FAM3C介導(dǎo)TGF-β1誘導(dǎo)的人血管平滑肌細(xì)胞表型轉(zhuǎn)換、活力增強(qiáng)及遷移*
2023-01-05 03:01:42訾亞飛王正力訾亞婉李雅馨劉楊東
中國病理生理雜志 2022年12期
關(guān)鍵詞:研究
訾亞飛,王正力,訾亞婉,李雅馨,劉楊東,趙 渝△
(1重慶醫(yī)科大學(xué)附屬第一醫(yī)院血管外科,重慶 400016;2重慶醫(yī)科大學(xué)附屬第一醫(yī)院呼吸及重癥醫(yī)學(xué)科,重慶 400016;3深圳大學(xué)附屬華南醫(yī)院血管外科,廣東深圳 518100)
內(nèi)膜增生(intimal hyperplasia)是一種特別的血管重塑狀態(tài),發(fā)生于動(dòng)靜脈內(nèi)瘺、動(dòng)脈粥樣硬化及血管再狹窄病變[1-2]。而引起內(nèi)膜增生的主要原因是由于血管平滑肌細(xì)胞(vascular smooth muscle cells,VSMCs)在多種信號分子[如轉(zhuǎn)化生長因子β1(transforming growth factor-β1,TGF-β1)]刺激下,由收縮表型[標(biāo)志物為α-平滑肌肌動(dòng)蛋白(α-smooth muscle actin,α-SMA)]轉(zhuǎn)化為合成表型[標(biāo)志物為骨橋蛋白(osteopontin,OPN)及波形蛋白(vimentin)等],且自身發(fā)生肥大及活力增強(qiáng),并由中膜遷移至內(nèi)膜引起血管內(nèi)膜增生[1-3]。雖然目前對VSMCs表型轉(zhuǎn)換有較多研究,但其有效治療靶點(diǎn)仍不清楚。
序列相似家族3成員C(family with sequence similarity 3 member C,F(xiàn)AM3C)又稱白細(xì)胞介素樣上皮-間充質(zhì)轉(zhuǎn)化(epithelial-mesenchymal transition,EMT)誘導(dǎo)因子(interleukin-like EMT inducer,ILEI)[4],是FAM3家族的成員,也是重要的細(xì)胞因子樣蛋白。研究表明,F(xiàn)AM3C在組織中普遍表達(dá)[5],由細(xì)胞內(nèi)分泌至細(xì)胞外,并且在多種癌細(xì)胞中高表達(dá),促進(jìn)癌癥進(jìn)展和轉(zhuǎn)移,而循環(huán)中的FAM3C是自噬和癌癥的生物標(biāo)志物[6-8]。另外,F(xiàn)AM3C作為TGF-β1的靶基因,其蛋白參與TGF-β1在腎小管上皮細(xì)胞及癌癥中誘導(dǎo)的EMT,其下調(diào)可抑制TGF-β1誘導(dǎo)的改變[9-10]。還有研究提示,F(xiàn)AM3C通過降低糖異生和脂肪相關(guān)基因的mRNA和蛋白水平而抑制糖異生,從而減輕肥胖小鼠的胰島素抵抗、高血糖及脂肪變性[11]。不僅如此,F(xiàn)AM3C還參與成骨細(xì)胞的分化[12]。然而,F(xiàn)AM3C是否參與TGF-β1誘導(dǎo)的VSMCs表型轉(zhuǎn)換尚未見報(bào)道。
