駝乳源乳鐵蛋白嵌合肽對口腔致齲菌抗菌作用的初步探究
2023-02-02 02:22:46李夢雨伊麗吉日木圖
食品與發酵工業 2023年2期
李夢雨,伊麗,吉日木圖
(內蒙古農業大學 食品科學與工程學院,內蒙古 呼和浩特,010000)
齲齒是世界上最常見的慢性口腔疾病之一。口腔細菌可附著于牙齒表面形成牙菌斑生物膜,同時其產生的酸性物質能夠腐蝕牙齒礦物質,最終形成齲齒[1]。齲齒常發病于兒童期,由此引發牙疼影響進食,出現機體營養不良的現象,阻礙兒童的正常發育。所以控制細菌生物膜保護口腔牙齒礦物質對于預防口腔齲齒至關重要。然而近些年在臨床治療過程中傳統抗生素的過度使用和濫用,不僅改變口腔和腸道菌群,口腔環境也極易發生波動,使抗生素的抑菌作用顯著降低。因此尋找一種對齲齒起預防及治療作用且安全持續有效的新型抗菌劑,是保護口腔健康的關鍵之一。
抗菌肽(antimicrobial peptides, AMPs),因具有天然來源、快速殺菌、不易產生耐藥性等特點受到了廣泛關注。大部分抗菌肽是陽離子抗菌肽,具有兩親性,一般由12~50個氨基酸短鏈構成。常見的二級結構包括α-螺旋、β-折疊、環(β-發夾)和延伸結構。通常抗菌肽在水溶液中呈現非結構狀態,在與膜脂雙層相互作用時表現出最終構型。抗菌肽可以破壞細胞膜、穿過細胞與核酸結合、抑制蛋白質合成并阻止細胞壁的合成發揮殺菌作用[2]。已有研究表明抗菌肽可以靶向殺死病原細菌,保護口腔健康微生物群落[3]。TAO等[4]發現口腔中的天然抗菌肽如防御素、導管素、組蛋白等,具有保護牙齒和口腔黏膜的作用。MAI等[5]報道抗菌肽對變異鏈球菌等口腔病原體和生物膜具有抗菌活性。然而,由于抗菌肽的半衰期較短且在體內的穩定性較低,大大限制了抗菌肽的應用。研究人員正在開發具有良好穩定性和低細胞毒性的抗菌肽。合成抗菌肽可以安全有效地維持口腔穩態保護機體健康,因此合成抗菌肽作為一種新型抗菌療法具有極大的潛力。
乳鐵蛋白(lactoferrin, LF),作為一種轉鐵蛋白,具有抗菌、抗病毒等特性,不僅構成宿主非特異性免疫系統,也是口腔唾液防御體系中的重要組成部分。LF在酸性條件下經胃蛋白酶消化得到lactoferricin(LFcin),在蛋白水解條件下得到lactoferrampin(LFampin)。LFcin具有以β-折疊為主的兩親性結構,更易黏附于帶負電荷的細胞膜,且LFampin的氨基端存在螺旋帽結構,能夠增加α-螺旋穩定性。因此與LF相比,LFcin和LFampin抗菌效果更強[6]。研究表明,在牙科領域,帶正電的乳鐵蛋白與革蘭氏陰性菌細胞壁的脂多糖結合導致變異鏈球菌等口腔細菌在牙齒表面的黏附功能障礙[7]。駝乳作為一種功能性乳品,具有低膽固醇、低糖,礦物質和脂肪酸種類及含量豐富且維生素C含量高等特點。駝乳中LF的含量為200~1 000 mg/L。與除人乳以外的其他動物乳相比,駝乳中乳鐵蛋白的含量明顯偏高,是牛乳的2.4倍、山羊乳的2.2倍[8]。
目前,國內主要通過提取純化獲取駝乳源生物活性肽并分析其營養特性和功能特性,在人工合成駝乳生物活性肽方面尚處于空白階段。此外,天然駝乳源乳鐵蛋白活性肽來源困難,成本昂貴,而LFcin和LFampin抗菌效果更強。因此,本研究通過化學合成駝乳源LFA-LFC嵌合肽,并探究其對3種主要致齲菌[變異鏈球菌(Streptococcusmutans)、唾液鏈球菌(Streptococcussalivarius)、遠緣鏈球菌(Streptococcussobrinus)]的抗菌能力及其作用機制。
