MLK3基因敲除抑制骨骼肌損傷后再生*
2023-01-05 03:01:58高詩(shī)娟張燕紅方光明
中國(guó)病理生理雜志 2022年12期
關(guān)鍵詞:小鼠
高詩(shī)娟,張燕紅,方光明,杜 杰
(首都醫(yī)科大學(xué)附屬北京安貞醫(yī)院,北京市心肺血管疾病研究所,北京 100029)
人體約80%的蛋白由骨骼肌儲(chǔ)存。骨骼肌萎縮不僅直接導(dǎo)致骨骼肌功能下降,還會(huì)導(dǎo)致全身營(yíng)養(yǎng)不良,嚴(yán)重影響生活質(zhì)量。骨骼肌在正常及病理狀況下處于損傷和再生的動(dòng)態(tài)過(guò)程。損傷和再生失衡會(huì)導(dǎo)致骨骼肌萎縮。骨骼肌再生由一群位于肌纖維基底膜和肌膜之間的肌衛(wèi)星細(xì)胞來(lái)完成。這群長(zhǎng)期處于相對(duì)靜息狀態(tài)的細(xì)胞在骨骼肌受到損傷刺激時(shí)被激活,繼而經(jīng)過(guò)增殖、分化、融合形成新的肌纖維,從而完成骨骼肌再生過(guò)程[1-2]。肌衛(wèi)星細(xì)胞的增殖和分化受多個(gè)信號(hào)通路調(diào)控。Notch信號(hào)通路的活化促進(jìn)肌衛(wèi)星細(xì)胞自我更新,抑制分化[3]。絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信號(hào)通路的激活則促進(jìn)肌衛(wèi)星細(xì)胞增殖和分化[4-5]。闡明肌衛(wèi)星細(xì)胞增殖和分化的調(diào)控機(jī)制,對(duì)于認(rèn)識(shí)如何維持骨骼肌穩(wěn)態(tài),阻止骨骼肌萎縮具有重要意義。
混合譜系激酶3(mixed lineage kinase 3,MLK3)是MAPK級(jí)聯(lián)反應(yīng)中重要的絲氨酸/蘇氨酸蛋白激酶,廣泛表達(dá)于骨骼肌、肺、肝及心臟等組織。MLK3通過(guò)激活MAPK下游成員c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)、細(xì)胞外信號(hào)調(diào)節(jié)激酶(extracellular signal-regulated kinase,ERK)及p38 MAPK等,調(diào)控細(xì)胞增殖、分化和凋亡[6]。MLK3基因敲除(MLK3gene knockout,MLK3KO)小鼠心臟功能下降,心臟纖維化加重[7-8]。但MLK3在骨骼肌損傷再生中的作用未見(jiàn)報(bào)道。本研究在MLK3KO小鼠及野生型(wild-type,WT)小鼠中建立骨骼肌損傷再生模型,探討MLK3在骨骼肌再生中的作用及其機(jī)制。
材料和方法
1 動(dòng)物
10~12周齡SPF級(jí)雄性C57BL6/J小鼠購(gòu)自北京唯尚立德公司,以C57BL6/J為背景的MLK3KO小鼠由上海邦耀公司協(xié)助構(gòu)建,均飼養(yǎng)于首都醫(yī)科大學(xué)附屬北京安貞醫(yī)院SPF級(jí)動(dòng)物房,動(dòng)物實(shí)驗(yàn)均遵守本醫(yī)院實(shí)驗(yàn)動(dòng)物管理委員會(huì)的相關(guān)規(guī)定。
