具有抗炎和促干細胞活性雙效應評價功能的新型結腸炎小鼠模型的構建及應用
2022-12-21 05:59:18鮑維廉尤科淵馮桂澤曹心悅沈曉燕
復旦學報(醫學版) 2022年6期
鮑維廉 尤科淵 馮桂澤 曹心悅 沈曉燕
(復旦大學藥學院藥理學教研室 上海 201203)
實驗和臨床證據表明,腸黏膜屏障功能受損是炎癥性腸病(inflammatory bowel disease,IBD)的驅動因素[1-3]。研究已將黏膜愈合作為IBD治療的關鍵目標,意味著患者腸黏膜結構和功能可完全修復,預示著患者臨床癥狀持續緩解和手術風險降低[4]。目前治療IBD的藥物[5-6]主要用于控制免疫炎癥,對于黏膜損傷的修復非常有限,在以黏膜愈合為主要終點的臨床試驗中,藥效仍不能令人滿意[7-9]。因此,尋找既能控制炎癥又能促進黏膜愈合的新靶標,并在此基礎上開發治療IBD的新藥物成為迫切需要解決的醫學問題。
在腸隱窩底部的Lgr5+細胞群具有成體干細胞潛能的腸干細胞(intestinal stem cell,ISC),可以在黏膜受損的情況下完成上皮系統的更新。在IBD患者中,Lgr5+ISCs的減少與疾病活動相關[10-13]。ISCs的丟失和功能異常是IBD慢性、反復發作性炎癥的重要因素[14]。而將ISCs移植到實驗性結腸炎動物的黏膜損傷部位,可以加速黏膜愈合[15-17]。Lgr5-EGFP-IRES-creERT2小鼠可以完成對ISCs的示蹤和分選。該小鼠攜帶Lgr5-EGFP-IRESCreERT2基因敲入型等位基因,破壞了內源Lgr5(Gpr49)基因功能,并表達來自Lgr5啟動子/增強子驅動的EGFP和CreERT2融合蛋白。該轉基因小鼠純合子不能存活,但是Lgr5-EGFP-IRESCreERT2雜合小鼠可存活且可繁育。在該種小鼠的腸隱窩基底柱狀細胞(即ISCs)中可觀察到EGFP熒光。Cre-ERT2融合蛋白由Cre重組酶和三重突變形式的人雌激素受體組成,該受體可與熱休克蛋白90(heat shock protein 90,HSP90)結合。由于HSP90的阻擋,雌激素受體結合的融合蛋白無法進入細胞核。雌激素和雌激素受體結合使融合蛋白擺脫HSP90,進入細胞核內發揮Cre酶活性。生理濃度下,受體與其天然配體(17β-雌二醇)不發生結合,但會與合成的雌激素受體配體4-羥基他莫昔芬或他莫昔芬結合,故可以通過注射他莫昔芬實現對Cre酶系統的控制。Lgr5-EGFP-IRES-creERT2小鼠與攜帶loxP側翼序列的小鼠雜交獲得的雙基因敲除小鼠受他莫昔芬誘導后,Cre酶介導的重組導致后代中表達Lgr5的細胞中loxP位點間的基因序列被敲除。如果沒有flox基因片段的存在,Lgr5-EGFP-IRES-creERT2僅 發 揮 對Lgr5陽 性細胞的熒光示蹤功能,其熒光強度在一定程度上可以表征小鼠腸黏膜屏障的自我修復能力。
