基于nrDNA和cpDNA序列的甘肅省4種肉蓯蓉種質資源分子鑒定
2022-12-28 15:25:42康舒淇徐麗張明惠陳博晉玲朱田田
中國中醫藥信息雜志 2022年12期
康舒淇,徐麗,張明惠,陳博,晉玲,2,3,朱田田,2,3
1.甘肅中醫藥大學,甘肅 蘭州 730000;2.西北中藏藥省部共建協同創新中心,甘肅 蘭州 730000;3.甘肅省珍稀中藥資源評價與保護利用工程研究中心,甘肅 蘭州 730000
肉蓯蓉為列當科肉蓯蓉屬植物肉蓯蓉Cistanche deserticolaY.C.Ma和管花肉蓯蓉Cistanche.tubulosa(Schenk)Wight的干燥帶鱗葉肉質莖[1]。全球約有22種肉蓯蓉屬植物,多分布于北半球歐亞溫暖的沙漠、荒漠等干燥地帶。據《中國植物志》記載,中國的肉蓯蓉屬植物主要有5種,包括肉蓯蓉C.deserticolaY.C.Ma、管花肉蓯蓉C.tubulosa(Schenk)Wight、鹽生肉蓯蓉Cistanche salsa(C.A.Mey.)G.Beck、沙蓯蓉Cistanche sinensisG.Beck及蘭州肉蓯蓉Cistanche.lanzhouensisZ.Y.Zhang,主要分布在新疆、內蒙古、甘肅等西北部地區[2]。肉蓯蓉不僅具有較高的藥用價值,因其有利于沙漠治理和土壤改善,也具有較高的生態效益[3]。甘肅省作為2020年版《中華人民共和國藥典》規定正品肉蓯蓉的主要分布省份之一,野生蘊藏量和人工種植面積的合理規劃和采收尤為重要[4]。隨著經濟發展及國民健康需求的不斷增加,肉蓯蓉需求量日益增長,通過對甘肅肉蓯蓉資源現狀進行實地調查[5],發現在皋蘭、靖遠等多個地區仍然存在對野生肉蓯蓉的大規模、滅絕式采挖現象,致使野生資源受到嚴重破壞。非藥典種在民間大量使用的現象時有發生,大量偽品肉蓯蓉進入藥市[6],種質混亂勢必造成商品肉蓯蓉質量參差不齊,嚴重影響其經濟價值,因此對不同種質的肉蓯蓉進行準確鑒定尤為重要。