2 主要試劑
蛇毒心臟毒素(cardiotoxin,CTX)購(gòu)自Merck;MLK3抗體(#4370S)、p-ERK1/2抗體(#4370)、ERK1/2抗體(#4695)、p-JNK抗體(#4668)、JNK抗體(#9252)、p-p38抗體(#4511)及p38抗體(#9212)均購(gòu)自Cell Signaling Technology;配對(duì)盒蛋白7(paired box protein 7,PAX7)抗體(AB_528428)購(gòu)自Developmental Studies Hybridoma Bank;麥胚凝集素(wheat germ agglutinin,WGA)抗體購(gòu)自Sigma;Alexa Fluor 555和Alexa Fluor 488熒光Ⅱ抗購(gòu)自Jackson ImmunoResearch;總RNA提取試劑TRIzol購(gòu)自Invitrogen;RevertAid First Strand cDNA Synthesis Kit(K1622)和T-PER組織蛋白提取液均購(gòu)自Thermo Fisher Scientific;SYBR Green Real-Time PCR試劑購(gòu)自TaKaRa;Masson染色液購(gòu)自北京索萊寶科技有限公司。
3 主要方法
3.1 骨骼肌損傷模型構(gòu)建10~12周齡的雄性WT及MLK3KO小鼠麻醉后,脛骨前肌注射CTX(10 μmol/L,30 μL)制備骨骼肌損傷模型[9]。
3.2 RNA提取及RT-qPCR使用TRIzol法提取成纖維細(xì)胞或小鼠脛骨前肌總RNA。使用RevertAid First Strand cDNA Synthesis Kit將2 μg總RNA逆轉(zhuǎn)錄成cDNA,步驟如下:RNA與Oligo dT混合后,65℃孵育5 min,加入逆轉(zhuǎn)錄酶及緩沖液,42℃60 min進(jìn)行逆轉(zhuǎn)錄反應(yīng),70℃5 min滅活逆轉(zhuǎn)錄酶。得到的cDNA在iQ5系統(tǒng)(Bio-Rad)中使用SYBR試劑進(jìn)行實(shí)時(shí)熒光定量PCR。以GAPDH為內(nèi)參照,目標(biāo)基因表達(dá)水平以2-ΔΔCt法進(jìn)行計(jì)算。MLK3的上游引物序列為5′-CCCTTTGCACAACTCATGGC-3′,下 游 引物 序列為5′-GAATGGAAGGAGTCCCGTGG-3′;Myog的上游引物序列為5′-CAGCCCAGCGAGGGAATTTA-3′,下游引物序列為5′-AGAAGCTCCTGAGTTTGCCC-3′;Myod的上游引物序列為5′-AGCATAGTGGAGCGCATCTC-3′,下游引物序列為5′-GGTCTGGGTTCCCTGTTCTG-3′;GAPDH的上游引物序列為5′-AATGCATCCTGCACCACC-3′,下游引物序列為5′-ATGCCAGTGAGCTTCCCG-3′。