Il10基因敲除小鼠是一種廣泛使用的結腸炎模型[18]。Il10-/-小鼠通常在8~12周齡開始發生漸進的、不連續的透壁型炎癥,炎癥從右結腸開始,隨著疾病進展向遠端延伸,可累積小腸;同時出現明顯的IBD癥狀,如體質量下降、稀便、便血、脫肛等。發病機制與人類IBD相似,病情呈漸進式發展,可用于早期干預研究,是研究IBD的理想模型。
本研究中,我們用Il10-/-小鼠和Lgr5-EGFP小鼠雜交得到Il10-/-;Lgr5-EGFP小鼠。該基因型小鼠經誘導形成慢性結腸炎模型,可以同時評價藥物對炎癥活動和干細胞活性的影響。成功建立這一模型可為結腸炎潛在藥物/治療手段的體內實驗提供一個可靠的雙靶點驗證的動物模型。
材料和方法
實驗動物Lgr5-EGFP-IRES-creERT2(Lgr5-EGFP)和B6.129P2-Il10tm1Cgn/J(Il10-/-)小鼠均引進自美國Jackson實驗室,引進后擴群繁殖,并將兩種基因型小鼠雜交繁殖。繁殖過程中所有小鼠均飼養于復旦大學藥學院實驗動物中心SPF級動物房,實驗動物使用許可證號:SCXK(滬)2014-0002。溫度(25±2)℃,濕度50%~60%,無菌飲水,自由進食和飲水,保持12 h光-暗循環。經鑒定得到目標基因型小鼠Il10-/-;Lgr5-EGFP及其對照基因型小鼠WT;Lgr5-EGFP。進行體內實驗的小鼠均為雄性,8~10周齡,體質量18~22 g,實驗過程中飼養條件同上。本研究通過復旦大學藥學院動物倫理委員會審批(批準號:2019-03-YL-SXY-01)。
小鼠的基因鑒定取3周齡小鼠,按性別分籠并剪鼠耳標記,剪3~5 mm鼠尾,提取DNA,對小鼠基因型進行PCR鑒定。對于Lgr5-EGFP,根據待測樣 品 數,將 表1中 的3種 引 物 以 每 份0.6、0.5和0.8 μL的體積混合,加入5×樣本體積的2×PCR Mix和1.1×樣本體積的DEPC水,制成鑒定工作液。將8 μL鑒定工作液加入2 μL DNA樣本中,得到完整的鑒定反應體系,置入PCR儀中,按照表2中的程序進行擴增反應,得到鑒定產物1。對于Il10,根據待測樣品數,將表3中的3種引物以每份1.0、0.5和0.5 μL的體積混合,加入5×樣本體積的2×PCR Mix和1×樣本體積的DEPC水,制成鑒定工 作液。