蛋白激酶B(protein kinase B,PKB/Akt)是一種絲氨酸/蘇氨酸蛋白激酶,可被許多生長因子和細(xì)胞因子以磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)依賴的方式激活[13]。Akt是一種重要的細(xì)胞內(nèi)分子,其激活對細(xì)胞存活和凋亡有重要影響,并在心血管疾病中調(diào)節(jié)多種細(xì)胞功能[14]。有研究報(bào)道Akt活化在TGF-β1促進(jìn)VSMCs表型轉(zhuǎn)換及增殖的過程中起著重要作用[15-16],而抑制Akt的激活可減輕血管重構(gòu)[17]。
本研究旨在探討FAM3C在TGF-β1誘導(dǎo)VSMCs表型轉(zhuǎn)換、活力增強(qiáng)及遷移中的作用及其潛在機(jī)制,以期獲得干預(yù)內(nèi)膜增生的相關(guān)靶點(diǎn)。
材料和方法
1 材料與試劑
原代人主動(dòng)脈平滑肌細(xì)胞(HTX3138)購自深圳豪地華拓生物科技有限公司;DMEM/F12培養(yǎng)液購自Gibco;胎牛血清購自Biosharp;青霉素和鏈霉素(C0222)購自碧云天;TGF-β1(100-21-2)購自杭州聯(lián)科生物技術(shù)股份有限公司;CCK-8(AR1160)購自武漢博士德生物工程有限公司;Akt inhibitor VIII(Akti-VIII;T3346)購自TargetMol;FAM3C小干擾RNA(si-FAM3C)和陰性對照小干擾RNA(si-NC)均由廣州銳博生物技術(shù)有限公司合成;jetPRIME transfection reagent(101000046)購自Polyplus;FAM3C過表達(dá)慢病毒(LV-FAM3C)及空載病毒(LV-GFP)由賽業(yè)(蘇州)生物科技有限公司合成;Transwell小室購自Corning;增強(qiáng)化學(xué)發(fā)光(enhanced chemiluminescence,ECL)顯影試劑盒(4AW011-500)購自北京四正柏生物科技有限公司;OPN兔抗(22952-1-AP)和辣根過氧化物酶標(biāo)記的山羊抗鼠IgG(SA00001-1)購自Proteintech;vimentin鼠抗(BF8006)、FAM3C兔抗(DF12605)、α-SMA兔抗(AF1032)、Akt兔抗(AF0836)、磷酸化Akt(phosphorylated Akt,p-Akt)兔抗(AF0016)及β-actin兔抗(AF7018)購自Affinity;增殖細(xì)胞核抗原(proliferating cell nuclear antigen,PCNA)抗體(D3H8P)購自Cell Signaling Technology;辣根過氧化物酶標(biāo)記的山羊抗兔IgG(A21020)、熒光標(biāo)記的山羊抗兔IgG(A23420)及熒光標(biāo)記的山羊抗鼠IgG(A23210)購自Abbkine。
2 主要方法
2.1 細(xì)胞培養(yǎng)及處理原代人VSMCs用DMEM/F12培養(yǎng)液加10%胎牛血清和1%青霉素-鏈霉素在37℃及5%CO2環(huán)境下培養(yǎng)。首先用不同濃度(0、1、2、5及10 μg/L)的TGF-β1作用24 h和10 μg/L的TGF-β1作用不同時(shí)間(0、6、12、24及48 h),以確定最佳作用濃度和時(shí)間,并以10 μg/L的TGF-β1作用不同時(shí)間(0、6、12及24 h)來誘導(dǎo)VSMCs表型轉(zhuǎn)換。