1 材料與方法
1.1 材料與試劑
駝乳源LFA-LFC嵌合肽(純度≥98%),上海楚肽生物科技有限公司采用化學固相合成法合成;S.mutansATCC 2517、S.salivariusATCC 13419、S.sobrinusATCC 33478,廣東省科學院微生物研究所;胰蛋白胨大豆肉湯(瓊脂)、營養肉湯(瓊脂)、腦心浸液肉湯(瓊脂),青島海博生物有限公司;PBS緩沖溶液、十二烷基硫酸鈉(sodium dodecyl sulfate, SDS),北京索萊寶科技有限公司;氯己定,上海易恩化學技術有限公司;細菌基因組DNA提取試劑盒(離心柱型),北京天根生化科技有限公司;所有無機溶劑均為國產分析純。
1.2 儀器與設備
博迅凈化工作臺,上海博迅醫療生物儀器股份有限公司;SQ810C蒸汽滅菌器,日本YAMATO公司;恒溫振蕩器、隔水式培養箱,上海一恒科學儀器有限公司;FE28臺式酸度計,梅特勒-托利多國際貿易有限公司;Synergy H1酶標儀、Gel-Doc XR+凝膠成像系統,美國Biotek公司;5810 R冷凍離心機,德國Eppendorf公司;小型水平電泳槽,美國BIO-RAD公司。
1.3 實驗方法
1.3.1 駝乳源LFA-LFC嵌合肽的合成及理化性質
本研究委托上海楚肽生物科技有限公司采用化學固相合成法,通過嵌入一個帶正電荷的賴氨酸連接乳鐵蛋白第265~284位氨基酸(LFampin)和17~30位氨基酸(LFcin)合成駝乳源LFA-LFC嵌合肽,純度≥98%。通過PreCHemPeptide軟件對駝乳源LFA-LFC嵌合肽進行分子特性的預測。使用質譜儀、高效液相色譜以及圓二色譜(circular dichroism, CD)法進行鑒定分析。
1.3.2 駝乳源LFA-LFC嵌合肽抗菌特性的測定
1.3.2.1 駝乳源LFA-LFC嵌合肽最低抑菌濃度、最低殺菌濃度的測定
按照美國試驗室標準微量肉湯稀釋法測定駝乳源LFA-LFC嵌合肽對S.mutans、S.salivarius和S.sobrinus3株菌的最低抑菌濃度(minimum inhibitory concentration, MIC)和最低殺菌濃度(minimum bactericidal concentration, MBC)。最低抑菌濃度是指肉眼觀察沒有細菌生長和OD600值無變化狀態時嵌合肽的濃度,最低殺菌濃度是指99.9%的細菌被抑制時的嵌合肽濃度。
將處于對數生長期的菌懸液用培養基稀釋至2×106CFU/mL。將嵌合肽濃度進行二倍梯度稀釋為2、4、8、16、32、64、128、256、512 μmol/L。試驗組每孔加入嵌合肽100 μL和菌懸液10 μL混合;陰性組為培養基100 μL和菌懸液10 μL混合;陽性組為0.2%(質量分數)氯己定100 μL和菌懸液10 μL混合;空白對照組為培養基100 μL和無菌水10 μL混合。將96孔板置于振蕩培養箱,37 ℃,50 r/min,培養24 h。培養結束后使用酶標儀在600 nm處測定混合液光密度值(OD600)。
以嵌合肽最低抑菌濃度為依據,配制1~8倍的MIC嵌合肽濃度,與稀釋至2×106CFU/mL的細菌菌懸液在37 ℃,50 r/min條件下共培養24 h。每孔取10 μL混合液涂布于瓊脂平板,倒置于恒溫培養箱,37 ℃,培養24 h。
1.3.2.2 不同pH培養基對駝乳源LFA-LFC嵌合肽抗菌效果的影響