將8 μL鑒定 工 作液 加 入2 μL DNA樣 本中,得到完整的鑒定反應體系,置入PCR儀中,按照表4中的程序進行擴增反應,得到鑒定產物2。對鑒定產物1和2進行1%瓊脂糖凝膠電泳,根據最終條帶所對應的Marker判斷分子量,進而判斷基因型。每只小鼠的DNA樣本將進行3次以上獨立的PCR及凝膠電泳,確保結果可靠。
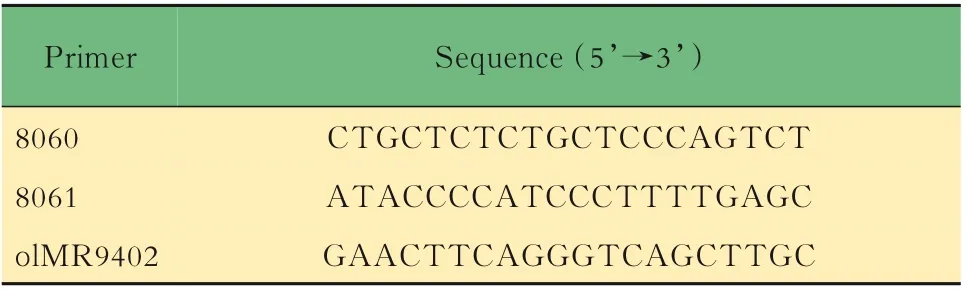
表1 Lgr5-EGFP小鼠DNA鑒定引物序列Tab 1 Primers for DNA identification of Lgr5-EGFP mice
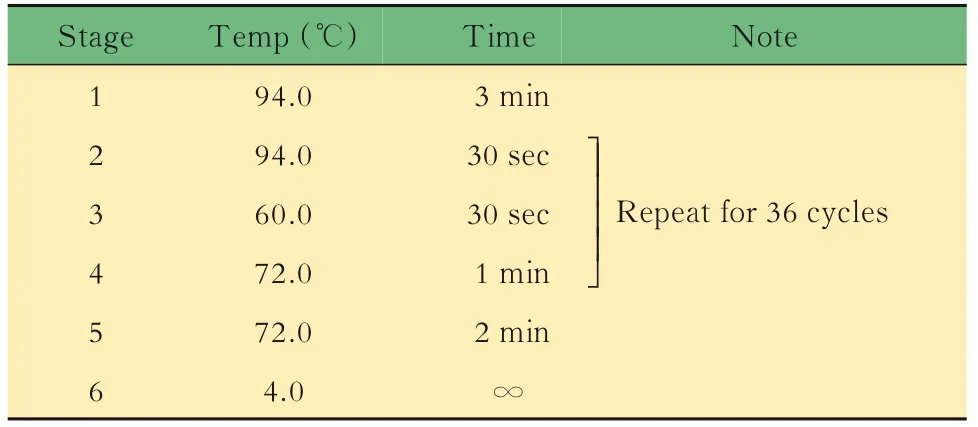
表2 Lgr5-EGFP小鼠DNA鑒定的擴增反應Tab 2 Amplification reaction for DNA identification of Lgr5-EGFP mice
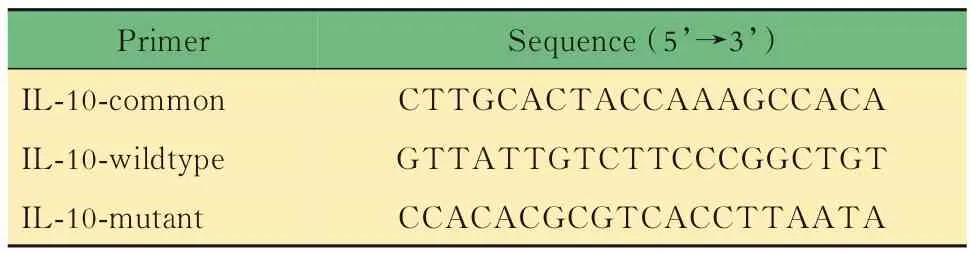
表3 Il10-/-小鼠鑒定PCR引物Tab 3 Primers for DNA identification of Il10-/-mice
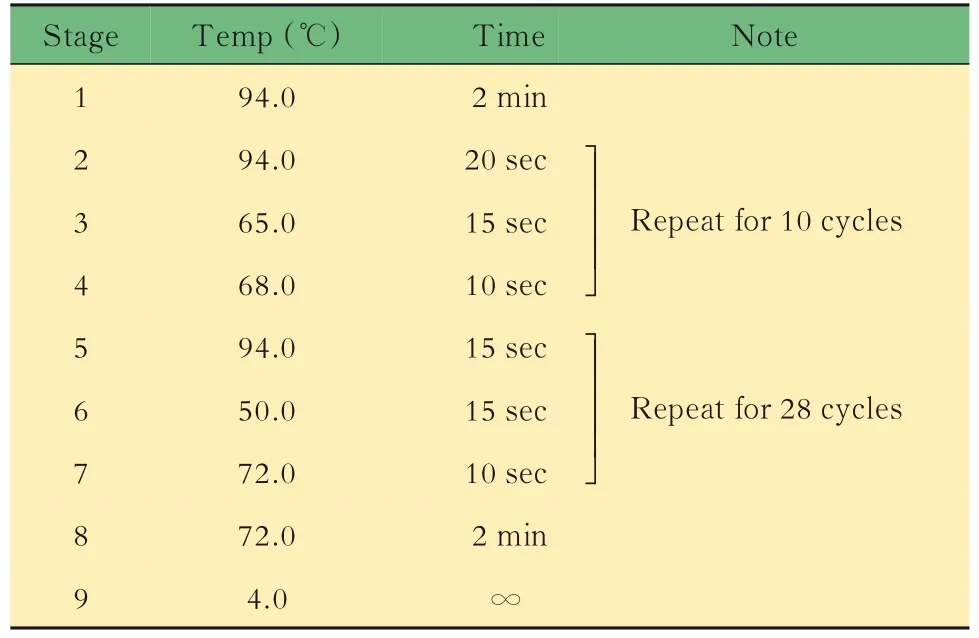
表4 Il10-/-小鼠鑒定的擴增反應Tab 4 Amplification reaction for DNA identification of Lgr5-EGFP mice
體內實驗取6只8周齡雄性WT;Lgr5-EGFP基因型小鼠作為對照組;Il10-/-;Lgr5-EGFP基因型小鼠共計18只,采用隨機數字表法分為3組:模型組、美沙拉嗪低劑量組和高劑量組,每組6只。模型組及給藥組給予飲用水配制的3% DSS溶液4天,在SPF環境下誘導結腸炎,每2天換一次DSS溶液,對照組小鼠正常飲水。美沙拉嗪采用0.5%羧甲基纖維素鈉配置成混懸液,發病后開始灌胃給藥,低劑量組為50 mg·kg-1·d-1,高劑量組為150 mg·kg-1·d-1,每周記錄小鼠體質量,觀察并記錄小鼠稀便、血便、脫肛和體質量減輕等情況,根據表5進行評分。7周后實驗結束,分離各實驗樣本,進行檢測。
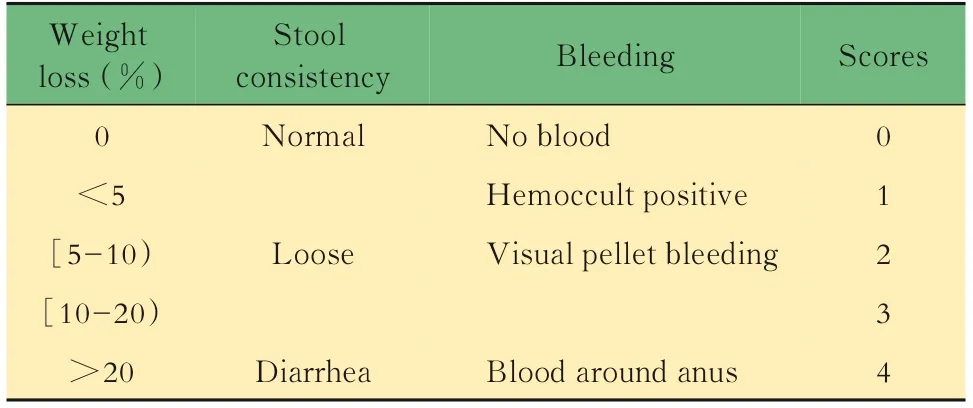
表5 疾病活動指數評分標準Tab 5 Criteria of disease activity index scoring