3.3 Western blot實(shí) 驗(yàn)分離CTX處 理 的WT及MLK3KO小鼠脛骨前肌,T-PER組織裂解液(補(bǔ)充0.5 mol/L EDTA、蛋白酶抑制劑和磷酸酶抑制劑)提取總蛋白后,用BCA蛋白濃度測(cè)定試劑盒進(jìn)行蛋白濃度測(cè)定。將總量為80 μg的蛋白經(jīng)過(guò)聚丙烯酰胺凝膠電泳分離,然后電轉(zhuǎn)至硝酸纖維素膜。5%脫脂奶粉室溫封閉1 h,分別加入MLK3、p-ERK1/2、ERK1/2、p-JNK、JNK、p-p38、p38及GAPDH等抗體,4℃孵育過(guò)夜。加入Ⅱ抗室溫孵育2 h,使用Odyssey紅外熒光掃描成像儀捕獲目的條帶。目的條帶定量分析采用ImageJ軟件。
3.4 免疫熒光染色收集CTX處理的WT及MLK3KO小鼠脛骨前肌制備冰凍切片,用預(yù)冷的冰丙酮固定20 min,室溫晾干20 min后PBS洗,0.3%Triton X-100透化10 min,2次。3% BSA室溫封閉1 h,加入PAX7抗體,4℃過(guò)夜,PBS洗3次,加入Alexa Fluor 555熒光二抗,室溫孵育40 min,PBS洗后,DAPI封片。使用Leica ST5激光掃描共聚焦顯微鏡觀察PAX7的熒光強(qiáng)度。
3.5 WGA染色收集CTX處理的WT及MLK3KO小鼠脛骨前肌制備石蠟切片(4 μm),經(jīng)二甲苯和梯度乙醇脫蠟至水后,抗原熱修復(fù),羊血清工作液室溫封閉30 min,滴加1∶100稀釋的WGA抗體,37℃避光孵育1 h,PBS洗后,DAPI封片液封片。倒置熒光電子顯微鏡(Nikon)采集圖像,NIS Br 3.0軟件計(jì)算肌纖維橫截面積,每張切片統(tǒng)計(jì)200個(gè)肌纖維。
3.6 Masson膠原纖維染色將WT及MLK3KO小鼠脛骨前肌石蠟切片脫蠟至水。使用Masson染色液染色,基本步驟如下:鐵蘇木素染色5~10 min,酸性乙醇分化5~15 s;Masson藍(lán)化液返藍(lán)3~5 min;麗春紅品紅染色5~10 min;磷鉬酸溶液洗1~2 min;苯胺藍(lán)染色1~2 min;乙醇脫水、中性樹(shù)膠封片。倒置電子顯微鏡(Nikon)采集圖像,膠原纖維呈藍(lán)色,骨骼肌細(xì)胞呈紅色。NIS Br 3.0軟件計(jì)算纖維化面積。
4 統(tǒng)計(jì)學(xué)分析
用GraphPad Prism 7.0軟件進(jìn)行統(tǒng)計(jì)分析。結(jié)果以均數(shù)±標(biāo)準(zhǔn)差(mean±SD)表示。兩組間比較用t檢驗(yàn),多組間比較用單因素方差分析。以P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
結(jié) 果
1 骨骼肌損傷后早期MLK3表達(dá)水平顯著升高
為探討MLK3是否參與骨骼肌損傷后修復(fù),收集CTX處理0、1、3和5 d的小鼠脛骨前肌,RT-qPCR檢測(cè)MLK3的表達(dá)變化,發(fā)現(xiàn)CTX處理后3 d骨骼肌中MLK3的mRNA表達(dá)顯著升高(P<0.01),見(jiàn)圖1A;Western blot結(jié)果顯示,CTX處理后3 d的骨骼肌中MLK3蛋白表達(dá)升高,見(jiàn)圖1B。這提示MLK3可能在骨骼肌損傷后再生過(guò)程中發(fā)揮作用。