分子鑒定技術因樣品用量少、速度快、準確性高,已被廣泛用于動植物的物種鑒定[7],而當前對甘肅肉蓯蓉的種質資源分子鑒定研究還未見報道。本研究以核基因片段nrDNA的ITS和CD序列、葉綠體基因片段cpDNA的matK和rbcL序列為研究對象,對甘肅省不同種質肉蓯蓉進行分子鑒定,以期尋找適合肉蓯蓉分類的基因序列,并探究種質內與種質間的差異,實現肉蓯蓉種質的快速準確鑒別,以此探討肉蓯蓉種質間的分類地位,為甘肅肉蓯蓉資源的評價、保護和利用提供參考依據。
1 儀器與試藥
OHAUS Corporation DV SERIES型電子天平,奧豪斯儀器有限責任公司;DK-98型數顯恒溫水浴鍋,天津市泰斯特儀器有限公司;TGL-16M型高速離心機,湖南湘儀實驗儀器開發有限公司;G1000改進型基因聚合酶鏈式反應(PCR)儀,杭州博日科技有限公司;4100型凝膠成像處理系統,上海天能科技有限公司;Vortex-Genie 2T型渦旋混勻器,德國IKA公司;BCD-451WDEMU1型冰箱,青島海爾股份有限公司。
NEP003-2型植物基因組提取試劑盒(批號B00B00103),北京鼎國昌盛科生物技術有限責任公司;β-巰基乙醇(批號20191103)、Agarose瓊脂糖(批號0000590598)、BSA(批號20191203)、引物、Taq DNA Polymerase(批號W9910e)、dNTPs(批號R6409)、GoldView核酸染料(批號321523BB)等PCR試劑,上海生工生物工程股份有限公司;DNA Marker(批號20190601),大連寶生物股份有限公司;乙醇、氯仿、異戊醇、異丙醇為分析純,天津市大茂化學試劑廠;水為屈臣氏純凈水。
肉蓯蓉樣品共27份,多數為自采,部分為市場購買,其中肉蓯蓉(C.deserticola)16份、沙蓯蓉(C.sinensis)6份、鹽生肉蓯蓉(C.salsa)4份、蘭州肉蓯蓉(C.lanzhouensis)1份。經甘肅中醫藥大學晉玲教授鑒定為以上基原肉蓯蓉的肉質莖,所有樣品采集后于干燥硅膠中保存,樣品來源信息見表1。

