結果預示著CiBD1在黏膜免疫的防御中可能發揮著重要作用,與高等脊椎動物β-防御素主要在各種器官或組織的上皮細胞中產生一致[15]。隨著物種進化, β-防御素結構、種類和功能變得多樣, 草魚作為早期進化的脊椎動物, 目前僅發現三種結構相似的β-防御素。而在人和鼠上有多達48種β-防御素[16,17], 這些β-防御素功能多樣, 如抗細菌、真菌、病毒和寄生蟲活性, 以及趨化, 誘導細胞成熟, 補體激活, 結合LPS和損傷修復等免疫調節功能[18]。CiBD1在結構上具有高度保守性, 在功能上也與哺乳動物存在很大的相似性。
3.2 CiBD1的抗菌活性及趨化活性檢測
大腸桿菌表達系統用于外源蛋白的表達具有方便高效且低廉的優勢, 通過腸激酶去除融合標簽可以最大程度地保留重組蛋白的活性。本研究顯示純化并腸激酶切去融合標簽的CiBD1對G+和G–具有一定的抑制活性, 但是最小殺菌濃度(MIC)較高。目前研究認為, 抗菌肽主要通過膜破損的方式殺菌[19], 其殺菌能力在一定范圍與所帶正電荷成正比, 推測CiBD1較弱的電荷限制了其體外殺菌能力, 相較于PI值更高的蛋白如CXCL20a[20], CiBD1對細菌具有更高的MIC, 這與其他魚類β-防御素的報道一致[21]。
關于防御素的趨化活性, 在高等哺乳動物已有大量報道, 而且不同類型人防御素表現出對不同細胞的趨化活性[22]。比如人β-防御素2是人上第一個被證明具有趨化活性的防御素[23], 其對肥大細胞具有趨化作用[24]; 在納克濃度下, HNP-1和HNP-2可以募集單核細胞集合到炎癥部位, 并對T細胞和未成熟的樹突狀細胞也有趨化活性[25]; 魚β-防御素趨化活性也有報道[26,27]。脊椎動物免疫細胞趨化是一種重要的宿主防御機制[28,29], 尤其是抗原提呈細胞的趨化對免疫應答的啟動起著重要作用[30—32]。本研究結果中發現50 ng/mL的CiBD1重組蛋白在體外對草魚白細胞具有明顯的趨化活性, 提示CiBD1具備免疫佐劑的應用潛力。
3.3 免疫佐劑效應探究
免疫佐劑是一類能夠非特異性增強機體對抗原的免疫應答反應, 但本身不具備抗原性的物質[33]。免疫佐劑具有減少抗原使用量, 迅速激活免疫系統,增強免疫應答等作用。免疫佐劑種類很多, 包含鋁佐劑、微生物類佐劑、蜂膠佐劑、油乳佐劑、左旋咪唑佐劑、中藥佐劑及小肽類佐劑等。一些常規佐劑雖然具有一定的免疫增強效果, 但同時會存在一些副作用, 如鋁佐劑在白鼠體內可進入大腦,存在安全風險; 左旋咪唑佐劑長期使用會引起腹瀉[34]。開發生物來源的小肽佐劑是疫苗佐劑的研究新方向, 因其具有更高的生物安全性。目前研究證實β-防御素具有較好的免疫佐劑效果, 如人β-防御素3作為免疫佐劑可以增強金黃色葡萄球菌滅活疫苗對小鼠保護效果, 潛在機理可能是通過上調CD80/CD86和MHCⅡ表達水平[35]; 重組禽β-防御素rAvBD12613作為免疫佐劑在新城疫活疫苗和滅活疫苗中均具有良好的免疫佐劑活性[36]。本研究探究了CiBD1蛋白對嗜水氣單胞菌滅活疫苗的佐劑增強效應, 各組存活率如圖6B所示, 疫苗+ CiBD1組較疫苗組顯著提高了草魚存活率, 表明CiBD1可以增強疫苗免疫效果, 提高草魚抵抗嗜水氣單胞菌的感染。白細胞介素-1 (IL-1)家族成員是先天免疫系統中非常重要的炎癥因子[37],IL-1包括IL-1α和IL-1β兩種形式,IL-1β可以激活IL-6表達, 選擇性介導對血管損傷的修復[38]。本研究發現免疫后14d, 疫苗+ CiBD1組頭腎中IL-1β轉錄水平顯著高于對照組和疫苗組; 后腸中IL-6轉錄水平顯著高于對照組。感染后14d, 與對照組相比, 疫苗+ CiBD1組頭腎中IL-1β和IL-6轉錄水平均有所下降。提示CiBD1可調節炎癥因子表達水平。補體和溶菌酶是魚類先天性免疫的重要免疫分子, 本研究發現,免疫后14d和感染后14d實驗組血清中溶菌酶和C3含量高于對照組。以上結果提示CiBD1作為免疫佐劑可以增強疫苗對草魚先天性免疫系統的激活。

圖6 嗜水氣單胞菌體外感染實驗Fig.6 A.hydrophila infection in vitro
MHC II對CD4+T淋巴細胞的抗原呈遞至關重要, 本研究結果顯示, 疫苗+CiBD1組在感染14d后MHCⅡ顯著上升, 說明CiBD1可以增強疫苗的抗原提呈, 促進適應性免疫應答的啟動。IgM是硬骨魚類血清中最主要的免疫球蛋白, 在硬骨魚類抗感染免疫中發揮著重要作用[39], 此外, 硬骨魚類所特有的免疫球蛋白IgZ在功能上與哺乳動物IgA類似,主要表達于皮膚、鰓等黏膜組織, 在黏膜免疫中起著重要作用。本研究發現, 免疫后14d, 疫苗+ CiBD1組頭腎IgM轉錄水平顯著高于對照組, 而后腸IgM和IgZ轉錄水平均顯著提高。感染后14d, 疫苗+CiBD1組頭腎中IgM仍維持較高水平, 而疫苗組后腸IgZ轉錄水平顯著高于疫苗+ CiBD1組。這提示在免疫階段IgM和IgZ在頭腎和后腸中會大量表達,參與機體的抗感染免疫反應; 感染后IgM和IgZ會發揮免疫防御作用, 參與病原體的清除。以上結果提示CiBD1可以提高疫苗對適應性免疫的應答能力,具有潛在的免疫佐劑效應。
4 總結
綜上所述, CiBD1 是一類高度保守的小分子陽離子抗菌肽,CiBD1呈組成型分布在草魚各組織并高表達于一些黏膜組織, 在機體受到病原或環境刺激時, 一些與外周環境接觸密切組織中的CiBD1轉錄水平表達更顯著。CiBD1重組蛋白不僅在體外具有一定的抑菌活性, 而且對草魚頭腎淋巴系細胞和髓系細胞均表現出較強的趨化活性。CiBD1可以提高嗜水氣單胞菌滅活疫苗的保護率和抗病力,具有良好的免疫佐劑效應。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