Figure 1.Mixed lineage kinase 3(MLK3)was up-regulated in regenerating tibial anterior(TA)muscles at early stage after cardiotoxin(CTX)-induced injury.A:MLK3 mRNA levels in TA muscles from wild-type mice at indicated time points after CTX injury(n=6);B:MLK3 protein expression in regenerating TA muscles at 3 d after injury(n=3).Mean±SD.**P<0.01 vs 0 d.圖1 MLK3在骨骼肌損傷修復(fù)早期表達(dá)升高
2 MLK3缺失導(dǎo)致骨骼肌損傷后再生能力下降
為探討MLK3是否在骨骼肌再生過(guò)程發(fā)揮作用,我 們 構(gòu) 建 了MLK3KO小鼠,Western blot驗(yàn)證MLK3KO小鼠骨骼肌纖維中無(wú)MLK3蛋白表達(dá)(圖2A)。進(jìn)而,WT及MLK3KO小鼠給予CTX處理誘導(dǎo)損傷。損傷后7 d,收集WT及MLK3KO小鼠骨骼肌,WGA染色觀察新生肌纖維大小及排列。未受損的WT及MLK3KO小鼠骨骼肌纖維排列緊密,肌纖維橫截面面積無(wú)差別;CTX誘導(dǎo)損傷7 d,MLK3KO小鼠骨骼肌纖維排列較對(duì)照組紊亂,新生的骨骼肌肌纖維橫截面面積顯著小于WT小鼠(P<0.01),見(jiàn)圖2B。這表明MLK3KO小鼠骨骼肌再生能力下降。
3 MLK3基因缺失加重骨骼肌損傷后纖維化
WT及MLK3KO小鼠脛骨前肌給予CTX處理后7 d,受損部位骨骼肌Masson染色評(píng)價(jià)膠原沉積情況。未損傷時(shí)MLK3KO和WT小鼠膠原纖維陽(yáng)性面積無(wú)顯著差異。損傷后7 d,MLK3KO和WT小鼠藍(lán)色膠原纖維陽(yáng)性面積均增加,但MLK3KO小鼠膠原纖維陽(yáng)性面積顯著高于WT小鼠(P<0.01),見(jiàn)圖3。這提示MLK3基因缺失加重骨骼肌損傷部位的纖維化。
4 MLK3基因缺失抑制肌衛(wèi)星細(xì)胞的增殖和分化
免疫熒光檢測(cè)WT及MLK3KO小鼠損傷后3 d骨骼肌中PAX7陽(yáng)性細(xì)胞數(shù),結(jié)果顯示MLK3KO小鼠骨骼肌中PAX7陽(yáng)性細(xì)胞比例顯著低于WT小鼠(P<0.01),見(jiàn)圖4A。RT-qPCR檢測(cè)肌衛(wèi)星細(xì)胞分化關(guān)鍵基因Myod和Myog的mRNA水平,結(jié)果顯示損傷后3 d的WT小鼠骨骼肌中Myod和Myog的表達(dá)較未損傷組顯著升高,但MLK3KO小鼠損傷后3 d的骨骼肌中Myod和Myog表達(dá)與WT小鼠相比均顯著降低(P<0.01),見(jiàn)圖4B。上述結(jié)果表明MLK3基因缺失抑制肌衛(wèi)星細(xì)胞的增殖和分化。