在驗(yàn)證敲減FAM3C效率的實(shí)驗(yàn)中,將細(xì)胞分為control(CTL)組、si-NC組和si-FAM3C組,之后再分為si-NC組、si-NC+TGF-β1組、si-FAM3C組和si-FAM3C+TGF-β1組;在熒光顯微鏡下驗(yàn)證過表達(dá)慢病毒是否轉(zhuǎn)染成功的實(shí)驗(yàn)中,將細(xì)胞分為空白(blank)組、LVGFP組和LV-FAM3C組,之后再分為LV-GFP組、LVGFP+TGF-β1組、LV-FAM3C組和LV-FAM3C+TGF-β1組;在Akt抑制劑處理的實(shí)驗(yàn)中,將細(xì)胞分為LV-GFP組、LV-GFP+TGF-β1組、LV-FAM3C組、LV-FAM3C+TGF-β1組、LV-FAM3C+Akti-VIII組 和LV-FAM3C+TGF-β1+Akti-VIII組,Akti-VIII(5 μmol/L)提前2 h加入處理組。各組處理前均饑餓細(xì)胞24 h。
2.2 細(xì)胞活力的測定以每孔1×104細(xì)胞在96孔板種板孵育。細(xì)胞貼壁后根據(jù)分組進(jìn)行處理,同時(shí)設(shè)置空白對照組及5個(gè)復(fù)孔。次日按照CCK-8試劑盒說明書,在各孔加入10 μL CCK-8溶液,37℃避光孵育2 h,酶標(biāo)儀在450 nm處測量各孔吸光度。
2.3 細(xì)胞轉(zhuǎn)染細(xì)胞傳代至6孔板中,融合70%~80%時(shí),按照轉(zhuǎn)染說明書將2 μL si-FAM3C(20 μmol/L)或2 μL si-NC(20 μmol/L)與2 μL jetPRIME transfection reagent加 入 到200 μL jetPRIME buffer中,充分混合15 min后,加入到各孔中,加入2 mL有血清培養(yǎng)液繼續(xù)培養(yǎng),轉(zhuǎn)染48 h后,Western blot檢測轉(zhuǎn)染效率,收集細(xì)胞進(jìn)行后續(xù)實(shí)驗(yàn)。si-FAM3C的序列 為5′-CTACAAAGCCTCCCAGATA-3′,si-NC的 序列為5′-GGCUCUAGAAAAGCCUAUGC-3′。
2.4 慢病毒轉(zhuǎn)染將狀態(tài)良好的對數(shù)期VSMCs種于6孔板中,次日將LV-GFP組和LV-FAM3C組細(xì)胞根據(jù)賽業(yè)公司提供的說明書按感染復(fù)數(shù)(multiplicity of infection,MOI)=50進(jìn)行慢病毒感染實(shí)驗(yàn)。轉(zhuǎn)染10 h后,更換新鮮培養(yǎng)液,培養(yǎng)48 h后在熒光顯微鏡下拍照驗(yàn)證,收集細(xì)胞進(jìn)行后續(xù)的實(shí)驗(yàn)。
2.5 Transwell實(shí)驗(yàn)將3×105細(xì)胞用無血清培養(yǎng)液懸浮,取200 μL加入到上室,700 μL含20%胎牛血清的培養(yǎng)液加入下室,并在37℃培養(yǎng)箱中孵育24 h,次日用甲醛固定15 min,結(jié)晶紫染色,棉簽擦拭上室殘留細(xì)胞,顯微鏡下拍照并計(jì)數(shù)。
2.6 免疫熒光染色將細(xì)胞種于24孔板中蓋玻片上。經(jīng)處理后,4%多聚甲醛固定細(xì)胞15 min,PBS洗滌3次,0.3%Triton X-100破膜10 min,正常山羊血清封閉1 h,加入vimentin和α-SMA抗體(1∶150稀釋),4℃過夜孵育。次日用熒光標(biāo)記的山羊抗兔及抗鼠IgG(1∶200稀釋)孵育1 h,經(jīng)4′,6-二脒基-2-苯基吲哚(4′,6-diamidino-2-phenylindole,DAPI)染核后,抗熒光淬滅劑封片,避光條件下熒光顯微鏡獲取圖像。