表1 肉蓯蓉樣品來源信息
2 方法與結果
2.1 肉蓯蓉基因組DNA提取與質量檢測
用75%乙醇擦拭樣品表面后置于2 mL離心管中,粉碎成細粉。采用植物基因組DNA快速抽提試劑盒提取樣品的基因組DNA,提取操作方法參照試劑盒說明書。由于肉蓯蓉藥材多糖含量較高,對DNA提取有一定干擾,故在此方法基礎上用75%乙醇純化DNA 3~4次。提取的基因組DNA于-20℃冰箱中保存。采用電泳1%瓊脂糖凝膠,1×TAE緩沖液,于電泳儀(220 V,50 mA,30 min)進行基因組DNA完整性檢測。電泳完成后,置于凝膠成像儀下觀察,選取條帶清晰明亮,無明顯拖尾雜帶的DNA用于后續分析。
2.2 PCR擴增及測序
2.2.1 引物信息
在查閱文獻[8-10]的基礎上進行篩選,選用擴增成功率較高的4對引物:ITS2-ITS3、CD1F-CD1R、matK3F-matK1R和rbcL1F-rbcL724R。引 物 信 息見表2。
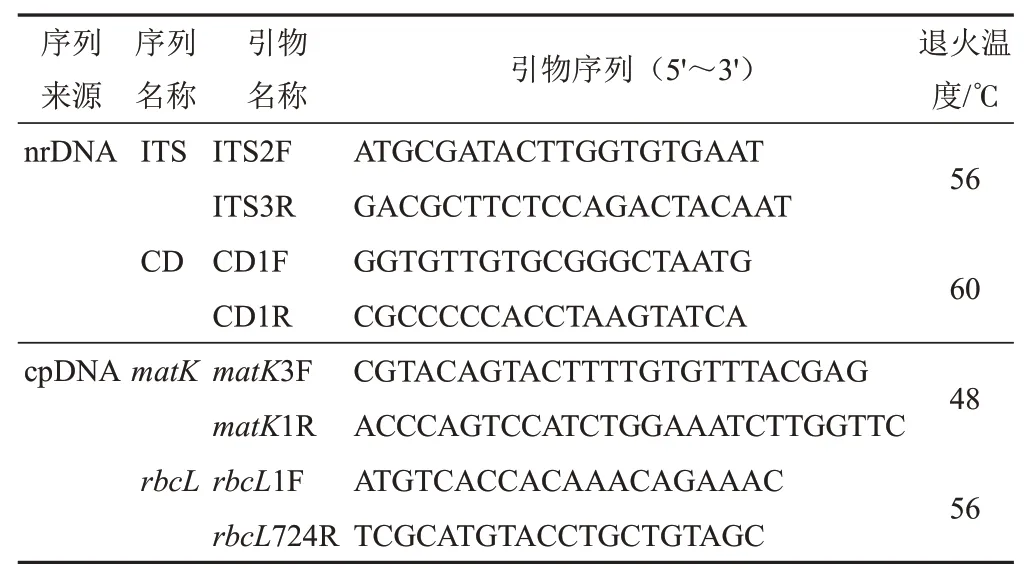
表2 肉蓯蓉基因組序列PCR引物信息
2.2.2 反應體系與檢測
PCR擴增總體系為25μL:10×PCR Buffer 2.5μL,dNTPs(10 mmol/L)2.0μL,BSA 0.5μL,TaqDNA聚合酶0.5μL,正向及反向引物各0.5μL,DNA模板2μL,ddH2O 15.5μL。PCR擴增程序為:95℃預變性5 min,94℃變性30 s,根據不同引物的退火溫度復性30 s,72℃延伸45 s,35個循環,72℃延伸7 min,4℃保存。27份肉蓯蓉樣品經4對引物擴增后電泳檢測,其擴增條帶清晰明亮,擴增成功率均為100%(見圖1)。將符合測序條件的PCR產物送上海生工生物工程股份有限公司進行雙向測序。
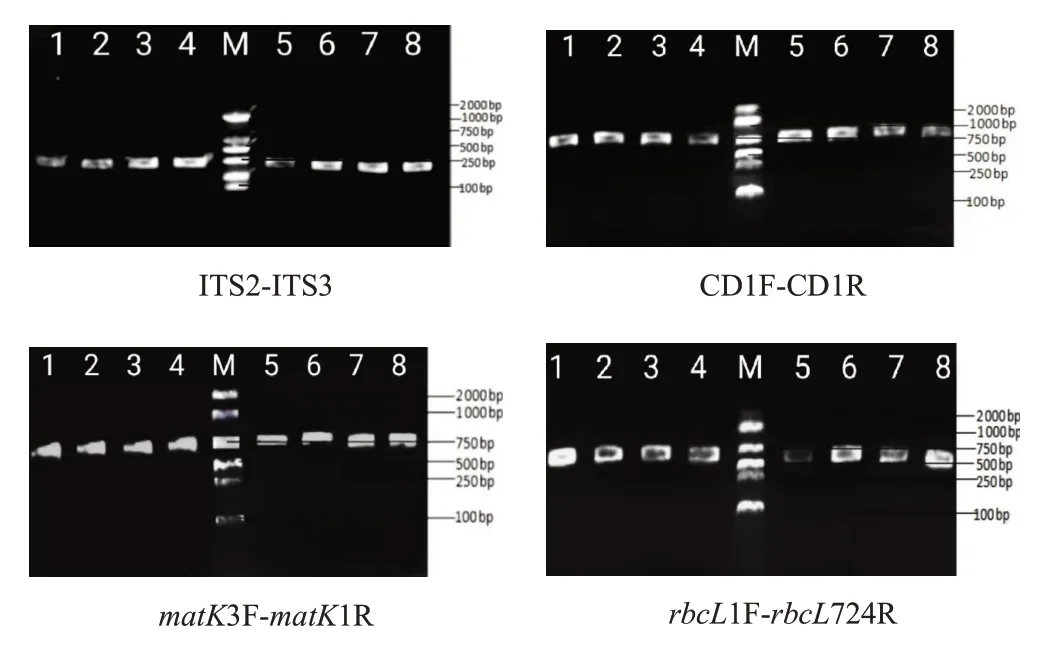
圖1 肉蓯蓉基因組序列部分PCR產物電泳