5 MLK3基因缺失抑制ERK1/2活化
Western blot檢 測(cè)CTX損 傷 后3 d的WT及MLK3KO小鼠骨骼肌中ERK1/2、JNK及p38磷酸化水平,結(jié)果顯示MLK3KO小鼠骨骼肌中磷酸化ERK1/2的水平與WT小鼠相比顯著下降(P<0.01),但磷酸化JNK及磷酸化p38的水平?jīng)]有顯著差異,見(jiàn)圖5。上述結(jié)果表明再生的骨骼肌中MLK3基因缺失抑制ERK1/2信號(hào)通路的激活。
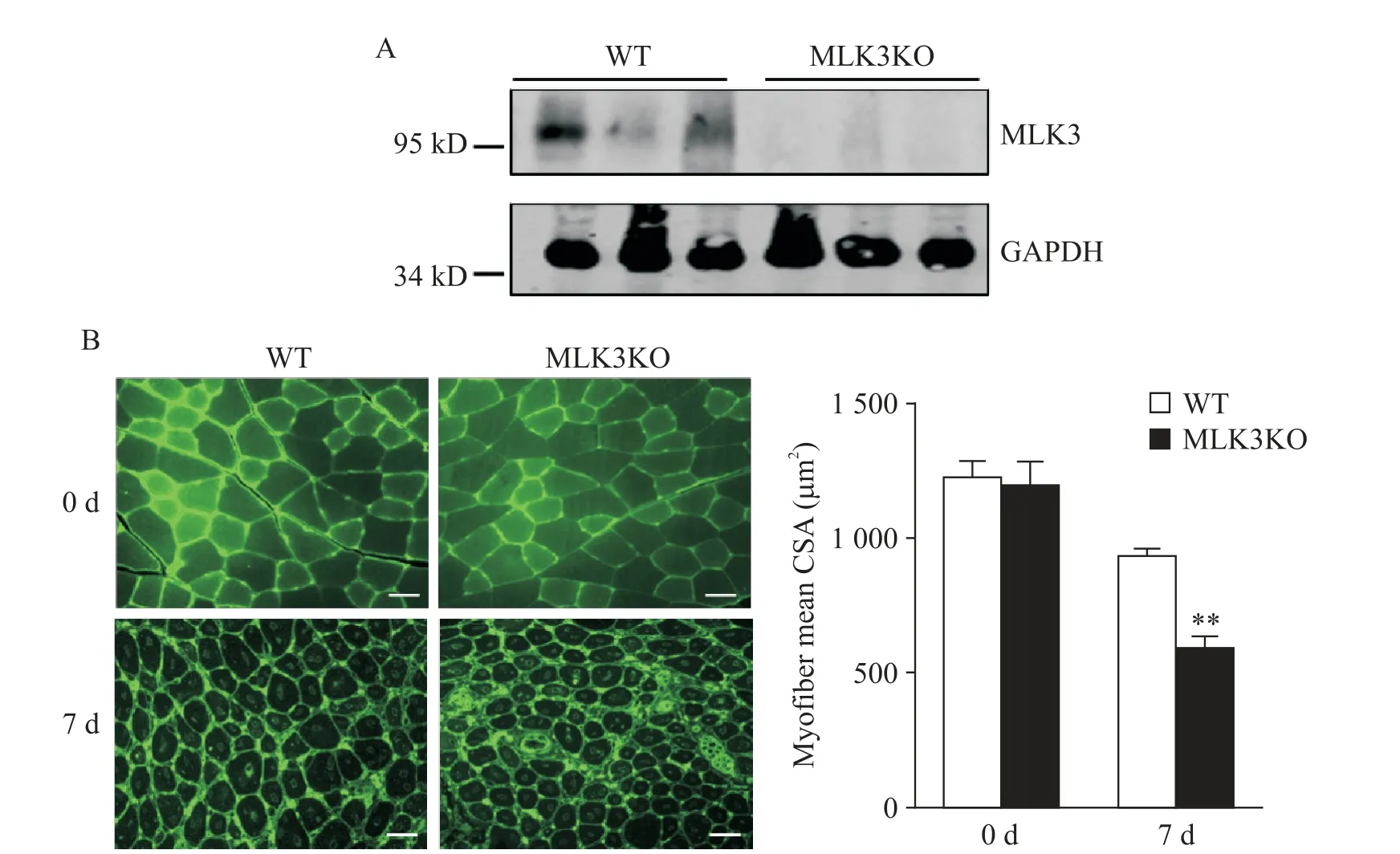
Figure 2.MLK3 deficiency resulted in impaired regeneration in injured muscle.A:Western blot analysis of MLK3 protein levels in muscles isolated from WT and MLK3KO mice;B:the WT and MLK3KO muscles were immunostained with WGA(green)at 0 and 7 d after CTX-induced injury(scale bar=20 μm),and the myofiber mean cross-sectional area(CSA)was analyzed(n=6).Mean±SD.**P<0.01 vs WT.圖2 MLK3基因缺失鼠骨骼肌損傷后再生能力下降
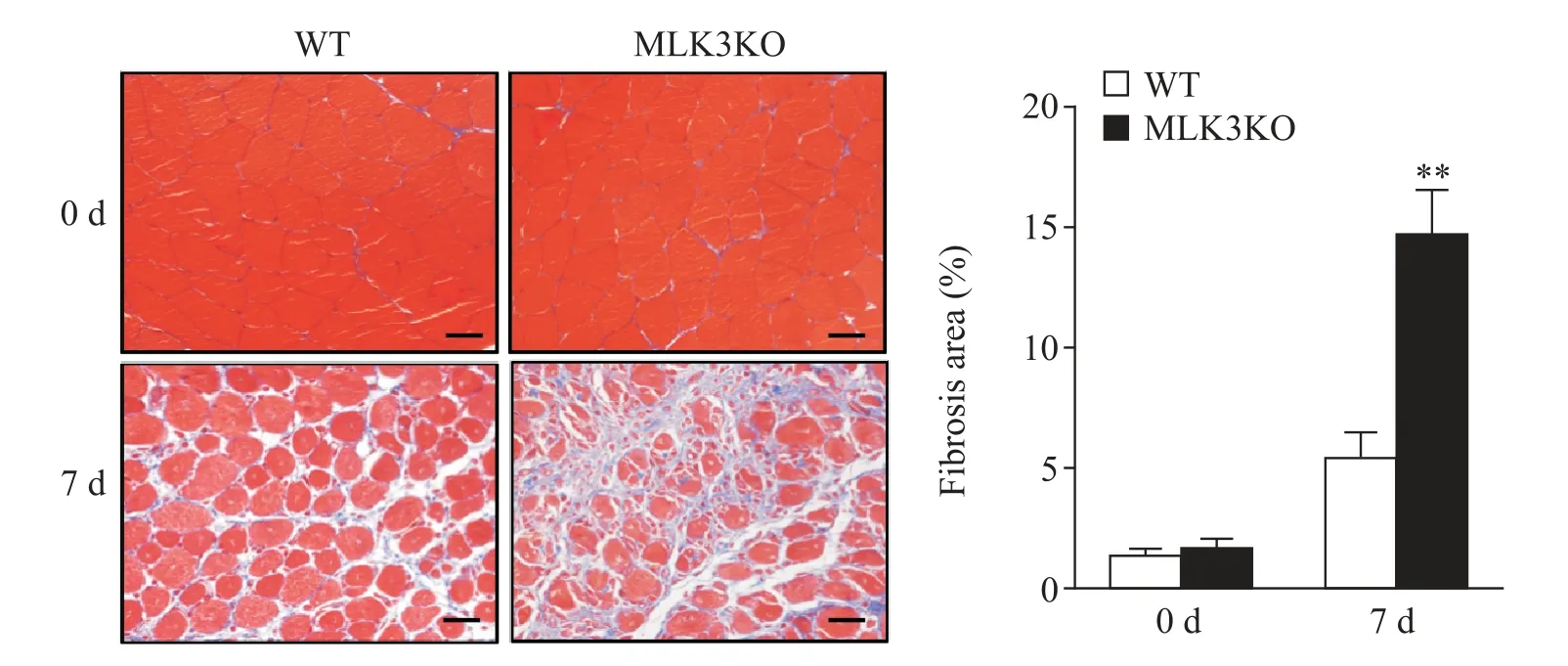
Figure 3.MLK3 deficiency resulted in increased muscle fibrosis in injured muscle.Masson staining was performed to detect collagen deposition(blue)in TA muscles isolated from WT and MLK3KO mice(scale bar=20 μm),and the ratio of fibrosis area per filed was calculated.Mean±SD.n=6.**P<0.01 vs WT.圖3 MLK3基因缺失加重骨骼肌損傷后纖維化
討 論
骨骼肌損傷后,肌衛(wèi)星細(xì)胞的活化、增殖和分化對(duì)于骨骼肌再生、促進(jìn)損傷后修復(fù),維持骨骼肌功能,阻止骨骼肌萎縮至關(guān)重要。本研究發(fā)現(xiàn),蛋白激酶MLK3基因缺失抑制肌衛(wèi)星細(xì)胞的增殖和分化,導(dǎo)致骨骼肌損傷后再生能力下降,間質(zhì)纖維化加重,表明MLK3在骨骼肌再生過(guò)程中發(fā)揮重要作用。