ELISA檢測小鼠炎癥因子使用水合氯醛麻醉小鼠,壓迫眼球,使眼球突出,用鑷子或止血鉗迅速摘除眼球并處死。獲得的外周血在室溫靜置2 h后,4℃下3 000×g離 心15 min,上 清 即 為 血 清,-80℃冰箱儲存。使用商品化的ELISA試劑盒檢測炎癥因子,并根據制造商提供的說明書進行實驗。通用步驟按濃度梯度稀釋標準品;根據實驗孔(包括空白和標準品)數量,確定所需的板條數目;每孔加入不同稀釋度的標準品或待測樣品100 μL,空白孔為稀釋液;加入一抗工作液或直接用膜封板,37℃孵育2 h;加入洗滌液洗滌,在濾紙上倒扣去除殘留液體,重復3次。加入酶標記二抗,37℃孵育30 min;每孔加入漂洗液150 μL,洗板,扣去殘留液體,重復3次;加入現配的TMB顯色液,37℃孵育10~30 min,加入終止液,停止反應;反應終止后10 min內,用酶標儀在對應波長處測得OD值;根據稀釋標準品擬合標準曲線,將待測樣品吸光度值代入標準曲線,計算樣品濃度。
RT-qPCR檢測小鼠結腸組織mRNA分離獲得小鼠結腸樣本,用無菌PBS沖洗結腸,去除腸腔內糞便。剪碎結腸組織,用組織勻漿機打碎,加入Trizol,離心取上清,室溫靜置10 min。向完成裂解的樣本中加入200 μL(Trizol體積的1/5)氯仿,劇烈振搖15 s,靜置2~3 min;分層后,4℃下12 000×g離心15 min,吸取最上層水相(400~500 μL),轉移至無RNase的EP管中。加入等體積(400~500 μL)異丙醇,輕柔顛倒,靜置10 min,可見白色絮狀物。4℃下2 000×g離心10 min,棄去上清。加入1 mL DEPC水和無水乙醇現配的75%乙醇,槍頭吹打混勻,洗滌沉淀,7 500×g離心5 min,棄上清,用移液槍吸盡殘留液體。通風櫥內室溫干燥5~10 min,根據沉淀體積加入適量DEPC水溶解RNA。使用Nano Drop 2000(美 國ThermoFisher公 司)檢 測RNA濃度,調整各樣本至同一濃度范圍。使用逆轉錄試劑盒和SYBR-qPCR試劑盒[翌圣生物科技(上海)股份有限公司]進行RT-qPCR檢測炎癥因子的表達,qPCR的引物如表6所示。
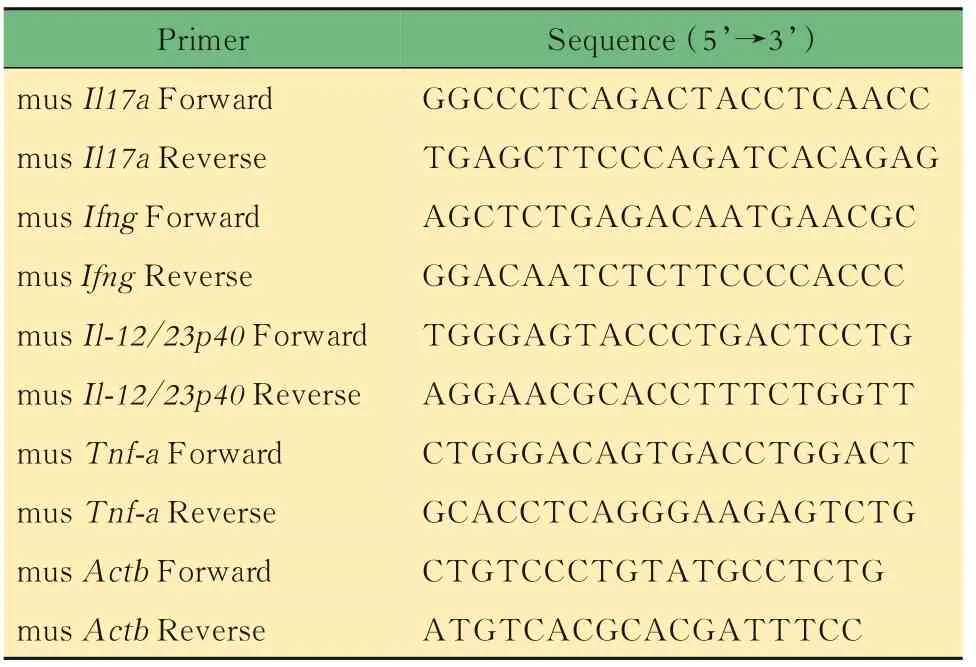
表6 qPCR引物序列Tab 6 Primer sequences for qPCR