參考杜鵑[9]的研究方法,使用1 mol/L HCl或1 mol/L NaOH調節液體培養基pH值分別為6.3、6.6、6.8、7.0、7.3、8.3 6個梯度。參照1.3.2.1測定MIC。
1.3.2.3 菌體時間-致死曲線的測定
將處于對數生長期的菌懸液用培養基稀釋至2×106CFU/mL。根據1.3.2.1試驗結果,選擇嵌合肽濃度為MIC、2×MIC。試驗組加入菌懸液與嵌合肽1∶1比例混合,陰性對照組加入菌懸液與無菌水混合。分別在0、1、2、5、15、30、60、120、240 min時移取菌懸液于0.85%(質量分數)無菌生理鹽水中,稀釋104倍,振蕩混勻后立即置于冰面,使菌體停止生長。之后取100 μL細菌稀釋液涂布平板,并標注日期、菌種,放進恒溫培養箱,37 ℃,培養24 h。培養結束后,計算菌落數,以時間為橫坐標,菌落數的對數值為縱坐標繪制時間-致死曲線。
1.3.3 駝乳源LFA-LFC嵌合肽殺菌作用機制的研究
1.3.3.1 預防菌體生物膜生成試驗
采用結晶紫染色法測定生物膜。將處于對數生長期的菌懸液用培養基稀釋至2×106CFU/mL。二倍梯度稀釋嵌合肽濃度為2、4、8、16、32、64、128 μmol/L。將菌懸液與不同濃度嵌合肽按照10∶1的比例混合于96孔板中,陰性對照組加入培養基與菌懸液混合,空白對照組加入無菌水和培養基混合。吹打混勻后,標注日期、菌種,置于恒溫培養箱,37 ℃,培養24 h。培養結束,輕輕吸去培養基上清液,每孔加入200 μL無菌0.85%生理鹽水,溫和沖洗3次,室溫晾干后加入200 μL 10%(體積分數)甲醇固定10 min,使附著的生物膜更牢固。吸去甲醇,加入100 μL 0.1%(質量分數)結晶紫,進行30 min染色,然后吸去結晶紫染色液,使用0.85%無菌生理鹽水溫和沖洗3次,室溫晾干。最后每孔加入150 μL 33%(體積分數)乙酸,反應10 min以溶解生物膜。將孔內溶液移至新的96孔酶標板中測定OD600值。生物膜形成量的計算如公式(1)所示:
(1)
1.3.3.2 消除菌體生物膜生成試驗
將處于對數生長期的菌懸液用培養基稀釋至2×106CFU/mL。二倍梯度稀釋嵌合肽濃度至8、16、32、64、128、256、512 μmol/L。使用96孔酶標板,試驗組、陰性組每孔加入200 μL菌懸液,空白組加入200 μL培養基,置于恒溫培養箱,37 ℃,培養24 h,使菌體形成生物膜。培養結束后,輕輕吸去96孔板中的懸濁液,加入不同濃度嵌合肽100 μL,陰性組和空白對照組分別加入培養基和無菌水代替嵌合肽。37 ℃,過夜培養。參照1.3.3.1測定生物膜剩余量。
1.3.3.3 透射電子顯微鏡觀察
將處于對數生長期的菌懸液用培養基稀釋至2×106CFU/mL。加入128 μmol/L駝乳源LFA-LFC嵌合肽和菌懸液以1∶1的比例混勻,陰性對照組加入無菌水和菌懸液混合。37 ℃,過夜培養。然后在4 500 r/min的條件下,離心5 min。棄掉上清液,加入PBS溶液清洗沉淀物,再次離心。重復沖洗過程2次后,加入2.5%(體積分數)戊二醛固定液后,至4 ℃冰箱固定12 h以上。將樣品送至邁斯普生物科技有限公司使用JEM1230型透射電鏡(transmission electron microscope, TEM)觀察,采集圖像進行分析。
1.3.3.4 激光共聚焦電子顯微鏡觀察