2.3 結果分析
2.3.1 樣品測序結果
將測序所得序列文件用Chromas和ContigExpress軟件去除兩端不穩定信號的低質量堿基,用以校對峰圖,并將每個樣品測序后的正反向序列拼接[11],對拼接后的每條序列峰圖進行檢查及人工校對,將處理得到的樣品序列及采集信息上傳至GenBank數據庫。ITS2F-ITS3R序列登錄號為OL827059-OL827085,CD1F-CD1R序列登錄號為OL827086-OL827112,matK3F-matK1R序列登錄號為OL827113-OL827139,rbcL1F-rbcL724R序列登錄號為OL827140-OL827166(見表3)。
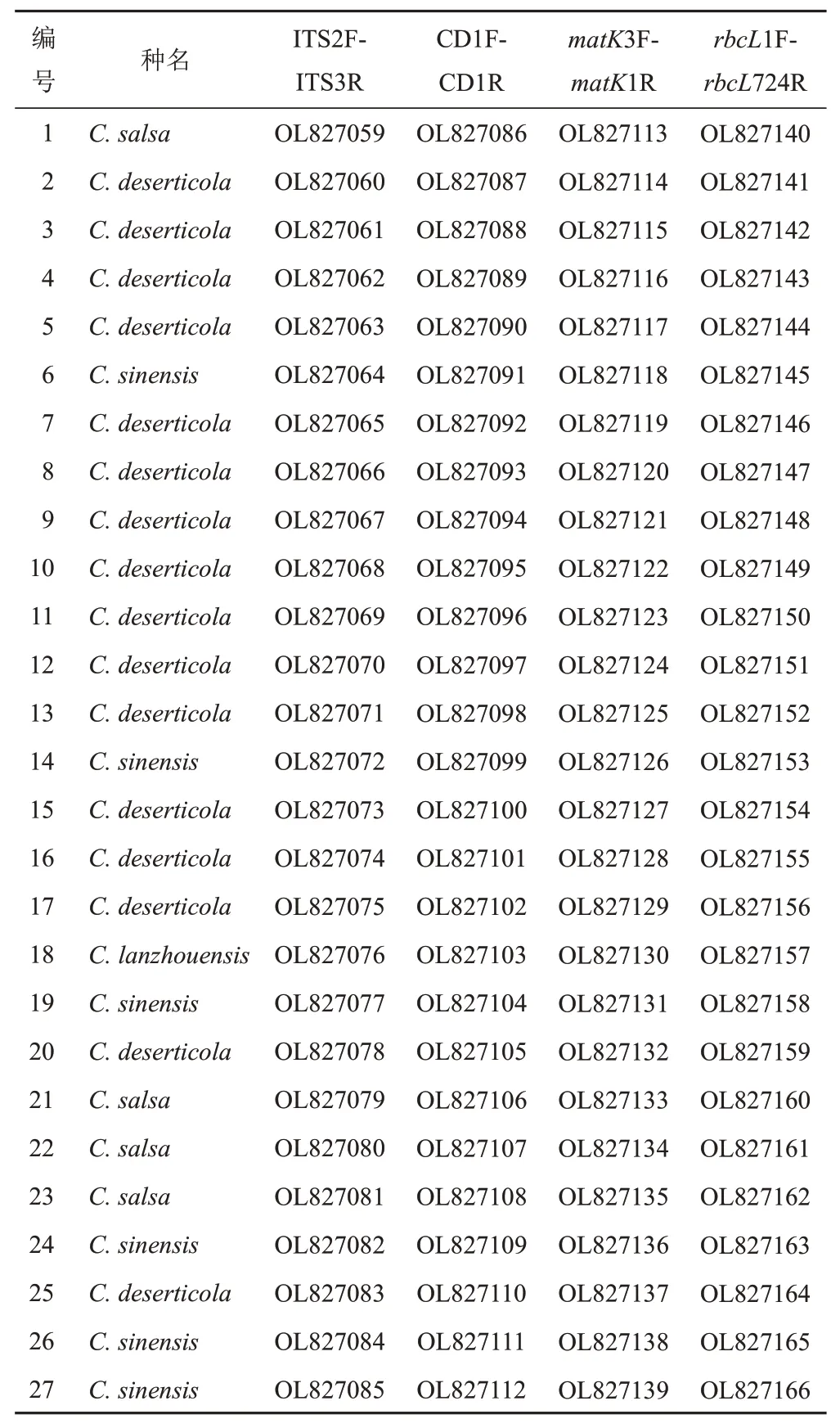
表3 肉蓯蓉樣品序列GenBank登錄號
2.3.2 肉蓯蓉序列特征
27份肉蓯蓉樣品均被成功擴增和測序,使用Mega X軟件[12]中ClustalW程序將4對引物擴增出的序列進行比對,再分別將nrDNA和cpDNA序列聯合得到2組合并序列的序列特征(見表4)。4對序列的GC含量分別為56.8%、56.9%、30.3%和41.3%,而cpDNA合并序列GC含量比nrDNA合并序列低,為34.6%。
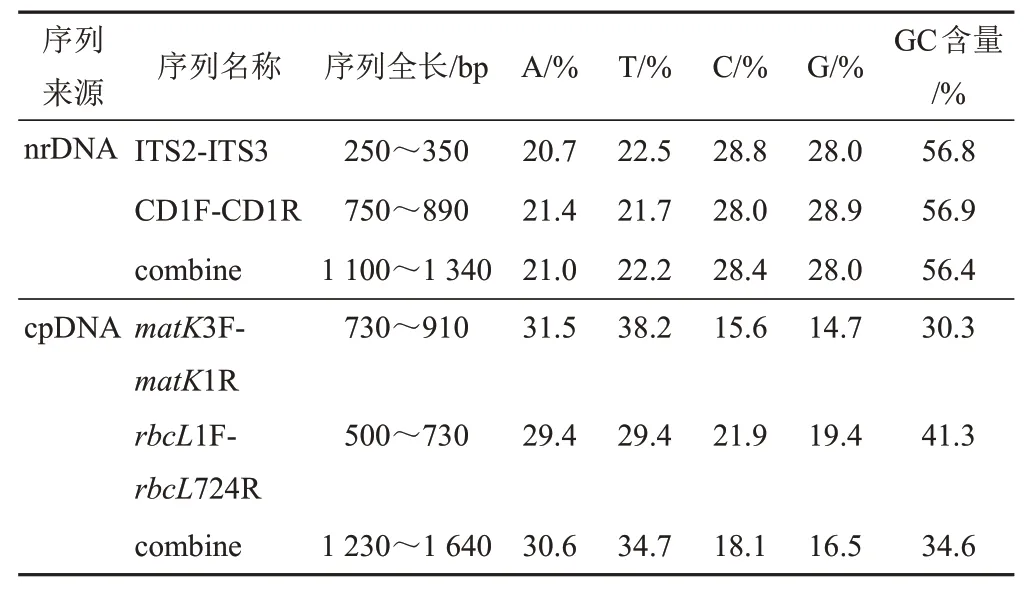
表4 肉蓯蓉樣品序列特征
2.3.3 核苷酸多態性
運用DanSP5.10.01軟件[13]進行核苷酸多態性分析,肉蓯蓉樣品在ITS、CD、matK和rbcL序列的堿基位點中存在著不同程度的變異,其中,matK序列的變異位點、單一突變位點、簡約信息性位點、插入-缺失位點及平均核苷酸差異數均明顯高于其余3對序列,共檢測到481個變異位點、369個單一突變位點、112個簡約信息位點和88個插入-缺失位點,平均核苷酸差異數為62.761。cpDNA合并序列的變異位點、單一突變位點、簡約信息性位點、插入-缺失位點及平均核苷酸差異數均明顯高于nrDNA合并序列,共檢測到534個變異位點、388個單一突變位點、146個簡約信息位點和120個插入-缺失位點,平均核苷酸差異數為77.276,但其核苷酸多樣性低于nrDNA合并序列(見表5)。
