環(huán)狀RNA在肝纖維化中的調控機制及研究進展
2022-12-21 02:49:52周家名李彬彬余宏宇
臨床與實驗病理學雜志 2022年11期
周家名,李彬彬,余宏宇
肝纖維化是一種復雜的損傷-應答修復反應,由各種慢性肝損傷觸發(fā)。竇周隙內肝星狀細胞(hepatic stellate cell, HSC)的異常活化和肝臟細胞外基質(extracellular matrix, ECM)的過度沉積,是肝纖維化發(fā)生的關鍵過程。進行性肝纖維化最終將導致肝硬化,預后差,并伴較高的病死率。鑒于目前對肝纖維化尚無有效的早期診斷和治療手段,故亟需進一步深入探討其發(fā)生、發(fā)展的分子機制,尋找具有臨床轉化價值的分子診療靶點。近年,環(huán)狀RNA(circular RNA,circRNA)在肝纖維化中的調控作用逐漸被重視。越來越多的研究發(fā)現,肝纖維化組織中circRNA表達譜有顯著差異性改變,circRNA可能作為肝纖維化的新分子生物學標志物和治療靶點。本文對近年circRNA在肝纖維化中的調控機制及研究進展進行綜述,并展望該領域的發(fā)展趨勢。
1 circRNA的生物學功能
2013年,Nature雜志上刊登了Memczak和Hansen等對circRNA的研究成果,從此揭開了circRNA的研究序曲[1-2]。Memczak等[1]在人、鼠和線蟲中分析得到數千種穩(wěn)定表達的circRNA,并發(fā)現circRNA似乎在細胞類型或發(fā)育階段特異性表達。circRNA一般來源于pre-mRNA的直接反向剪接或線性RNA拼接過程中形成的套索RNA中間體,以共價鍵形成閉合環(huán)狀結構。根據其來源和構成,circRNA主要可分為外顯子環(huán)化的circRNA(exonic circRNA, ecircRNA)、內含子環(huán)化的circRNA(intronic circRNA, ciRNA)和外顯子-內含子共同環(huán)化的circRNA(exon-intron circRNA, EIciRNA)三種類型[3-4]。此外,近年一些報道發(fā)現新的circRNA形式,如融合circRNA、轉錄讀取circRNA等[5-6]。circRNA的主要功能包括充當miRNA海綿、蛋白支架、調節(jié)轉錄和調控翻譯等[7-10]。其中,EIciRNA/ciRNA通常定位于胞核,并主要參與基因轉錄[11-12]。ecircRNA通常定位于胞質,此類circRNA多具有顯著的“超級海綿”能力,可通過與miRNA的競爭性結合導致miRNA下游靶基因釋放,提高靶基因水平[13-14]。
2 肝纖維化的發(fā)生機制
肝纖維化實質是肝內慢性損傷后膠原纖維持續(xù)異常增多的病理過程[15],存在于竇周隙內的HSC是肝纖維化發(fā)生的主要效應細胞。HSC激活是肝纖維化發(fā)生的關鍵事件,其活化過程可分為啟動階段和持續(xù)階段。啟動階段中受損實質細胞(如肝細胞)旁分泌或細胞損傷碎片刺激引發(fā)炎癥反應,促進HSC發(fā)生早期活化。啟動階段刺激的維持會導致HSC進入由自分泌和旁分泌,共同刺激調節(jié)的持續(xù)活化階段,出現增殖、趨化、纖維生成、收縮性、基質降解和維生素A喪失等細胞的行為改變[16-17]。活化后的HSC(即肌成纖維細胞)產生α-SMA,并大量分泌ECM,使竇周隙內出現以膠原纖維為主的ECM異常沉積,引發(fā)肝纖維化。此外,肝纖維化也受到肝內其他細胞和多種細胞因子、信號通路的共同調節(jié)[18-19]。