使用異硫氰酸熒光素(fluorescein isothiocyanate isomer, FITC)作為熒光探針標記駝乳源LFA-LFC嵌合肽(FITC-嵌合肽)委托上海楚肽生物科技有限公司合成。將處于對數生長期的菌懸液用培養基稀釋至2×106CFU/mL。將無菌PBS溶液與菌懸液以1∶1的比例混勻,在1 000 r/min,4 ℃條件下,離心30 s,棄掉上清液后,重懸于無菌PBS溶液中。使用Hoechst 33258(10 μg/mL,30 min)和尼羅紅(1 μmol/L,30 min)染料連續染色細菌細胞。沖洗后加入128 μmol/L FITC-嵌合肽作用30 min。然后加入100 μL 2.5%戊二醛,固定20 min。再次輕輕沖洗樣品,并將細菌沉淀物重懸于無菌PBS溶液中。取樣品置于蓋玻片,通過激光共聚焦電子顯微鏡(confocal laser scanning microscope, CLSM)觀察。
1.3.3.5 菌體DNA電泳遷移率的測定
將處于對數生長期的菌懸液用培養基稀釋至2×106CFU/mL。使用細菌基因組DNA提取試劑盒提取菌體DNA,通過微量分光光度計測定DNA純度和濃度,凍存于-20 ℃冰箱備用。二倍梯度稀釋嵌合肽濃度為2、4、8、16、32、64、128 μmol/L。將不同濃度的嵌合肽和菌體DNA以1∶4的比例混勻,陰性對照組加入無菌水和菌體DNA。37 ℃,孵育5 min后,加入2 μL 6×loading buffer染料,然后在120 V,30 min條件下,進行1%瓊脂糖凝膠核酸電泳檢測。電泳結束后,利用UV凝膠成像系統觀察菌體DNA遷移率,分析其在凝膠中的阻滯情況。
1.3.4 駝乳源LFA-LFC嵌合肽溶血性的測定
參照文獻[10]對駝乳源LFA-LFC嵌合肽溶血性進行測定。取適量無菌綿羊血于無菌離心管中,在4 ℃、4 000 r/min的條件下,離心10 min。棄掉上清液,將綿羊血紅細胞沉淀與無菌PBS溶液以1∶1比例混勻,在4 ℃、4 000 r/min條件下,離心10 min。重復沖洗過程4次后,紅細胞于4 ℃冰箱低溫貯存備用。
將嵌合肽與綿羊血紅細胞以9∶1的比例混勻,陽性組加入SDS溶液和綿羊血紅細胞混勻,空白組加入PBS溶液和綿羊血紅細胞混勻。分別置于恒溫培養箱,在37 ℃條件下,反應3 h。反應結束后,在4 ℃、4 000 r/min條件下,離心10 min。取上清液100 μL加入96孔酶標板中,通過酶標儀測定OD540值。溶血率的計算如公式(2)所示:

(2)
1.4 數據處理與分析
本研究中所有試驗均重復3次。試驗數據采用IBM SPSS Statistic 26軟件和Microsoft Office Excel 2020軟件進行統計分析,以平均值±標準差進行表示,P<0.05代表差異具有統計學意義。
2 結果與分析
2.1 駝乳源LFA-LFC嵌合肽的合成及理化性質
本研究采用高效液相色譜鑒定嵌合肽純度達到98%以上。通過質譜儀和軟件分析對嵌合肽的分子特性進行檢測。結果表明,駝乳源LFA-LFC嵌合肽分子質量為4 357.26 g/mol,等電點為11.60,凈電荷值為10.9,親水殘基比達到25%。研究發現抗菌肽主要通過自身所帶正電荷與細菌細胞膜所帶負電荷區域發生靜電作用,進而發揮抑菌效果[11]。由此可以看出抗菌肽的凈電荷值與疏水性影響其抗菌特性。嵌合肽的凈電荷值與親水殘基比越高,其抗菌性能越強。天然陽離子抗菌肽所帶正電荷數一般為2~9個[1]。梁東升等[12]設計篩選得到的抗變異鏈球菌多肽AT-7凈電荷值為5,已經具有良好的抗菌效果。與之相比,駝乳源LFA-LFC嵌合肽所帶正電荷更多。