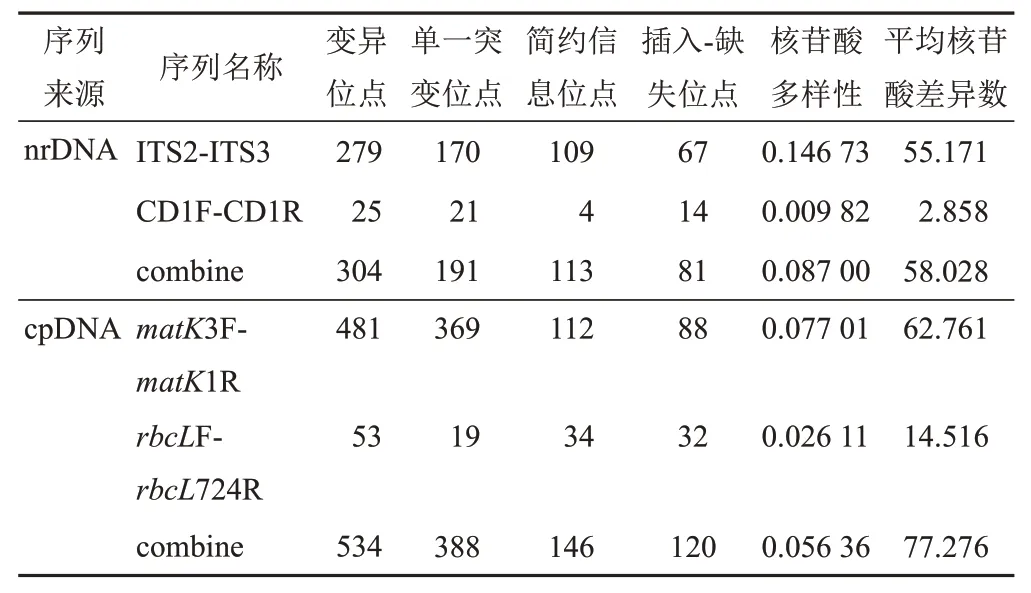
表5 肉蓯蓉樣品核苷酸多態性信息
2.3.4 單倍型多樣性
根據DanSP5.10.01軟件分析結果,發現肉蓯蓉樣品的4對序列和2組合并序列的單倍型多樣性均大于0.5,值越大,則可推斷多樣性程度越高。在4對序列中,matK序列中單倍型數量最多,共形成21個單倍型;2組合并序列中,cpDNA序列的單倍型多樣性更高,其單倍型數量達到24個(見表6)。
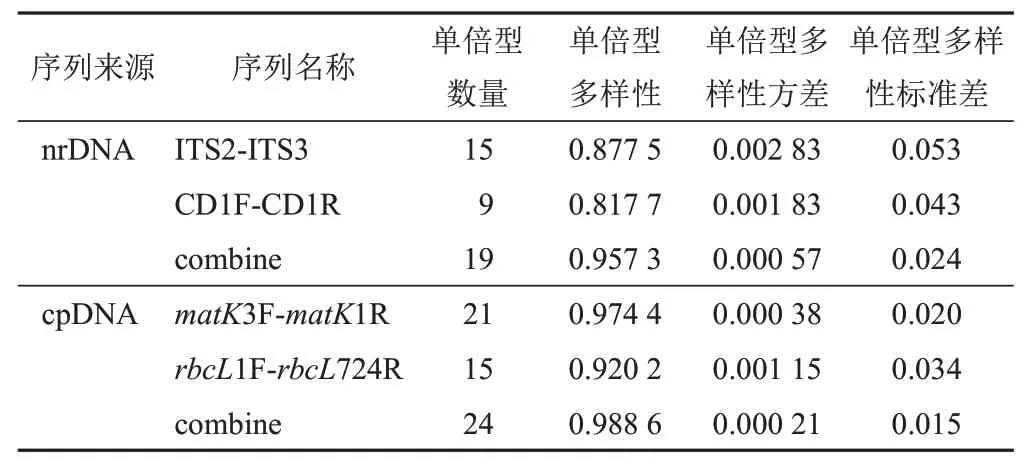
表6 肉蓯蓉樣品單倍型多樣性
2.3.5 種質遺傳距離分析
基于kimura2-parameter(K2P)模型計算肉蓯蓉ITS、CD、matK和rbcL4對序列的種質遺傳距離(見表7)可知,4對序列的平均遺傳距離為0.209 3、0.010 8、0.109 4、0.027 1,經比較其遺傳距離ITS>matK>rbcL>CD。4對序列得到的所有樣品遺傳距離范圍分別為0~1.405 6、0~0.064 1、0~0.992 5、0~0.077 3。對比可得,ITS和matK序列的變異性較大,CD和rbcL序列較保守。此外,計算遺傳距離均顯示蘭州肉蓯蓉與肉蓯蓉的遺傳距離最近,使用matK序列通過GraphPad Prism 8.0.1軟件繪制遺傳距離矩陣熱圖,遺傳距離為0.001 2(見圖2)。
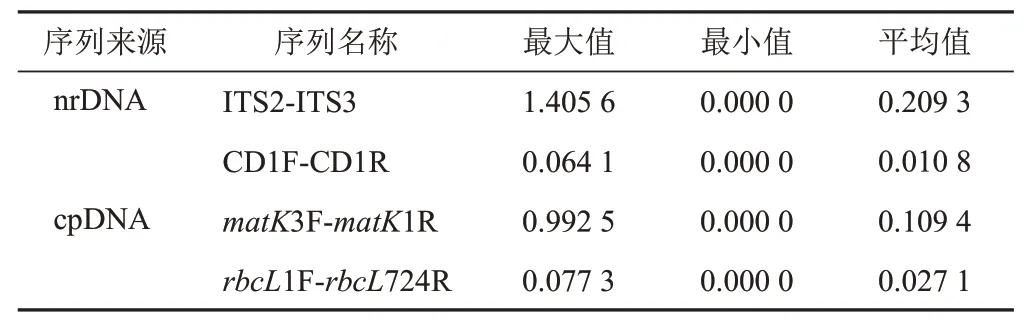
表7 肉蓯蓉不同序列種質遺傳距離
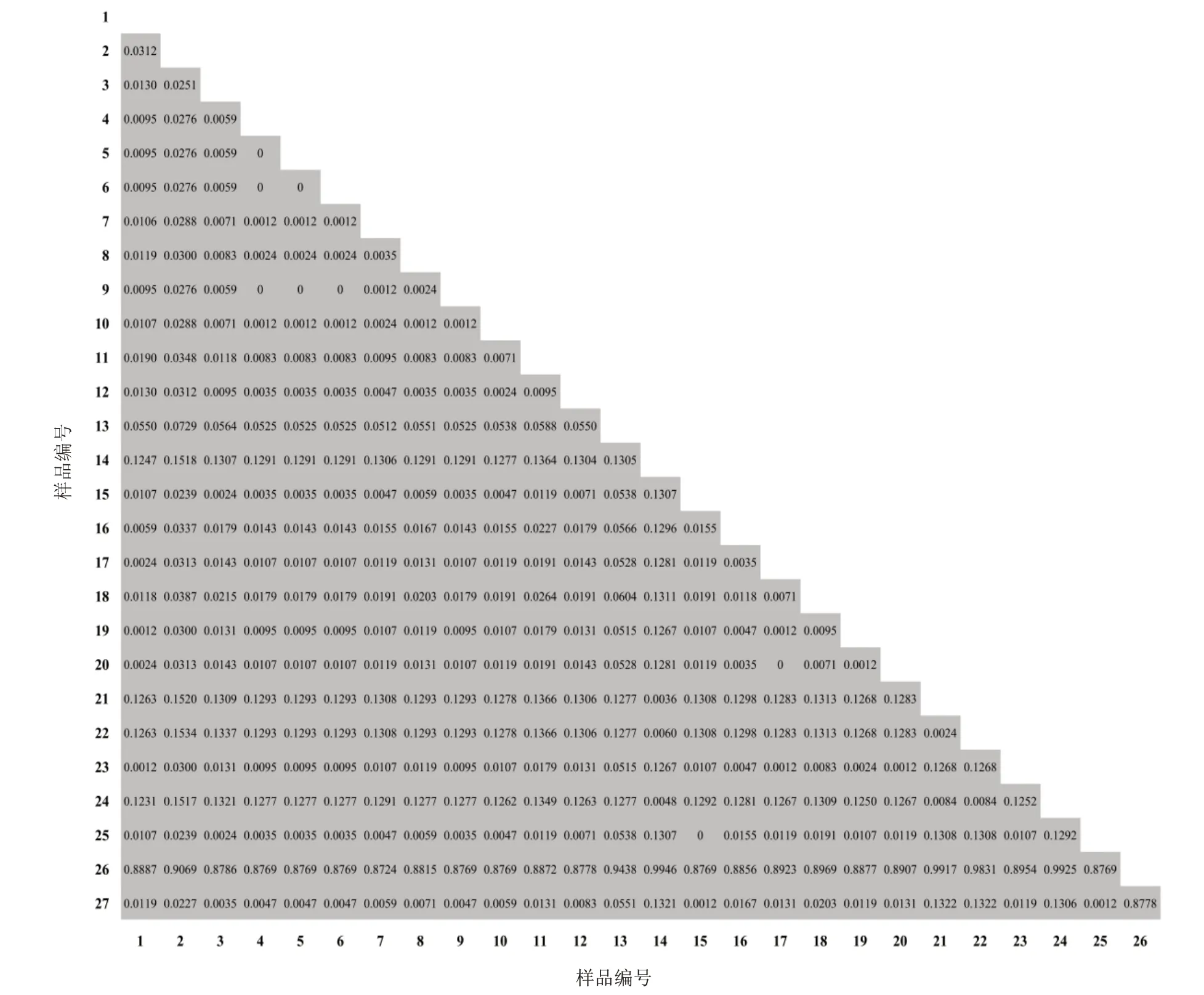
圖2 肉蓯蓉matK序列遺傳距離矩陣
2.3.6 聚類分析