2.7 Western blot實(shí)驗(yàn)細(xì)胞處理后,用含磷酸酶及蛋白酶抑制劑的RIPA裂解液在冰上提取蛋白,BCA蛋白檢測試劑測定蛋白濃度。加入上樣緩沖液并加熱變性,SDS-PAGE分離蛋白,轉(zhuǎn)移到聚二氟乙烯膜。5%脫脂奶粉室溫下封閉2 h,OPN、vimentin、β-actin、PCNA、α-SMA、FAM3C、Akt及p-Akt抗體(1∶1 000稀釋)4℃孵育過夜。次日用TBST洗滌3次,辣根過氧化物酶標(biāo)記的山羊抗兔IgG(1∶10 000稀釋)和山羊抗鼠IgG(1∶5 000稀釋)室溫孵育2 h,TBST洗滌3次。ECL發(fā)光液顯影,ImageJ軟件分析圖像。
3 統(tǒng)計(jì)學(xué)處理
所有統(tǒng)計(jì)分析及制圖均使用GraphPad Prism 8進(jìn)行。數(shù)據(jù)以均數(shù)±標(biāo)準(zhǔn)差(mean±SD)表示。采用單因素方差分析檢驗(yàn)多組間差異,后用Turkey法進(jìn)行兩兩比較。以P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
結(jié) 果
1 TGF-β1對VSMCs中FAM3C表達(dá)的影響
TGF-β1在作用24 h時(shí)隨著濃度(0、1、2、5和10 μg/L)增加上調(diào)VSMCs中FAM3C的表達(dá),TGF-β1(10 μg/L)隨作用時(shí)間(0、6、12、24及48 h)增加上調(diào)FAM3C表達(dá),以10 μg/L及24 h時(shí)最為顯著(P<0.05),故作為后續(xù)實(shí)驗(yàn)條件,見圖1。此結(jié)果亦提示FAM3C是TGF-β1的靶基因。
2 TGF-β1誘導(dǎo)VSMCs表型轉(zhuǎn)換
10 μg/L TGF-β1呈時(shí)間依賴性(0、6、12和24 h)上調(diào)OPN及vimentin表達(dá),下調(diào)α-SMA表達(dá),0、6、12及24 h組間均有顯著差異(P<0.05),vimentin免疫熒光與Western blot結(jié)果趨勢類似,見圖2。
3 敲減FAM3C抑制TGF-β1誘導(dǎo)的VSMCs表型轉(zhuǎn)換、活力增強(qiáng)及遷移
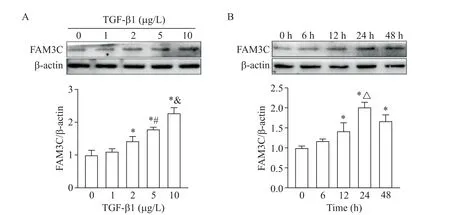
Figure 1.Effects of TGF-β1 on the expression of FAM3C in VSMCs.Primary human VSMCs were treated with 0,1,2,5 and 10 μg/L of TGF-β1 for 24 h(A)or 10 μg/L TGF-β1 for 0,6,12,24 and 48 h(B).Mean±SD.n=3.*P<0.05 vs 0 μg/L or 0 h;#P<0.05 vs 2 μg/L;&P<0.05 vs 5 μg/L;△P<0.05 vs 12 h.圖1 TGF-β1對VSMCs中FAM3C表達(dá)的影響
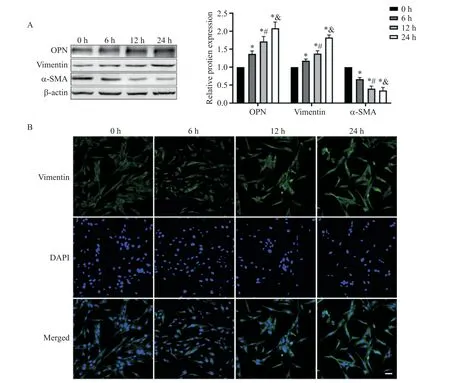