抗菌肽中存在不同的二級構象,影響抗菌肽的空間結構進而影響其功能穩定性。為了進一步探究駝乳源LFA-LFC嵌合肽的功能特性,本研究通過CD測定其二級結構,分別包含:12.38%的α-螺旋結構、40.66%的β-折疊結構、15.24%的β-轉角結構和31.65%的無規則卷曲。其中α-螺旋和β-折疊占比達到50%以上。與TU等[13]對合成肽GH12進行二級結構的鑒定結果相似。鑒于上述研究結果發現駝乳源LFA-LFC嵌合肽具有一定的抗菌特性潛能,具備進一步探究的價值。
2.2 駝乳源LFA-LFC嵌合肽的合成及理化性質
2.2.1 駝乳源LFA-LFC嵌合肽MIC、MBC的測定
MIC試驗結果顯示嵌合肽對S.mutans、S.salivariu的抑菌效果更強,MIC值達到32 μmol/L,比S.sobrinus的MIC值低2倍。MBC試驗結果與上述結果一致,S.mutans、S.salivariu的MBC值較小,均為128 μmol/L,而S.sobrinus的MBC值達到256 μmol/L(表1)。由此可知,該嵌合肽對3株致病菌均表現出抑菌活性。
鐘亨任[14]研究表明抗菌肽GHc和GHd對變異鏈球菌的MIC值分別為12.6、13.1 μmol/L。其MIC值較低可能與抗菌肽的二級結構有關,α-螺旋含量較高,可以更有效地發揮抗菌作用。而何佳寧等[15]合成得到的短鏈抗菌肽KR-1對S.sobrinus菌的MIC值為51.2 μmol/L,與本研究中嵌合肽的抗菌效果相似。

表1 駝乳源LFA-LFC嵌合肽MIC、MBC的測定Table 1 Determination of the LFA-LFC chimeric peptide MIC、MBC
2.2.2 不同pH培養基對駝乳源LFA-LFC嵌合肽抗菌效果的影響
口腔是復雜的動態環境,正常生理條件下口腔唾液的pH值維持在6.6~7.1[16]。因此本研究以口腔環境酸堿度為依據,測定不同pH培養環境下嵌合肽對S.mutans、S.salivariu、S.sobrinusMIC值的變化。結果發現不同pH值對嵌合肽的MIC值有較為顯著的影響(表2)。pH值為6.3~7.1時,各細菌的MIC值沒有發生變化,均為64 μmol/L。隨著pH值增大,嵌合肽對3株菌均呈現出更強的抗菌效果。當pH值降低為8.3時,3株菌的MIC值全部縮小4倍。這一結果符合鏈球菌產酸耐酸的生理特性。同時說明堿性環境更有利于嵌合肽發揮對口腔致齲菌的抑制作用。杜鵑[9]研究結果表明,pH上升有利于抗菌肽抗菌活性的增強,與本研究結果一致。營秀[17]研究唾液對抗菌肽pm11抗菌作用的影響,發現pm11溶解在唾液或超純水中抑菌效果在統計學上無差異。而駝乳源LFA-LFC嵌合肽在pH值6.3~7.1時,MIC值無變化。當pH值達到7.3及以上,其抑菌效果增強。這可能是相較于培養基,口腔環境中唾液成分更復雜,對抗菌肽的抑菌效果產生干擾。
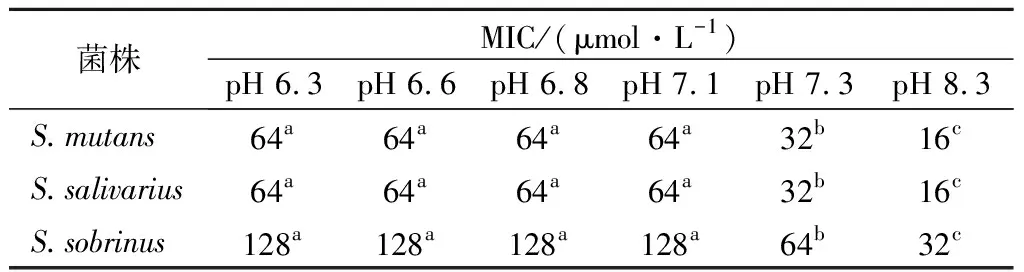
表2 不同pH值駝乳源LFA-LFC嵌合肽MIC的測定Table 2 Determination of the LFA-LFC chimeric peptide MIC at different pH
2.2.3 菌體時間-致死曲線的測定