通過GenBank數據庫下載草蓯蓉屬Boschniakia植物丁座草Boschniakia himalaicaHook.f.et Thoms.序列信息(ITS和CD為AY911212.1,matK為MH659839.1,rbcL為MG546905.1),以其為外類群與nrDNA和cpDNA序列及2組合并序列構建鄰接法(NJ)聚類樹[14],結果表明,外類群丁座草可單獨聚為一支,4對序列在屬水平上的鑒定成功率均為100%。ITS和CD序列聚類圖顯示,27份肉蓯蓉樣品可聚為3類,其中樣品27單獨為一個進化支,樣品6、11、12、13、14、19、21、22、26、24為一個進化支,其余為一個進化支(見圖3A、圖3B)。matK序列聚類圖顯示,27份肉蓯蓉樣品可聚為4類,其中樣品13單獨為一個進化支,樣品22、21、24、14為一個進化支,樣品19、1、23、16、20、18、17為一個進化支,其余為一個進化支(見圖3C)。rbcL序列聚類圖顯示,27份肉蓯蓉樣品可聚為4類,其中樣品26、6、2分別為一個進化支,其余為一個進化支(見圖3D)。2組合并序列聚類圖表明,nrDNA合并序列可將大多數沙蓯蓉、鹽生肉蓯蓉與正品肉蓯蓉區分開,只有樣品11、12、13未能區分(見圖3E);cpDNA合并序列則可將樣品13單獨聚為一個進化支,其余肉蓯蓉聚為另一個進化支,從而進行有效鑒別區分(見圖3F)。所有構建的NJ聚類樹均顯示樣品1、23、21、22(鹽生肉蓯蓉)分別為不同進化支,樣品1和23為一個進化支,樣品21和22則為另一個進化支,且蘭州肉蓯蓉始終與肉蓯蓉聚為一支。
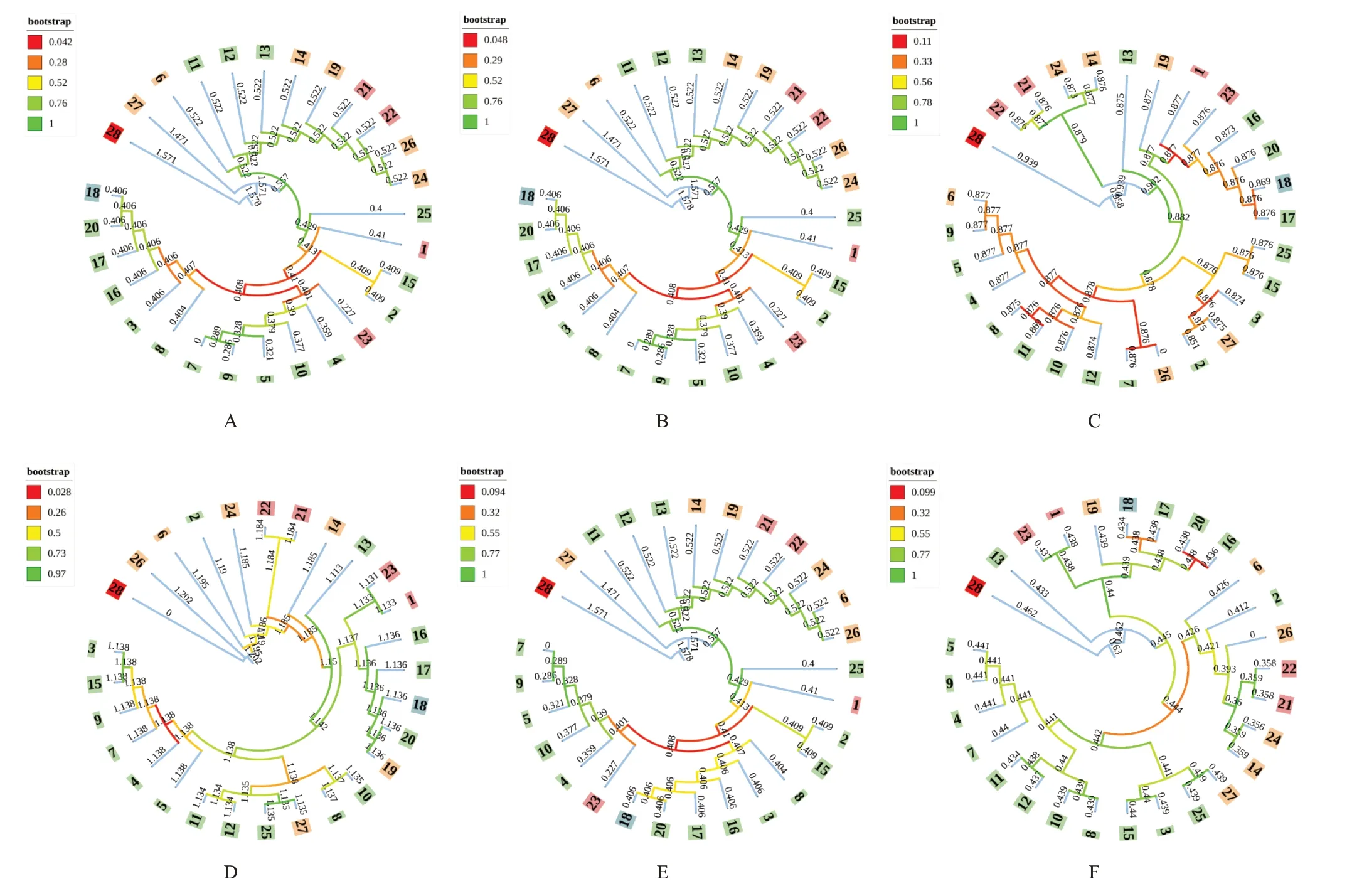
圖3 肉蓯蓉樣品基因組序列NJ系統聚類樹
2.3.7 方差分析