骨骼肌損傷后修復(fù)過(guò)程大致分為三個(gè)階段:炎癥期(數(shù)小時(shí)~7 d);再生期(2~7 d),肌衛(wèi)星細(xì)胞激活、增殖、分化、融合形成新的肌纖維;重塑期(5~30 d),細(xì)胞外基質(zhì)產(chǎn)生、新生肌纖維變大重塑等。其中,骨骼肌損傷后早期肌衛(wèi)星細(xì)胞增殖、分化對(duì)整個(gè)修復(fù)至關(guān)重要[10]。本研究發(fā)現(xiàn)CTX誘導(dǎo)損傷后早期(3 d)MLK3表達(dá)顯著升高,提示MLK3可能參與骨骼肌損傷后再生的早期細(xì)胞事件。

Figure 4.MLK3 deficiency impaired the proliferation and differentiation of muscle satellite cells.A:At 3 days after injury,PAX7 positive cells in WT and MLK3KO muscles were detected by immunofluorescence staining for PAX7(scale bar=20 μm),and percentages of PAX7 positive cells per field were analyzed(n=4);B:the mRNA levels of Myod and Myog in WT and MLK3KO muscles at 0 and 3 d after injury were assessed by RT-qPCR(n=6).Mean±SD.**P<0.01 vs WT.圖4 MLK3基因缺失抑制肌衛(wèi)星細(xì)胞增殖和分化

Figure 5.MLK3 deficiency inhibited phosphorylation of ERK1/2.Western blot analysis of the phosphorylation levels of ERK1/2,JNK and p38 in WT and MLK3KO muscles at 3 d after injury.Mean±SD.n=5.**P<0.01 vs WT.圖5 MLK3基因缺失抑制ERK1/2磷酸化
骨骼肌再生期肌衛(wèi)星細(xì)胞的激活、增殖、分化、融合受到生肌調(diào)節(jié)因子(myogenic regulatory factors,MRFs;包括PAX7、MyoD、MyoG等)的序貫表達(dá)所嚴(yán)格調(diào)控。骨骼肌損傷后,靜息的PAX7陽(yáng)性肌衛(wèi)星細(xì)胞活化,轉(zhuǎn)化為PAX7陽(yáng)性、MyoD陽(yáng)性的肌母細(xì)胞,繼而分化成MyoG陽(yáng)性的肌細(xì)胞。單個(gè)核的肌細(xì)胞融合成多個(gè)核肌細(xì)胞,并與殘留受損的肌纖維融合,形成新的肌纖維,從而完成骨骼肌再生過(guò)程[11]。該過(guò)程受到細(xì)胞內(nèi)多種信號(hào)通路的調(diào)控。比如Notch信號(hào)激活通過(guò)抑制MyoD的表達(dá)而抑制分化[3]。MAPK信號(hào)通路在骨骼肌再生過(guò)程中發(fā)揮重要作用:血管緊張素1結(jié)合Tie-2受體,激活下游ERK1/2,促進(jìn)肌衛(wèi)星細(xì)胞的增殖分化[4]。胰島素樣生長(zhǎng)因子激活p38,活化的p38介導(dǎo)再生關(guān)鍵因子MyoD的表達(dá),促進(jìn)骨骼肌再生[5]。MLK3作為MAPK級(jí)聯(lián)通路中ERK1/2及p38上游的關(guān)鍵激酶,其對(duì)ERK1/2及p38的激活是否在骨骼肌再生過(guò)程中發(fā)揮作用既往未見(jiàn)研究報(bào)道。