3 肝纖維化領域circRNA的研究現狀及主要調控機制
近年,隨著高通量技術的發(fā)展和研究手段的進步,circRNA在HSC活化/肝纖維化過程中的調控機制研究不斷深入。文獻報道多數circRNA通過海綿吸附miRNA,調節(jié)肝纖維化相關基因的表達水平,直接參與肝纖維化調控(圖1)。這些circRNA通過控制早期炎癥反應,調節(jié)效應細胞活化及調控ECM生成等方式,調控肝纖維化過程,有望成為肝纖維化的新生物學標志物和潛在治療靶點(表1)。
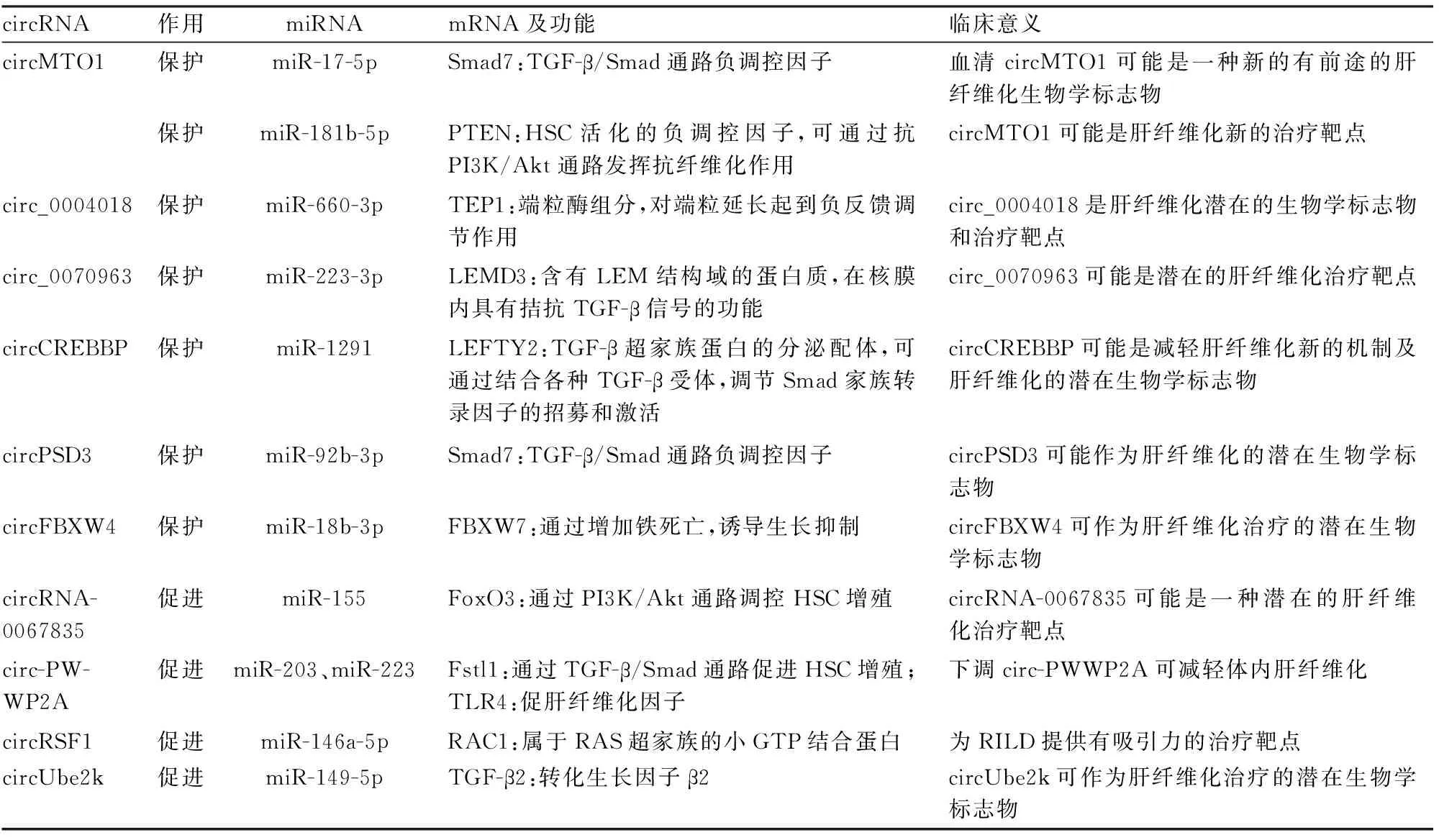
表1 肝纖維化中circRNA-miRNA-mRNA的調控軸

圖1 circRNA調控肝纖維化的作用機制
3.1 對肝纖維化發(fā)揮保護性作用circRNA
3.1.1circMTO1 Wang等[20]發(fā)現血清circMTO1在慢性乙型肝炎患者中顯著下調,ROC曲線分析顯示,血清circMTO1可作為慢性乙型肝炎患者肝纖維化的診斷生物學標志物,并與纖維化分期和HAI評分呈負相關。circMTO1可通過海綿吸附miR-17-5p調控Smad7的表達。Smad7是TGF-β/Smad通路中的負調控因子[21],circMTO1可通過吸附miR-17-5p促進Smad7表達,抑制肝纖維化。
Jin等[22]報道circMTO1在肝硬化患者的肝臟中表達減少,在肝纖維化小鼠肝臟和激活的HSC中表達下調。circMTO1和miR-181b-5p共定位于細胞質,miR-181b-5p可抑制PTEN的表達。PTEN是HSC活化的負調控因子[23],該研究證實circMTO1至少部分通過吸附miR-181b-5p調控PTEN表達而抑制HSC活化。
3.1.2circ_0004018 Li等[24]發(fā)現circ_0004018在CCl4誘導的肝纖維化小鼠模型中表達水平降低,并且circ_0004018可作為miR-660-3p的海綿,調節(jié)miR-660-3p靶基因TEP1的表達。TEP1是端粒酶的組成部分,可參與端粒的維持。circ_0004018可通過靶向miR-660-3p/TEP1軸抑制HSC細胞的增殖和激活。
3.1.3circ_0070963 Ji等[25]通過circRNA芯片分析發(fā)現circ_0070963在肝纖維化過程中表達降低。研究證實circ_0070963是miR-223-3p海綿,可通過調控miR-223-3p/LEMD3軸抑制肝纖維化中HSC的激活。LEMD3含有LEM結構域的蛋白質,在核膜內具有拮抗TGF-β信號的功能[25]。circ_0070963可通過吸附miR-223-3p促進LEMD3的表達。
3.1.4circCREBBP Yang等[26]發(fā)現circCREBBP在CCl4誘導的肝纖維化小鼠模型中顯著低表達。在體內使用AAV8-circCREBBP過表達circCREBBP,可通過降低血清丙氨酸轉氨酶和天門氨酸轉氨酶含量、肝臟羥脯氨酸水平、膠原沉積以及降低促纖維化基因和促炎細胞因子水平,預防CCl4誘導的肝纖維化進展。LEFTY2是TGF-β超家族蛋白的分泌配體,circCREBBP可以通過靶向miR-1291/LEFTY2軸減輕肝纖維化。
3.1.5circPSD3 Bu等[27]提取CCl4誘導的肝纖維化小鼠的肝組織和HSC細胞,發(fā)現circPSD3在肝組織和培養(yǎng)的HSC中明顯下調。此外,體外功能喪失和功能獲得實驗表明,circPSD3抑制HSC的激活和增殖。circPSD3可作為miR-92b-3p的海綿,進而促進Smad7的表達。