冰凍切片的制作和DAPI染色去除小鼠結腸的內糞便,置于4%多聚甲醛中固定24 h以上。將組織先后置于15%和30%的蔗糖溶液內,在4℃冰箱中脫水。濾紙濾干脫水的組織,將目的部位修平,切面朝上放于樣本托中,OCT包埋劑滴至周圍,置于冰凍切片機的速凍臺上,直至OCT凝固,切片,切片厚度8~10 μm,于-20℃保存。載玻片用PBS(pH=7.4)洗滌3次。晾干,DAPI染色10 min,滴加含抗熒光淬滅劑的封片劑,封片。于Zeiss 710熒光共聚焦顯微鏡下觀察并采集圖像。
統計學處理用GraphPad Prism 8.0軟件對數據進行統計學分析及制圖,所有數據用±s表示。血清炎癥因子、結腸炎癥因子mRNA表達和結腸部位Lgr5+ISCs觀察的定量數據比較采用單因素方差分析,并采用Bonferroni法進行組間比較;小鼠體質量比較采用兩因素重復測量方差分析,并采用Bonferroni法進行組間比較;小鼠疾病活動指數的比較采用Kruskal-Wallis分析,并采用Dunn’s法進行組間比較。P<0.05為差異有統計學意義。
結 果
Il10-/-;Lgr5-EGFP小鼠基因鑒定對小鼠鼠尾提取的基因組DNA,使用特定的引物序列進行PCR擴增,并對產物進行瓊脂糖凝膠電泳。根據所示條帶(圖1)判斷小鼠的基因型。Lgr5-EGFP基因在174和298 bp處都有清晰條帶,為Lgr5-EGFP雜合子小鼠。Il10基因僅在137 bp處有清晰條帶,為WT小鼠;Il10基 因 僅 在312 bp處有清晰條帶,為Il10-/-純合子小鼠。①~⑥號小鼠的基因鑒定結果(圖1):③號(泳道3和9)和⑤號(泳道5和11)為WT;Lgr5-EGFP小鼠,②號(泳道2和8)為Il10-/-;Lgr5-EGFP小鼠。
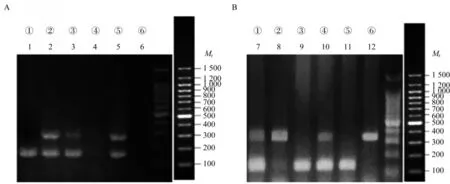
圖1 小鼠基因型鑒定Fig 1 Genotype identification of mice
美沙拉嗪有效緩解Il10-/-;Lgr5-EGFP模型的體質量下降和疾病評分對Il10-/-;Lgr5-EGFP小鼠進行持續4天的3% DSS誘導,可在SPF環境下形成慢性結腸炎模型。為驗證該模型,我們設置了WT;Lgr5-EGFP小鼠的對照組和長期給予50或150 mg·kg-1·d-1美沙拉嗪的治療組。實驗過程中發現,正常對照組小鼠體質量平穩,飲食、飲水正常,精神狀態佳,活動頻繁,糞便形態未見明顯異常;以開始給予DSS誘導為第0周,模型組小鼠在誘導后前2周體質量驟降,第3周體質量略有恢復,第4周開始體質量再次持續下降,飲食少,食欲差,精神狀態萎靡,出現腹瀉、肉眼血便、膿便等癥狀。而相比于模型組,治療組體質量下降減緩,小鼠稀便和血便情況明顯減少,各項癥狀得到明顯改善。結果顯示(圖2),在體質量變化和疾病活動評分上,全程模型組和對照組都存在顯著差異;美沙拉嗪低劑量組在第2、5、6和7周體質量降低顯著改善,在第2、6和7周疾病活動評分顯著改善;美沙拉嗪高劑量組在第1、2、4、5、6和7周體質量降低顯著改善,在第2~7周疾病活動評分顯著改善。