Figure 2.TGF-β1 induced phenotypic switch of VSMCs.A:the protein expression of OPN,vimentin and α-SMA detected by Western blot;B:immunofluorescence staining for vimentin(scale bar=50 μm).Mean±SD.n=3.*P<0.05 vs 0 h;#P<0.05 vs 6 h;&P<0.05 vs 12 h.圖2 TGF-β1誘導(dǎo)VSMCs表型轉(zhuǎn)換
與si-NC和CTL組相比,si-FAM3C組中FAM3C表達(dá)降低(P<0.05),提示已敲減FAM3C(圖3A)。與si-NC組相比,si-NC+TGF-β1組細(xì)胞活力增強(qiáng)(P<0.05),OPN、vimentin和PCNA表達(dá)增加(P<0.05),α-SMA表達(dá)降低(P<0.05),細(xì)胞遷移數(shù)量增加(P<0.05);與si-NC+TGF-β1組相比,si-FAM3C+TGF-β1組中TGF-β1對VSMCs的上述作用被削弱;此外,與si-NC組相比,si-FAM3C組細(xì)胞活力減弱(P<0.05),OPN、vimentin和PCNA表達(dá)減少(P<0.05),α-SMA表達(dá)增加(P<0.05),細(xì)胞遷移數(shù)量減少(P<0.05),見圖3B~D。上述結(jié)果提示敲減FAM3C抑制VSMCs表型轉(zhuǎn)換、活力及遷移。
4 過表達(dá)FAM3C促進(jìn)TGF-β1誘導(dǎo)的VSMCs表型轉(zhuǎn)換、活力增強(qiáng)及遷移
與blank組 相比,LV-GFP和LV-FAM3C組熒 光強(qiáng)度增加,提示慢病毒轉(zhuǎn)染成功(圖4A)。與LVGFP組相比,LV-FAM3C組細(xì)胞活力增強(qiáng)(P<0.05),OPN、vimentin和PCNA表達(dá)增加(P<0.05),α-SMA表達(dá)減少(P<0.05),細(xì)胞遷移數(shù)量增多(P<0.05),vimentin和α-SMA免疫熒光結(jié)果與Western blot結(jié)果趨勢類似;與LV-GFP+TGF-β1組相比,LV-FAM3C+TGF-β1組趨勢與LV-FAM3C組類似,提示過表達(dá)FAM3C增強(qiáng)TGF-β1對VSMCs表型轉(zhuǎn)換、活力及遷移的作用,見圖4B~D及圖5。
5 FAM3C通過Akt介導(dǎo)TGF-β1誘導(dǎo)的VSMCs改變
與si-NC組和LV-GFP組相比,TGF-β1提高p-Akt蛋白水平(P<0.05);敲減FAM3C可降低p-Akt蛋白水平及減弱TGF-β1的誘導(dǎo)作用(P<0.05);過表達(dá)FAM3C對p-Akt蛋白水平的作用與敲減FAM3C相反;而加入Akti-VIII后,p-Akt蛋白水平降低(P<0.05),OPN和vimentin表達(dá)減少(P<0.05),α-SMA表達(dá)增加(P<0.05),見圖6。
討 論