將采樣地不同但屬于同種的肉蓯蓉樣品歸為一組,共分為3組,即肉蓯蓉組、鹽生肉蓯蓉組和沙蓯蓉組,采用Arlequin3.5軟件[15]對nrDNA和cpDNA合并序列進行方差分析(見表8),結果表明,肉蓯蓉遺傳變異主要來自組內變異,即同一物種不同種質間分化明顯。ITS和CD的合并序列遺傳分化指數(Fst)為0.206 8,基因流(Nm)為1.01;matK和rbcL的合并序列Fst 為0.230 7,Nm為0.92,說明肉蓯蓉不同種質間存在一定基因流動,但基因交流不頻繁。

表8 肉蓯蓉樣品基因組序列方差分析
3 討論
以往研究表明,nrDNA ITS序列進化速率快,變異信息位點較豐富,被廣泛應用于植物的遺傳多樣性分析[16]。而cpDNA序列則可提供豐富的遺傳信息,為評估物種遺傳多樣性和親緣關系提供科學參考[17]。本研 究 利 用4對 引 物(ITS2-ITS3、CD1F-CD1R、matK3F-matK1R和rbcL1F-rbcL724R)進行序列比對、遺傳距離分析及NJ聚類樹的構建,結果表明,肉蓯蓉種質資源多樣性程度較高,且在種質間水平上均有豐富的變異性。畢毓芳等[18]研究表明,matK序列是進化速率最快的葉綠體基因組序列,常在科級、屬級、種間和種內水平上被用于系統進化研究。馬麗等[19]在對石榴種質資源進行基因組序列分析時發現石榴物種的matK基因片段擴增及測序成功率高,聚類準確度較強。董斌等[20]研究發現,在matK序列變異位點較多的情況下,遺傳差異也較大,有利于鑒別同一物種的不同種質。本研究中,matK序列是4對序列中變異位點最多的序列,具有481個變異位點,其中單一突變位點為369個,所占比例最大,是變異的主要來源。按Grant等[21]提出的標準,單倍型多樣性以0.5為臨界值,核苷酸多樣性以0.005為臨界值,二者值越大,群體的多樣性程度越高。4對序列中matK序列單倍型多樣性最高,而cpDNA合并序列的單倍型多樣性明顯高于nrDNA合并序列,因此,matK序列更適合作為肉蓯蓉遺傳多樣性分析的序列,也證明了葉綠體基因片段在群體遺傳學研究中的優越性。此外,對鹽生肉蓯蓉,由于采集樣品地理位置的差異,發現不同地區的同種肉蓯蓉也存在地理遺傳差異性,與楊俊譽等[22]研究結果一致。