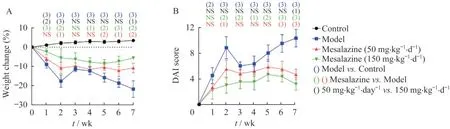
圖2 DSS誘導的Il10-/-;Lgr5-EGFP小鼠體質量和疾病活動指數變化Fig 2 Body weight and disease activity index changes of DSS-induced Il10-/-;Lgr5-EGFP mice
美沙拉嗪有效抑制Il10-/-;Lgr5-EGFP模型的炎癥因子的表達和組織病變對小鼠的血清炎癥因子進行ELISA檢測(圖3),和對照組相比,模型組小鼠血清中IL-12/23 p40、IL-17A、TNF-α和IFN-γ等炎癥因子含量顯著增加(P<0.001)。50和150 mg·kg-1·d-1的美沙拉嗪均可顯著抑制Il10-/-;Lgr5-EGFP模型組血清中各炎癥因子水平(P<0.001)。150 mg·kg-1·d-1美沙拉嗪對血清中IL-12/23 p40和TNF-α的 抑 制效果優于50 mg·kg-1·d-1美沙拉嗪(P<0.01)。
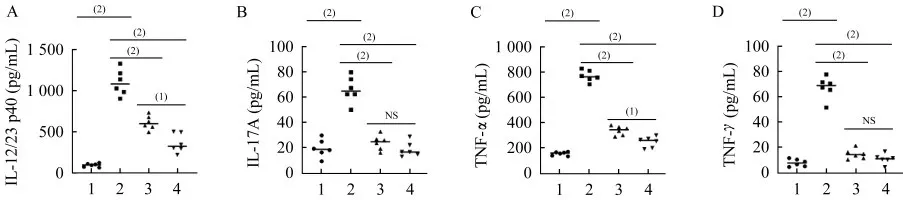
圖3 DSS誘導的Il10-/-;Lgr5-EGFP小鼠血清炎癥因子Fig 3 Inflammatory cytokines in serum of DSS-induced Il10-/-;Lgr5-EGFP mice
我們通過RT-qPCR定量檢測了各組小鼠結腸組織中炎癥因子的mRNA表達水平。和對照組相比,模型組小鼠結腸組織中Il12/23 p40、Il17a、Tnf-a和Ifng的mRNA表達水平顯著上升(P<0.001),50和150 mg·kg-1·d-1美沙拉嗪均可顯著抑制模型組各炎癥 因子的mRNA表達水平,150 mg·kg-1·d-1美沙拉 嗪 對Il12/23 p40、Tnf-a和IfngmRNA表 達 的 抑制作用顯著優于50 mg·kg-1·day-1美沙拉嗪(圖4)。
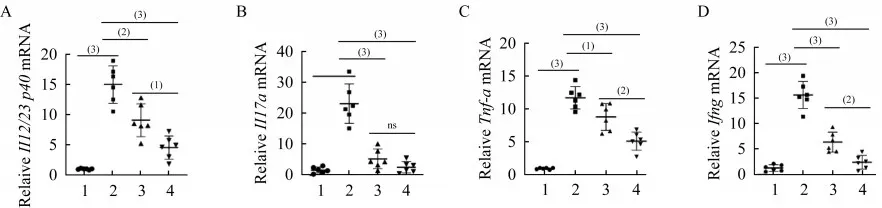
圖4 DSS誘導的Il10-/-;Lgr5-EGFP小鼠結腸炎癥因子mRNA表達Fig 4 Colonic inflammatory cytokine mRNA expression of DSS-induced Il10-/-;Lgr5-EGFP mice