正常VSMCs以收縮表型存在,有利于對抗血管張力,維持血管壁穩(wěn)態(tài)。然而,在多種病理狀態(tài)下,VSMCs的合成表型轉(zhuǎn)化增加,導(dǎo)致增殖、遷移和細(xì)胞外基質(zhì)合成,從而引起內(nèi)膜增生,是引起動(dòng)脈粥樣硬化、血管再狹窄和高血壓的重要原因之一[14,18-19],許多細(xì)胞因子參與VSMCs表型轉(zhuǎn)換的調(diào)節(jié),其中TGFβ1通過激活其下游Smad2/3和PI3K/Akt等通路來發(fā)揮作用[2,16]。此外,TGF-β1還可通過促進(jìn)縫隙連接蛋白(connexin)43表達(dá)而導(dǎo)致VSMCs的增殖活力增加[20]。在本研究中,TGF-β1呈時(shí)間依賴性調(diào)節(jié)VSMCs表型轉(zhuǎn)換,并可增強(qiáng)細(xì)胞活力及遷移能力,與既往研究結(jié)果一致[2],因此對TGF-β1下游靶基因的調(diào)節(jié)是干預(yù)內(nèi)膜增生的關(guān)鍵。
FAM3C作為FAM3家族重要成員之一,較多研究涉及細(xì)胞EMT及遷移。FAM3C通過Akt及細(xì)胞外信號調(diào)節(jié)激酶通路參與TGF-β1誘導(dǎo)的腎小管上皮EMT,促進(jìn)腎纖維化[10],還可通過PI3K/Akt途徑參與胃癌的EMT及細(xì)胞遷移過程[8],但是否參與血管內(nèi)膜增生還未見報(bào)道。本研究結(jié)果表明,在VSMCs中,TGF-β1可使FAM3C蛋白表達(dá)增加,驗(yàn)證了FAM3C是TGF-β1的靶基因。為進(jìn)一步研究FAM3C在VSMCs表型轉(zhuǎn)換中的作用,我們分別對FAM3C行敲減及過表達(dá)處理。敲減FAM3C可提高收縮表型標(biāo)志物α-SMA表達(dá),降低TGF-β1誘導(dǎo)的合成表型標(biāo)志物OPN和vimentin表達(dá),同時(shí)降低VSMCs活力及遷移能力,而過表達(dá)FAM3C可促進(jìn)TGF-β1對VSMCs表型轉(zhuǎn)換、活力及遷移的調(diào)節(jié),因此我們證實(shí)FAM3C介導(dǎo)TGFβ1誘導(dǎo)的VSMCs表型轉(zhuǎn)換、活力增強(qiáng)及遷移。FAM3C的作用可能與其形態(tài)變化有關(guān),研究表明二聚體FAM3C較單聚體可使癌細(xì)胞侵襲增加[21],但在VSMCs中是否發(fā)生形態(tài)改變還需進(jìn)一步研究。此外,有研究指出FAM3C上調(diào)陰陽1(yin yang 1,YY1)表達(dá),最終激活A(yù)kt-cyclin D1通路促進(jìn)乳腺癌細(xì)胞增殖和遷移[7];而在頸動(dòng)脈血管損傷后上調(diào)YY1,可抑制VSMCs收縮型標(biāo)志物的表達(dá),從而導(dǎo)致內(nèi)膜增生[22]。在本實(shí)驗(yàn)中,過表達(dá)及敲減FAM3C均可獨(dú)立于TGFβ1調(diào)節(jié)VSMCs表型轉(zhuǎn)換、活力及遷移,因此FAM3C可能通過調(diào)節(jié)YY1或Akt參與對VSMCs的調(diào)節(jié)。