駝乳源LFA-LFC嵌合肽對S.mutans、S.salivariu和S.sobrinus3株菌均表現出顯著的殺菌效果(圖1)。縱向比較嵌合肽濃度對3株菌的抑菌效果,可以發現1×MIC濃度嵌合肽對S.mutans、S.salivariu和S.sobrinus在30 min時,菌體全部到達致死狀態。而2×MIC濃度嵌合肽對S.mutans、S.salivariu和S.sobrinus分別為5、2、15 min時,菌體已經到達完全致死狀態。嵌合肽濃度越高,其對細菌的抑制作用越明顯。橫向比較嵌合肽對3株菌的抑菌效果亦存在強弱差,其中嵌合肽對S.salivariu的抑制作用最強,對S.sobrinus的抑制作用最弱。該結果與2.2.1對嵌合肽的MIC、MBC的測定結果一致。
魏詩[18]研究發現,抗菌肽濃度對S.mutans殺菌時間及程度呈濃度依賴現象。同時TU等[13]研究結果表明2×MIC抗菌肽GH12在60 min內將S.mutans全部殺死,4×MIC只需要20 min即可使S.mutans致死。這與本研究結果一致,高濃度駝乳源LFA-LFC嵌合肽對細菌的抑制效果更迅速。
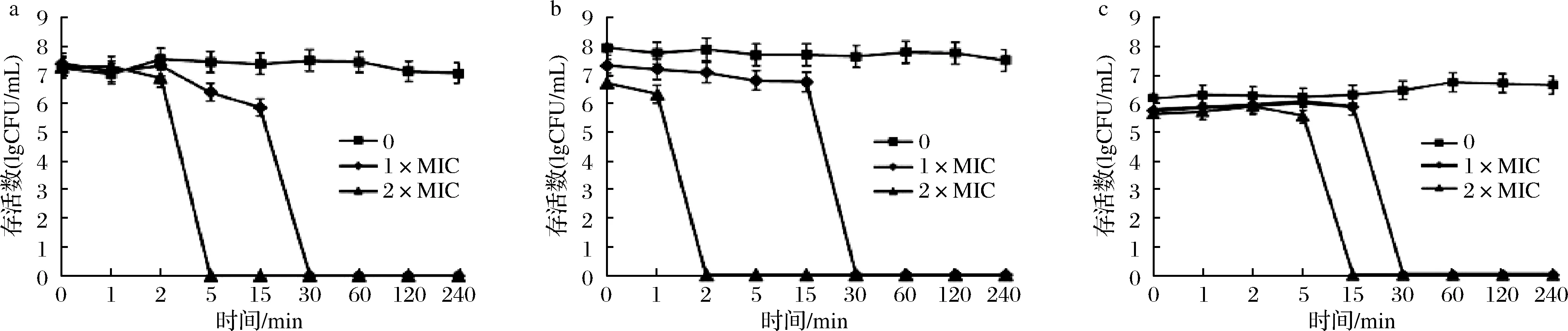
a-S.mutans;b-S.salivariu;c-S.sobrinus圖1 時間-致死曲線Fig.1 Time-Killing curves
2.3 駝乳源LFA-LFC嵌合肽殺菌作用機制的研究
2.3.1 預防菌體生物膜生成試驗
人體口腔內的病原菌可在口腔內形成生物膜使病原微生物在牙齒表面附著,且生物膜的酸化現象可導致牙齒脫礦,加速口腔疾病的發展。因此破壞口腔內生物膜的生成是預防齲齒的重要環節。本研究將嵌合肽與口腔致齲菌共培養探究嵌合肽對口腔內生物膜形成的影響。研究發現,當嵌合肽濃度達到16 μmol/L時,生物膜生成量開始有所減少。在嵌合肽濃度達到128 μmol/L時,S.mutans、S.salivariu的生物膜生成量減少約80%,幾乎被完全抑制;雖然嵌合肽對預防S.sobrinus生物膜生成作用較弱,但減少量也較為顯著,達到70%以上(圖2)。由此可知,高濃度嵌合肽有效預防生物膜的生成。這是由于嵌合肽帶正電荷,與革蘭氏陽性菌細胞表面帶負電荷的磷壁酸發生靜電作用,抑制細菌生長繁殖。進一步損害生物膜的完整性,破壞細菌的生存環境。
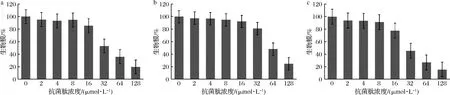
a-S.mutans;b-S.salivariu;c-S.sobrinus圖2 預防變異鏈球菌、唾液鏈球菌、遠緣鏈球菌生物膜的形成Fig.2 Inhibition of biofilm formation of S.mutans, S.salivariu, and S.sobrinus
ZHANG等[19]通過掃描電子顯微鏡觀察牙釉質表面生物膜的生成發現,抗菌肽DPS-PI處理后的切片表面菌體稀疏分布,表明DPS-PI可以有效抑制牙釉質生物膜的形成。李欣蔚等[20]研究發現抗菌肽GH12對口腔致齲菌生物膜同樣表現出一定的損傷作用。這與本研究的試驗結果一致,證明駝乳源LFA-LFC嵌合肽對預防生物膜生成發揮較為顯著的作用,具備維護口腔健康的潛能。
2.3.2 消除菌體生物膜試驗
駝乳源LFA-LFC嵌合肽對口腔致齲菌已形成的生物膜表現出一定的消除作用(圖3)。隨著嵌合肽濃度增大,生物膜剩余量逐漸減少。當嵌合肽濃度達到512 μmol/L時,嵌合肽對S.mutans、S.salivariu已形成的生物膜消除率可達到約50%,對S.sobrinus已形成的生物膜消除率可達到約40%。本研究結果雖然表明嵌合肽可以減緩已形成的生物膜發展,但是消除效果并不理想。