病理組織學觀察結果顯示,模型組小鼠結腸出現嚴重潰瘍、隱窩結構破壞和嚴重的炎癥細胞浸潤,而美沙拉嗪組的炎癥細胞浸潤明顯減少,有大量完整隱窩結構保留,表明該模型引起的慢性炎癥對上皮屏障產生了明顯破壞,且可被陽性藥緩解。
這些結果表明,Il10-/-;Lgr5-EGFP模型小鼠具有明顯的炎癥和上皮屏障損失的表型,且可以被美沙拉嗪顯著抑制。
美沙拉嗪有效挽救Il10-/-;Lgr5-EGFP模型的ISCs丟失對小鼠結腸的冰凍切片進行DAPI染色,在熒光共聚焦顯微鏡下觀察Lgr5-EGFP的表達情況。對照組小鼠結腸隱窩底部存在一定數量的Lgr5-EGFP陽性細胞,而模型組小鼠結腸隱窩底部的Lgr5-EGFP陽性細胞數明顯減少(圖5)。美沙拉嗪可以在一定程度上恢復小鼠結腸隱窩底部的Lgr5-EGFP陽性細胞數量。這些結果表明,Il10-/-;Lgr5-EGFP模型表現出類似CD患者Lgr5+ISCs丟失的表型,而美沙拉嗪可以修復ISCs丟失(圖6)。
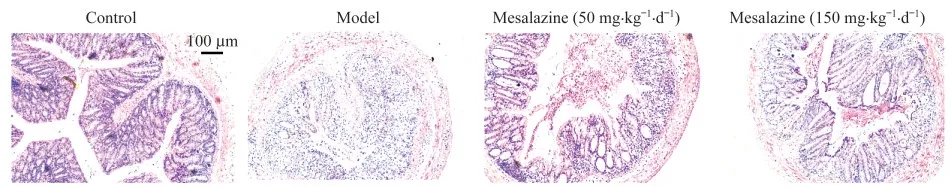
圖5 DSS誘導的Il10-/-;Lgr5-EGFP小鼠結腸切片的HE染色圖片Fig 5 HE staining images of colon sections in DSS-induced Il10-/-;Lgr5-EGFP mice
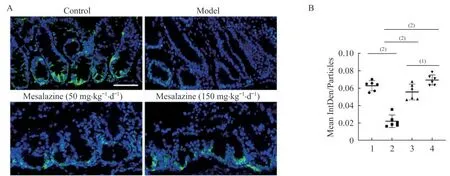
圖6 DSS誘導的Il10-/-;Lgr5-EGFP小鼠結腸部位Lgr5+ISCs觀察Fig 6 Observation of Lgr5+ISCs in colon of DSS-induced Il10-/-;Lgr5-EGFP mice
討 論
ISCs在上皮屏障恢復過程中扮演著重要角色,被認為是炎性腸病潛在治療靶點[10-16]。目前對于ISCs在IBD中作用的研究采用的是7天DSS誘導的急性結腸炎模型。急性DSS結腸炎更適用于研究上皮破壞后黏膜內穩態的暫時性喪失以及初始損傷后黏膜愈合的機制,但該模型并不能體現人類IBD特有的慢性和復發性特征,其作為IBD藥物篩選模型的合理性仍存在爭議[19]。因此,我們選擇了和人類IBD更為接近的Il10-/-小鼠和干細胞示蹤的Lgr5-EGFP小鼠雜交,獲得可以通過綠色熒光蛋白觀察ISCs數量的慢性結腸炎模型小鼠。在SPF級飼養環境中,每只Il10-/-小鼠自發出現結腸炎表型的周齡并不相同,這可能會提升其作為藥物評價模型的不可控性。