Akt作為重要的細(xì)胞內(nèi)因子,參與調(diào)控細(xì)胞增殖、分化及凋亡等細(xì)胞功能。研究表明Akt通過抑制VSMCs凋亡而導(dǎo)致內(nèi)膜增厚[16,23],此外,Akt通過激活哺乳動(dòng)物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)而介導(dǎo)攝食抑制因子(nesfatin-1)誘導(dǎo)的VSMCs表型轉(zhuǎn)換、周期轉(zhuǎn)換及增殖[24]。另有研究表明,芙蓉葉通過抑制Akt/激活蛋白1(activator protein-1,AP-1)通路,抑制腫瘤壞死因子α(tumor necrosis factor-α,TNF-α)誘導(dǎo)的VSMCs遷移[25]。因此,Akt在內(nèi)膜增生發(fā)生發(fā)展中發(fā)揮重要作用。在本研究中,TGF-β1使Akt活化,且Akt活化在FAM3C敲減后被阻斷,提示Akt是FAM3C的下游蛋白。有研究提示,過表達(dá)FAM3C可激活熱休克因子1(heat shock factor 1,HSF1)-鈣調(diào)素(calmodulin,CaM)-Akt通路,并增加Akt磷酸化,從而抑制糖異生[26],另外,血管緊張素II(angiotensin II,Ang II)通過增加Ca2+的攝入而誘導(dǎo)早期生長反應(yīng)因子1(early growth response protein-1,Egr-1)表達(dá),而導(dǎo)致內(nèi)膜增生,敲減CaM抑制Ang II對VSMCs表型的改變[27]。更重要的是,作為CaM的下游激酶,鈣/CaM依賴性蛋白激酶II(Ca2+/CaM-dependent protein kinase II,CaMKII)通過調(diào)控線粒體對Ca2+的攝取及VSMCs遷移,導(dǎo)致血管損傷后的新生內(nèi)膜形成[28]。在本研究中,過表達(dá)FAM3C激活A(yù)kt,促進(jìn)TGF-β1對Akt的活化,其機(jī)制可能與CaM的表達(dá)有關(guān),而Akt抑制劑作用后阻斷了FAM3C對VSMCs表型轉(zhuǎn)換的作用,因此FAM3C可能通過調(diào)節(jié)Akt參與TGF-β1誘導(dǎo)的VSMCs表型轉(zhuǎn)換。這也可解釋敲減及過表達(dá)FAM3C獨(dú)立調(diào)節(jié)VSMCs表型轉(zhuǎn)換的作用與Akt的激活有關(guān),所以抑制Akt對VSMCs表型轉(zhuǎn)換調(diào)節(jié)起到重要作用。
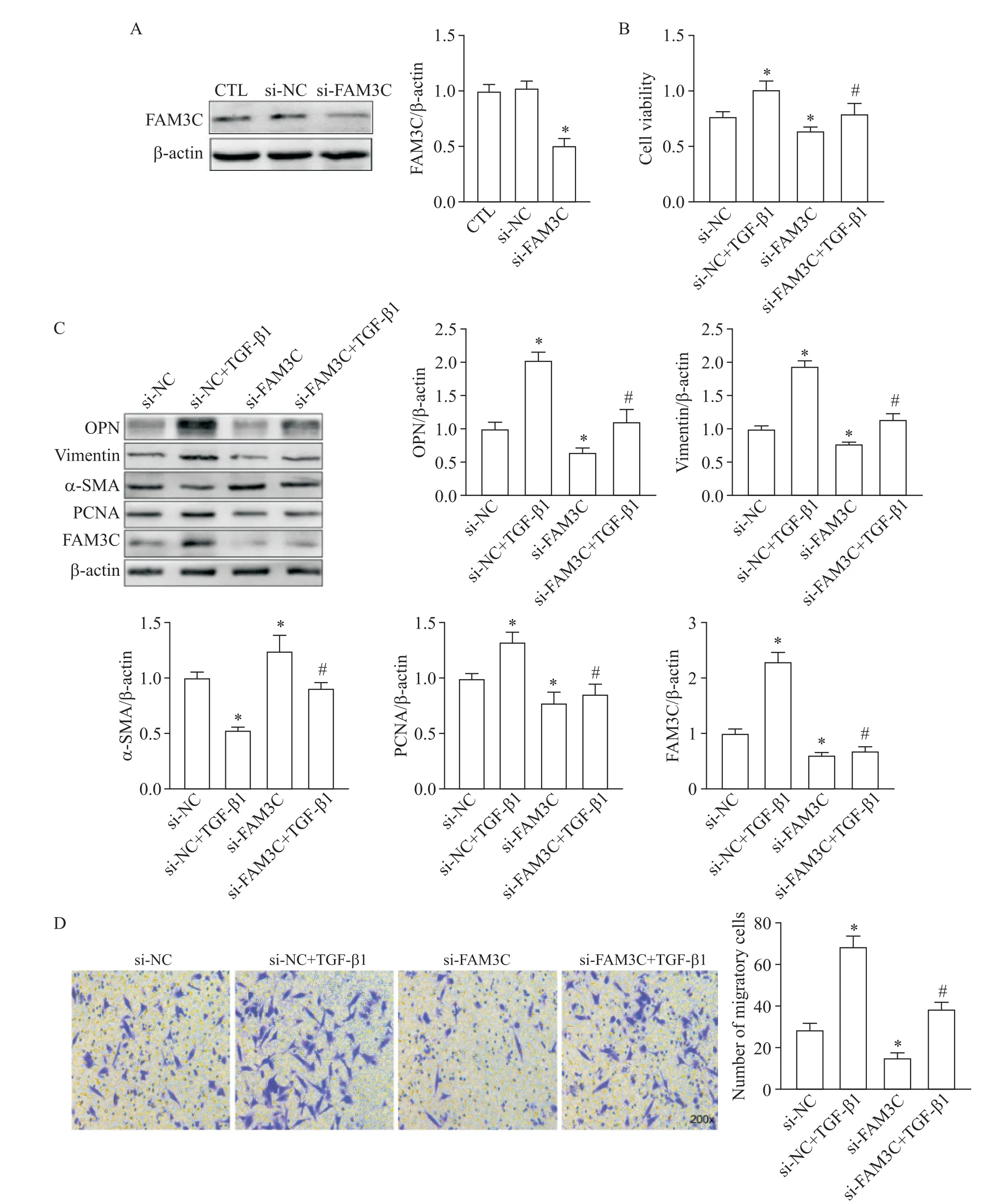
Figure 3.Knockdown of FAM3C inhibited TGF-β1-induced phenotypic switch and attenuated viability and migration of VSMCs.A:verification of FAM3C knockdown by Western blot;B:the cell viability detected by CCK-8 assay;C:the expression of phenotypic markers,PCNA and FAM3C detected by Western blot;D:the cell migration detected by Transwell assay(×200).Mean±SD.n=3.*P<0.05 vs CTL or si-NC group;#P<0.05 vs si-NC+TGF-β1 group.圖3敲減FAM3C抑制TGF-β1誘導(dǎo)的VSMCs表型轉(zhuǎn)換并減弱細(xì)胞活力和遷移能力
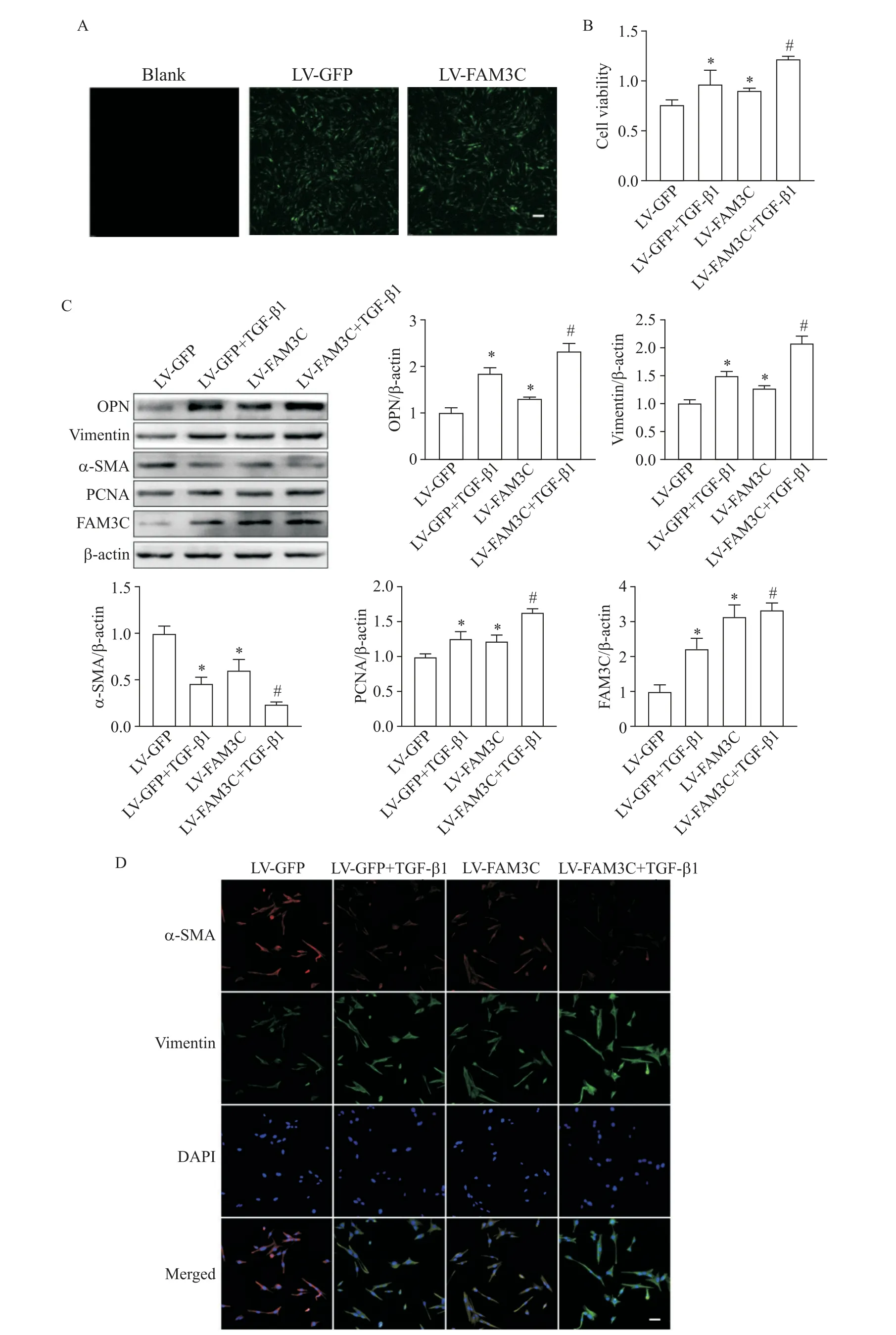