3.1.6circFBXW4 Chen等[28]通過circRNA-seq獲得肝纖維化進展和衰退小鼠HSC中circRNA、miRNA和mRNA的表達譜。他們發(fā)現過表達circFBXW4可抑制HSC的活化增殖,誘導細胞凋亡,減輕小鼠肝纖維化損傷,并具有抗炎作用。circFBXW4可能通過靶向miR-18b-3p/FBXW7軸調節(jié)HSC激活和抑制纖維化;其下游靶點FBXW7可通過增加鐵死亡誘導生長抑制,并有助于HSC細胞的自噬降解功能[29]。
3.2 具有促纖維化作用的circRNA
3.2.1circRNA-0067835 Zhu等[30]發(fā)現胸腺素-β4(thymosin, Tβ4)可通過circRNA-0067835/miR-155/FoxO3信號通路調控HSC激活,他們使用circRNA微列陣鑒定Tβ4相關circRNA,發(fā)現circRNA-0067835在Tβ4缺失的LX-2細胞中表達明顯增加。FoxO3所屬的FoxO家族,可通過PI3K/Akt通路調控HSC增殖[31-32]。circRNA-0067835可作為miR-155的海綿促進FoxO3的表達,發(fā)揮促纖維化作用。
3.2.2circRNA-PWWP2A Liu等[33]發(fā)現TGF-β和脂多糖誘導的circRNA-PWWP2A,可通過海綿吸收miR-203和miR-223促進肝纖維化。該研究結果表明,circ-PWWP2A是TGF-β和脂多糖在HSC激活過程中共同的下游反應器,circRNA-PWWP2A通過海綿吸附miR-203、miR-223,并促進其下游靶基因Fstl1、TLR4釋放,在肝纖維化過程中發(fā)揮重要調控作用。Fstl1和TLR4均為促纖維化因子,在HSC激活和促進纖維生成中發(fā)揮重要作用[34-35]。
3.2.3circRSF1 Chen等[36]發(fā)現circRSF1通過調節(jié)miR-146a-5p/RAC1促進受輻射HSC的炎癥和纖維化表型。輻射后的人HSC細胞系LX-2中RAC1表達升高,并可促進LX-2細胞增殖和α-SMA表達,而這些促進過程均被miR-146a-5p模擬物阻斷。RAC1是一種GTPase,可調節(jié)細胞生長,過表達circRSF1可通過吸附miR-146a-5p增加RAC1的表達,并促進輻射后LX-2細胞的炎癥和纖維化表型。circRSF1為放射性肝損傷(radiation-inducedliverdisease, RILD)提供有吸引力的治療靶點。
3.2.4circUbe2k Zhu等[37]發(fā)現,circUbe2k可通過調控miR-149-5p/TGF-β2促進肝纖維化。在CCl4誘導的肝纖維化小鼠和受TGF-β1刺激的LX-2細胞中,circUbe2k表達增強并可調節(jié)肝纖維化的進展。抑制circUbe2k表達可降低HSC的激活和增殖,抑制CCl4誘導的肝纖維化小鼠和TGF-β1刺激的LX-2細胞中α-SMA和I型膠原alpha 1鏈(collagen type I alpha 1 chain, Col1a1)的表達。circUbe2k可以通過吸附miR-149-5p,從而導致TGF-β2升高。
4 circRNA對肝纖維化的間接調控