因此,我們對相同周齡的Il10-/-小鼠采用3% DSS連續誘導4天,造成小鼠腸黏膜的初始損傷,之后恢復正常飲水,使小鼠產生持續的慢性結腸炎。我們觀察到在誘導7周后,Il10-/-小鼠出現明顯的Lgr5+ISCs耗竭,這和在人類CD中觀察到的ISCs減少相一致[10]。但是,我們也發現,不經DSS初始誘導的Il10-/-小鼠會自發產生輕度結腸炎癥狀,并出現明顯的隱窩增生異常。這種上皮細胞過度增殖的表現提示,IL-10缺失在上皮屏障未遭到破壞的情況下,可能并不會造成ISCs耗竭。這種模型上的差異同樣體現在藥物干擾上。有研究報道,美沙拉嗪對于DSS誘導的大鼠腸炎模型中的Lgr5+ISCs并沒有顯著的保護作用[20]。而在本模型中,美沙拉嗪有效促進了Lgr5+ISC的恢復。推測原因可能是:DSS模型的作用機制源于DSS對上皮細胞的毒性,進而破壞黏膜屏障,導致共生菌的易位,引發急性炎癥;而本模型盡管使用了DSS作為初始誘導劑,但之后形成的慢性腸炎更接近IL-10缺失導致的免疫缺陷型腸炎,主要由Treg下調導致的免疫耐受受損引起[21]。有研究指出,美沙拉嗪通過芳香烴受體增加結腸中TGF-β1活性形式的水平,從而促進CD4+Treg細胞分化[22],且TGF-β1已經被證明是ISCs自我更新的必要信號途徑[23]。這樣就解釋了美沙拉嗪在Treg缺失主導Il10-/-小鼠模型表現出更好的ISCs恢復效果。
綜上所述,我們開發了一種具有炎癥免疫和ISCs雙靶向效應評價功能的新型炎癥性腸病小鼠模型,并用美沙拉嗪治療證明了該模型的可行性,對于篩選新型IBD藥物具有重要意義。但本文在對照組設置上存在一些缺陷,缺少野生型小鼠的DSS誘導組作為對照。據文獻報道,野生型小鼠給予DSS足以誘導急性結腸炎模型,但會在2周內開始逐步恢復至接近正常體質量水平,理論上來說,不會像Il10-/-小鼠給予DSS模型一樣形成慢性炎癥。出于完整的實驗設計考慮,應當設置一組野生基因型小鼠給予4天DSS誘導作為完整模型的對照。
作者貢獻聲明鮑維廉實驗設計和執行,數據收集和分析,論文撰寫和修改。尤科淵,馮桂澤,曹心悅實驗執行,數據收集。沈曉燕論文構思和審閱,基金申請,技術支持。
利益沖突聲明所有作者均聲明不存在利益沖突。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
數學物理學報(2020年2期)2020-06-02 11:29:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