Figure 4.Overexpression of FAM3C promoted TGF-β1-induced phenotypic switch and enhanced viability of VSMCs.A:verification of lentivirus(LV)infection(scale bar=100 μm);B:the cell viability detected by CCK-8 assay;C:the expression of phenotypic markers,PCNA and FAM3C detected by Western blot;D:immunofluorescence staining for α-SMA and vimentin(scale bar=50 μm).Mean±SD.n=3.*P<0.05 vs LV-GFP group;#P<0.05 vs LV-GFP+TGF-β1 group.圖4過表達(dá)FAM3C促進(jìn)TGF-β1誘導(dǎo)的VSMCs表型轉(zhuǎn)換并增強(qiáng)細(xì)胞活力
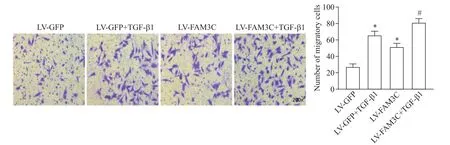
Figure 5.Overexpression of FAM3C promoted TGF-β1-induced migation of VSMCs.The cell migration was detected by Transwell assay(×200).Mean±SD.n=3.*P<0.05 vs LV-GFP group;#P<0.05 vs LV-GFP+TGF-β1 group.圖5過表達(dá)FAM3C促進(jìn)TGF-β1誘導(dǎo)的VSMCs遷移
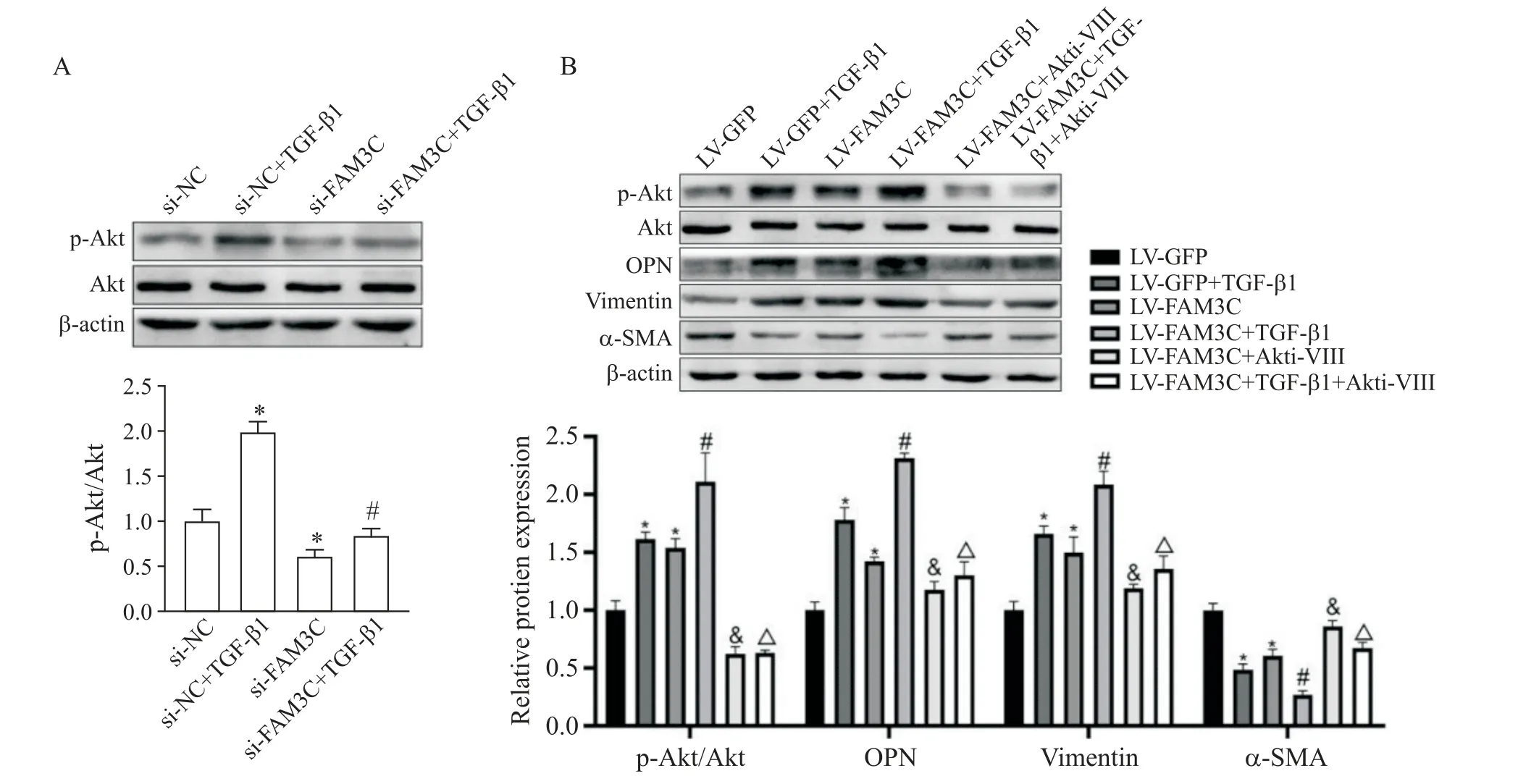
Figure 6.FAM3C mediated TGF-β1-induced phenotypic changes in VSMCs through Akt.A:effects of TGF-β1 and/or FAM3C knockdown on Akt phosphorylation;B:effects of TGF-β1 and/or FAM3C overexpression combined with Akt inhibitor VIII(Akti-VIII)on Akt phosphorylation and phenotypic marker expression.Mean±SD.n=3.*P<0.05 vs LV-GFP group;#P<0.05 vs LV-GFP+TGF-β1 group;&P<0.05 vs LV-FAM3C group;△P<0.05 vs LV-FAM3C+TGF-β1 group.圖6 FAM3C通過Akt介導(dǎo)TGF-β1誘導(dǎo)的VSMCs表型改變
總之,本研究發(fā)現(xiàn)FAM3C通過激活A(yù)kt參與并介導(dǎo)TGF-β1誘導(dǎo)的VSMCs表型轉(zhuǎn)換、活力增強(qiáng)及遷移,因此FAM3C可能是干預(yù)內(nèi)膜增生的潛在靶點(diǎn)。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國際商務(wù)財(cái)會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