骨骼肌損傷后,位于肌纖維基底膜和肌膜之間的肌衛(wèi)星細(xì)胞增殖、分化,形成新的肌纖維,從而完成骨骼肌再生。PAX7是肌衛(wèi)星細(xì)胞的標(biāo)志物,并且在肌衛(wèi)星細(xì)胞的增殖中發(fā)揮重要作用[12]。本研究在損傷后3 d的WT及MLK3KO小鼠骨骼肌中通過(guò)免疫熒光觀察肌衛(wèi)星細(xì)胞PAX7陽(yáng)性細(xì)胞比例,發(fā)現(xiàn)MLK3KO小鼠PAX7陽(yáng)性細(xì)胞比例下降。PAX7表達(dá)于靜息的肌衛(wèi)星細(xì)胞及由肌衛(wèi)星細(xì)胞分化而來(lái)的肌母細(xì)胞,PAX7陽(yáng)性細(xì)胞比例下降,提示肌衛(wèi)星細(xì)胞增殖及分化能力下降。損傷后3 d,MLK3KO小鼠骨骼肌中分化關(guān)鍵基因Myod和Myog的表達(dá)下降,進(jìn)一步證實(shí)了MLK3基因缺失導(dǎo)致肌衛(wèi)星細(xì)胞增殖及分化能力減弱。基于既往研究ERK1/2及p38活化促進(jìn)肌衛(wèi)星細(xì)胞增殖和分化及MLK3對(duì)ERK1/2和p38的激活作用,我們推測(cè)MLK3可能通過(guò)活化ERK1/2或p38促進(jìn)肌衛(wèi)星細(xì)胞的增殖、分化。Western blot檢測(cè)CTX損傷后3 d的WT及MLK3KO小鼠骨骼肌中磷酸化ERK1/2和p38水平,顯示MLK3KO小鼠中磷酸化ERK1/2水平下降,而p38磷酸水平?jīng)]有變化,提示MLK3通過(guò)激活ERK1/2促進(jìn)肌衛(wèi)星細(xì)胞的增殖和分化。MLK3可以激酶依賴和激酶非依賴的方式激活ERK1/2[6,13]。在肌衛(wèi)星細(xì)胞的增殖和分化過(guò)程中,MLK3通過(guò)何種方式激活ERK1/2還有待探討。另外,骨骼肌損傷再生過(guò)程中,何種機(jī)制介導(dǎo)了MLK3的表達(dá)升高也不清楚,后續(xù)需要深入探討。
綜上所述,本研究結(jié)果顯示蛋白激酶MLK3基因缺失通過(guò)抑制ERK1/2活化降低骨骼肌損傷后肌衛(wèi)星細(xì)胞的增殖和分化,并導(dǎo)致骨骼肌損傷后再生能力受損,表明MLK3通過(guò)ERK1/2信號(hào)通路在骨骼肌再生中發(fā)揮重要作用,調(diào)控該信號(hào)通路可能有望成為骨骼肌萎縮的治療新方向。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫(huà)刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