4.1 circRNA可能通過細胞間串擾調控肝纖維化Zhu等[38]研究證明,脂肪來源的間充質干細胞(adipose derived stromal cell, ADSC)分泌的外泌體可以用來傳遞circRNA治療肝纖維化。他們發(fā)現mmu_circ_0000623在CCl4誘導的肝纖維化小鼠中表達下調。體外和體內實驗表明,ADSC分泌的外泌體,尤其是來自mmu_circ_0000623修飾的ADSC分泌的外泌體,可通過促進自噬激活顯著抑制CCl4誘導的肝纖維化。
4.2 circRNA在肝內其他肝纖維化相關疾病中的研究近年,非酒精性脂肪性肝病(non-alcoholic fatty liver disease, NAFLD)、非酒精性脂肪性肝炎(non-alcoholic steatohepatitis, NASH)逐漸成為肝纖維化/肝硬化發(fā)生的重要誘因。Day和James提出NAFLD/NASH發(fā)生過程中的“兩擊”假說(“two-hit” hypothesis),即第一次攻擊導致肝臟脂肪堆積,第二次攻擊導致炎癥和纖維化。一些研究發(fā)現circRNA可能通過調節(jié)脂質代謝和氧化應激參與調控NAFLD的發(fā)病機制。例如circRNA_0046367可通過內源性調控miR-34a表達,消除脂肪變性中的脂質過氧化、凋亡和線粒體功能障礙,并消除miR-34a對過氧化物酶體增殖物激活受體α(peroxisome proliferator activated receptor α, PPARα)的抑制作用。circScd1可通過JAK2/STAT5途徑在NAFLD中增加肝細胞脂質沉積,從而促進脂肪肝疾病。
5 總結與展望
肝纖維化的發(fā)生機制非常復雜,肝慢性損傷后的早期炎癥反應、HSC活化和ECM異常沉積,是肝纖維化發(fā)生的關鍵級聯事件。越來越多的證據表明,肝纖維化發(fā)生過程中circRNA呈現出顯著差異性表達。這些circRNA可海綿吸附miRNA,引起miRNA下游肝纖維化相關靶基因釋放,通過調節(jié)HSC增殖和ECM沉積、調控HSC細胞的自噬降解、調節(jié)膠原基因的表達及調節(jié)促炎因子水平等,直接調控肝纖維化過程。此外,某些circRNA可能通過細胞間串擾,以及通過調節(jié)肝內脂質代謝、脂質過氧化損傷等方式對肝纖維化產生間接調控。
目前,circRNA在肝纖維化中的具體調控機制的研究仍處于早期階段,大量circRNA的功能尚未明確。現有的circRNA功能機制研究仍較為局限,且處于基礎實驗階段,尚未有臨床轉化的相關報道。因此,circRNA在HSC活化/肝纖維化發(fā)生過程中的功能機制仍有待分析,具有臨床轉化價值的circRNA仍有待于進一步挖掘。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
四川勞動保障(2021年9期)2022-01-18 05:11:08
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
文苑(2018年21期)2018-11-09 01:23:06
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國衛(wèi)生(2016年9期)2016-11-12 13:28:08
中國衛(wèi)生(2015年9期)2015-11-10 03:11:12
中國衛(wèi)生(2014年3期)2014-11-12 13:18:12
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
中國火炬(2014年4期)2014-07-24 14:22:19