LEIVA-SABADINI等[21]研究發現從蜂蜜中獲得的抗菌肽Hec-Evs盡管對變異鏈球菌生物膜表現出一定程度的抑制,但仍然可以黏附在表面并建立暴露于較低濃度抗菌肽時的早期生物膜。而賈麗麗[22]的研究也表明抗菌肽對生物膜的消除作用較弱,需要機械作用輔助進行清理。因此我們推測這可能與嵌合肽的抗菌機制有關。針對已形成的生物膜,需要尋找另外一種高效率的方法代替。
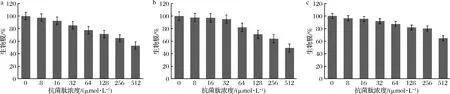
a-S.mutans;b-S.salivariu;c-S.sobrinus圖3 消除生物膜試驗Fig.3 Eradication of preformed biofilm
2.3.3 TEM觀察
通過TEM觀察駝乳源LFA-LFC嵌合肽對不同菌體的作用效果(圖4)。未經嵌合肽處理的細菌細胞結構完整,細胞膜清晰可見,尤其是細胞質區域的電子密度致密均勻。與之相比,經128 μmol/L嵌合肽處理后,3株細菌的細胞均呈現不同程度的結構變化。S.mutans細胞膜已經完全溶解,細胞內部開始出現孔洞。S.salivariu細胞膜明顯出現泡沫狀裂解,細胞質區域的電子密度減小且不均勻。S.sobrinus細胞膜喪失完整性,出現細胞質壁分離現象,細胞質內容物大量泄漏。TEM結果表明駝乳源LFA-LFC嵌合肽可能通過作用于細胞膜發揮抑菌或殺菌的作用。這與耿紅娟[23]探究雙功能嵌合肽(TBP-1-GGG-hBD3-3)對口腔鏈球菌抗菌機制的研究結果一致。
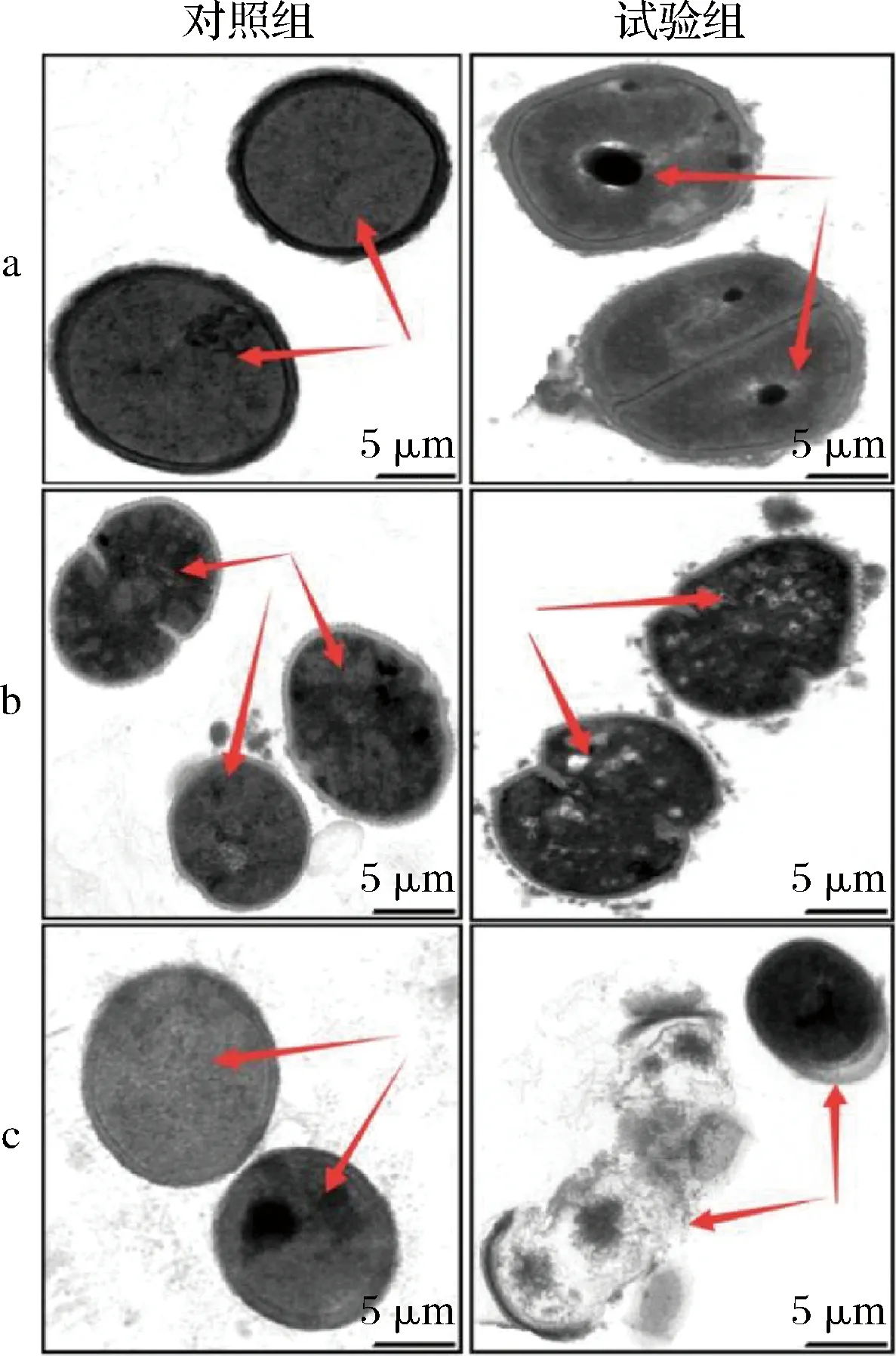
a-S.mutans;b-S.salivariu;c-S.sobrinus圖4 菌體TEM圖像Fig.4 TEM images of bacterial cells
2.3.4 CLSM觀察
Hoechst 33258和尼羅紅在熒光染色過程中,Hoechst 33258可以進入細胞內部與DNA結合,在CLSM下觀察可看到藍色熒光;尼羅紅作為一種親脂性染料,可以與細胞膜中的脂質成分結合,在CLSM下觀察可看到紅色熒光;FITC是生物學應用廣泛的綠色熒光素衍生物,常用作蛋白標記。FITC-嵌合肽在CLSM下觀察可看到綠色熒光。通過CLSM成像進一步定位駝乳源LFA-LFC嵌合肽在細菌細胞內部的位置(圖5)。可以看到,未經處理的細菌細胞周圍被帶有紅色熒光的細胞膜完整環繞,細胞內部存在大量帶有藍色熒光的核酸物質。經128 μmol/L FITC-嵌合肽處理后,細胞膜被破壞,紅色熒光逐漸消失,大部分綠色熒光在細胞質中累積,而藍色熒光減少。CLSM結果表明駝乳源LFA-LFC嵌合肽可以穿過細胞膜障礙并滲透到細胞內部。
WANG等[24]使用FITC作為熒光探針標記抗菌肽F1(綠色),FM-4-64(紅色)標記細胞膜,并使用CLSM觀察FITC-F1在細胞中的位置變化。結果發現FITC-F1滲入細胞,隨著時間的延長在細胞質中累積,最終細胞膜消失,FITC-F1覆蓋全部細胞質。這與本研究結果一致。
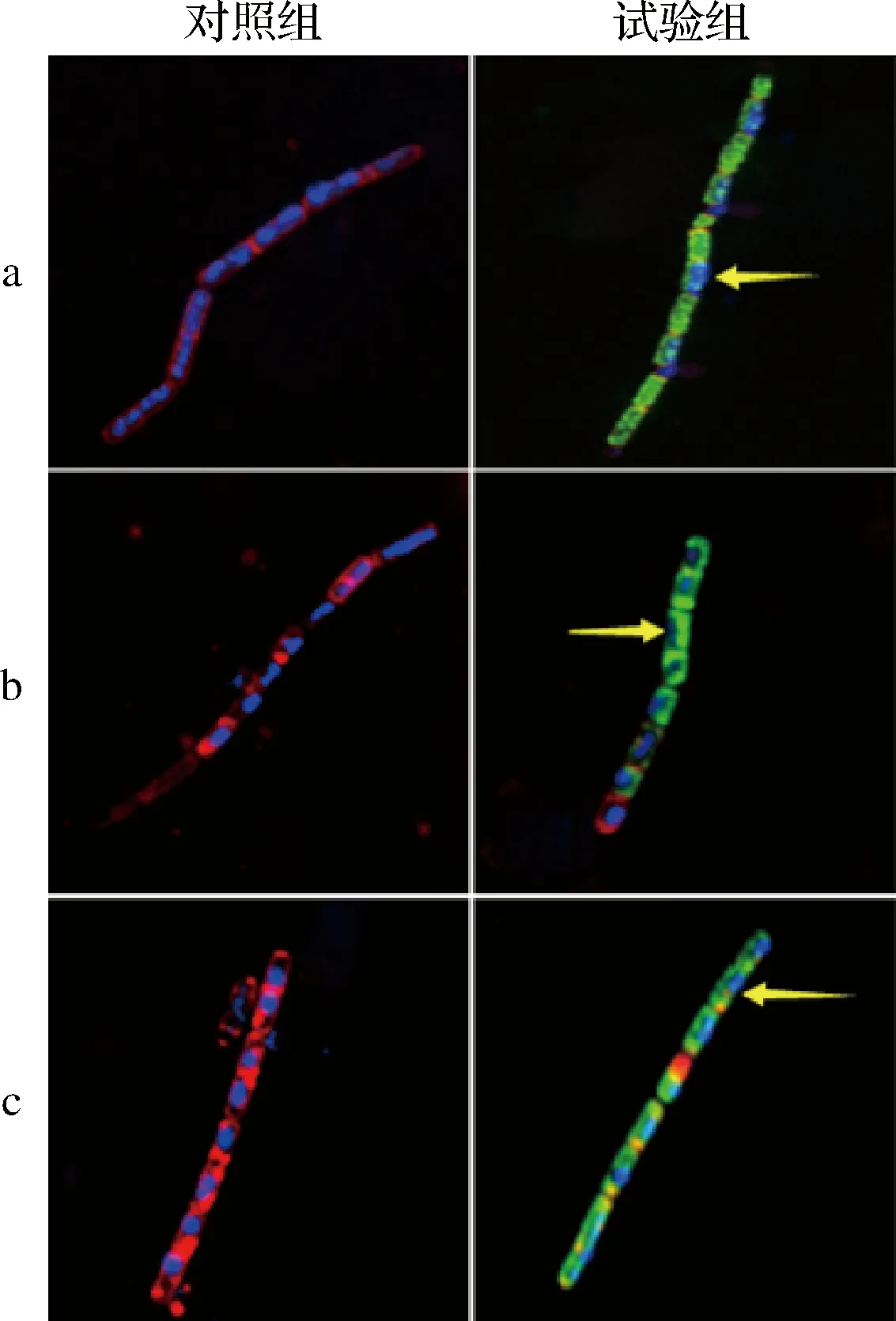
a-S.mutans;b-S.salivariu;c-S.sobrinus圖5 菌體CLSM圖像Fig.5 CLSM images of bacterial cells