分子鑒定技術可有效對中藥材進行準確鑒別,是對傳統經典鑒定方法的有效補充,彌補了經典鑒定方法的不足[23]。近年來,許多學者為實現對物種進行快速鑒定,對適合植物物種鑒定的基因序列進行了深入研究,其中主要包括ITS、ITS2、psbA-trnH、matK和rbcL序列[24]。徐素素等[25]認為,單獨類型的序列鑒別存在一定局限性,分子鑒定體系的完善需要多條序列聯合應用。李波等[26]在研究中使用核基因片段與葉綠體片段相結合的方法,從而達到提高物種鑒別效率。本研究以nrDNA和cpDNA序列構建NJ聚類樹,發現單獨某對序列并不能在種水平上有較好的鑒別效率。ITS和CD序列聚類圖表明,多數肉蓯蓉與沙蓯蓉、鹽生肉蓯蓉親緣關系較遠,會分別聚于不同進化支,但也存在小部分肉蓯蓉與其他物種聚為一支的情況;matK和rbcL序列聚類圖表明,沙蓯蓉與鹽生肉蓯蓉親緣關系較近,可聚為同一大支,而兩物種又會分別聚為不同小支,但部分肉蓯蓉和鹽生肉蓯蓉存在聚類混亂的情況,不能準確鑒別。故將nrDNA和cpDNA序列聯合使用,可進一步實現對肉蓯蓉種質的快速、準確鑒別。
蘭州肉蓯蓉在《中國植物志》中作為單獨的物種具有一定植物學分類地位,但近年來也有學者建議將蘭州肉蓯蓉及沙蓯蓉歸為同個物種[27]。而陳博等[6]在對甘肅肉蓯蓉進行指標性成分含量測定研究發現,蘭州肉蓯蓉與沙蓯蓉成分含量存在一定差異,與肉蓯蓉成分含量較為相似。本研究中,通過計算遺傳距離及構建NJ聚類樹,結果蘭州肉蓯蓉與肉蓯蓉的遺傳距離較近,均可聚為同一個進化支。由此,建議將蘭州肉蓯蓉與其他肉蓯蓉進行分類區別,考慮作為正品肉蓯蓉的藥源補充,提高其資源開發利用度,后續可就此問題進行深入研究。
近年來,隨著對肉蓯蓉藥材需求量的提升,為保證臨床安全用藥,對肉蓯蓉種質資源的鑒別、評價及保護至關重要。本研究基于nrDNA和cpDNA序列,對甘肅省4種肉蓯蓉進行分子鑒定,結果表明肉蓯蓉種質資源多樣性程度較高,可為進一步細化肉蓯蓉不同種質資源的分類提供參考。