2.3.5 菌體DNA電泳遷移率的測定
由圖6可知,在嵌合肽的作用下DNA發生不同程度的遷移。當嵌合肽濃度分別在0~8、0~8、0~16 μmol/L時,S.mutans、S.salivariu和S.sobrinus菌體DNA遷移幾乎沒有受到阻滯,在泳道中出現明亮條帶。S.mutans和S.salivariu菌體DNA在嵌合肽濃度達到16 μmol/L時,遷移逐漸受到阻滯;在32 μmol/L嵌合肽濃度時,S.sobrinus菌體DNA遷移開始受到阻滯,部分DNA阻滯在凝膠孔中,泳道條帶亮度減弱。在64 μmol/L時,S.salivariu菌體DNA被全部阻滯在凝膠孔中,未在泳道中發現遷移條帶;而S.mutans和S.sobrinus菌體DNA在128 μmol/L嵌合肽濃度時被全部阻滯。已有文獻報道,菌體DNA是大多數陽離子抗菌肽的理想結合位點[25]。這主要是由于DNA帶負電,陽離子抗菌肽與DNA結合,使DNA受到損傷。DNA作為生命活動重要的遺傳物質,其完整性是生命發育和運轉的必要條件。由此可知,駝乳源LFA-LFC嵌合肽作用于細菌DNA,使之在遷移過程中出現阻滯現象,抑制DNA遺傳復制,影響細菌正常的生長繁殖。
杜鵑[9]研究表明高濃度抗菌肽P26才能與細菌DNA完全結合,發揮抑制生長的作用。這也能證明駝乳源LFA-LFC嵌合肽可以通過進入細菌內部與DNA結合發揮良好的抑菌作用。
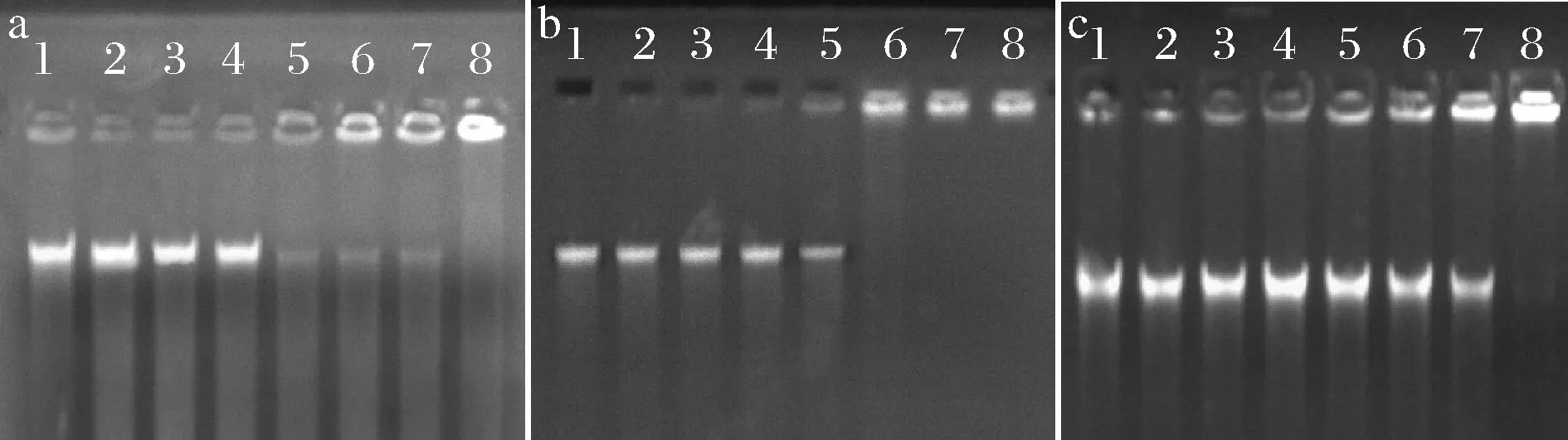
泳道1~8分別為嵌合肽濃度0、2、4、8、16、32、64、128 μmol/L的試驗組 a-S.mutans;b-S.salivariu;c-S.sobrinus圖6 菌體DNA電泳遷移率Fig.6 Electrophoretic mobility of bacterial DNA
2.4 駝乳源LFA-LFC嵌合肽溶血性的測定
本研究通過測定綿羊血紅細胞溶血率評估駝乳源LFA-LFC嵌合肽對哺乳動物細胞的損傷性。結果發現,嵌合肽濃度對溶血率存在一定影響(圖7)。當嵌合肽濃度達到32 μmol/L時,作用3 h,嵌合肽的溶血率僅達到約20%,之后溶血率上升趨勢減緩。練家惠等[10]通過改造溶血率較高的天然抗菌肽Temporin-1Dra,得到新的合成肽溶血率在2~4×MIC下仍低于10%,明顯低于駝乳源LFA-LFC嵌合肽。有文獻表示,抗菌肽所帶電荷數對其溶血率產生影響,正電荷數越高,溶血現象越嚴重[10]。因此我們猜測駝乳源LFA-LFC嵌合肽溶血率相對較高可能是因為其所帶正電荷數過高。口腔屬于動態環境,復雜多變,目前市面上漱口水作用時間一般為30 s,由此可以推測,當嵌合肽被應用于口腔產品時,作用時間減少,溶血率也會相應降低。
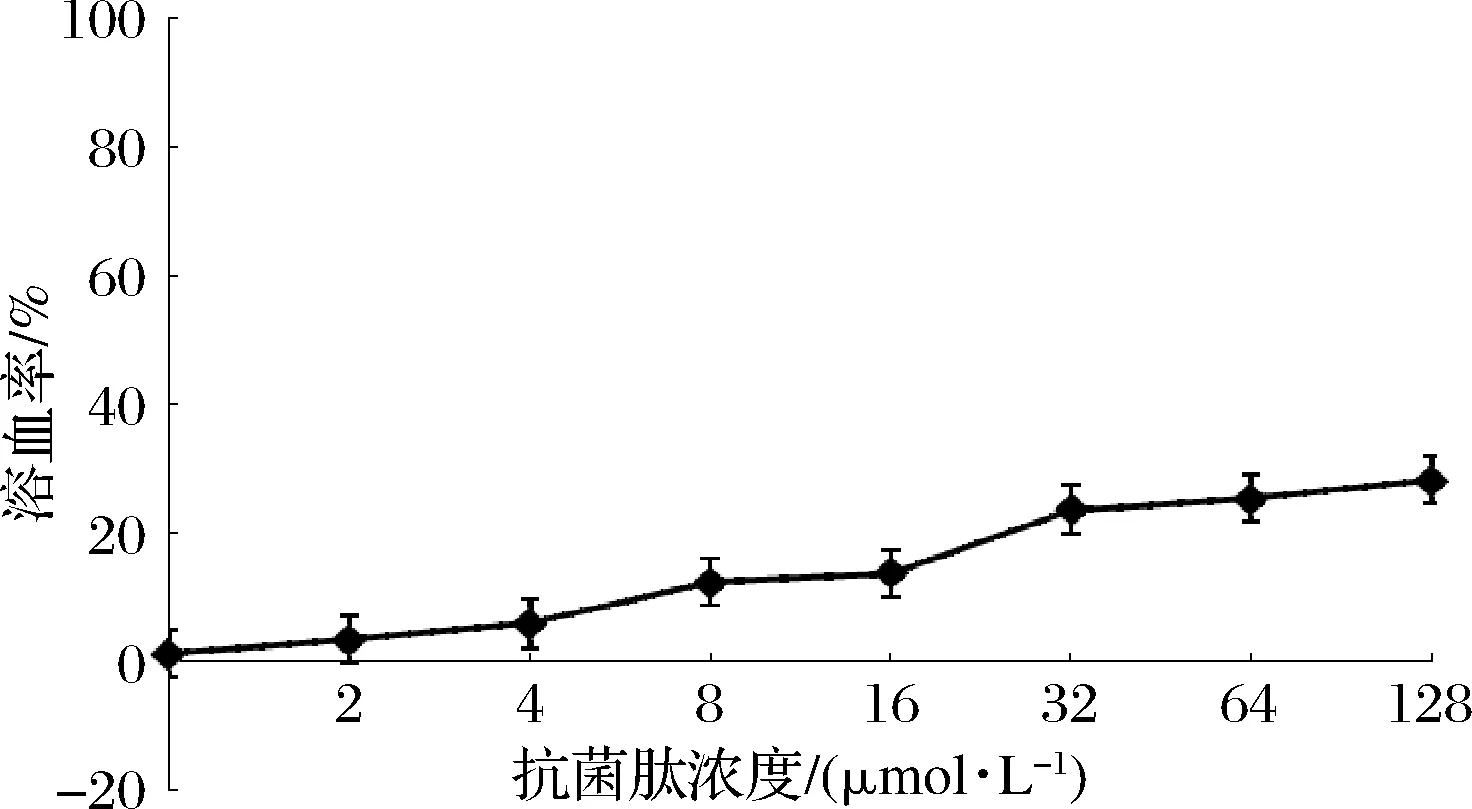
圖7 駝乳源LFA-LFC嵌合肽溶血率Fig.7 Hemolysis rate of LFA-LFC chimeric peptide
3 結論
本研究通過增加陽離子數,嵌入1個賴氨酸連接LFampin265-284和駝乳LFcin17-30獲得駝乳源LFA-LFC嵌合肽,對S.mutans、S.salivariu和S.sobrinus具有快速高效的抑制殺滅作用;可以穿過細胞膜進入細胞內部,與菌體DNA相互作用,破壞細胞的遺傳物質,阻礙細菌生長繁殖。這為優化抗菌肽結構,深入探究抗菌肽的作用機制,維護人體口腔環境健康提供新思路。
