二肽基肽酶4抑制劑在風濕免疫病中的潛在作用*
2023-02-14 14:01:00李新露黃劼董凌莉鐘繼新
醫藥導報 2023年2期
李新露,黃劼,董凌莉,鐘繼新,2
(華中科技大學同濟醫學院附屬同濟醫院1.風濕免疫科;2.過敏與臨床免疫研究所,武漢 430030)
二肽基肽酶4(dipeptidyl peptidase 4,DPP4),也稱為CD26,于1966年作為蛋白酶首次被發現[1],以同源二聚體或四聚體形式表達于細胞表面。DPP4的底物包括腸促胰島素肽、趨化因子、細胞因子以及神經肽。它通過從N端切割形如X-脯氨酸或X-丙氨酸(X為任意氨基酸)的二肽,改變其底物的生物學活性。DPP4可將胰高血糖素樣肽-1 (glucagon-like peptide-1,GLP-1)轉變為無活性形式,從而使其喪失結合GLP-1受體,并誘導胰腺β細胞釋放胰島素的功能。DPP4抑制劑能夠結合在由DPP4胞外區一些關鍵氨基酸殘基(E205、E206、R125、Y547、S630、Y662、Y666、D708、N710以及H740)組成的活性結構域,進而誘發構象改變,阻止DPP4與其底物如GLP-1的結合,最終起到控制血糖的作用。
自2006年以來,多種DPP4抑制劑已被批準用于治療2型糖尿病,可降低患者血糖水平。而近年來,DPP4被發現具有代謝調節之外的多種新功能,例如調節免疫細胞的活化及功能、促進纖維化和介導病毒進入宿主細胞等,并可被抑制劑所抑制。因此對DPP4抑制劑的免疫調節作用及其對風濕性疾病的潛在影響進行全面總結具有重要的指導意義。本綜述重點關注DPP4抑制劑在免疫調節方面的新證據,并總結了DPP4抑制劑對風濕性疾病的潛在影響。
1 DPP4的新功能
1.1DPP4的免疫調控作用
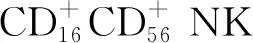
樹突狀細胞(dendritic cell,DC)上DPP4的表達最先在傳入淋巴結的部分亞群上檢測到。流式細胞術分析進一步表明,DPP4在經典樹突狀細胞上高表達。此外,與對照組比較, DPP4在肥胖小鼠和人類的內臟脂肪組織(visceral adipose tissue,VAT)中的DC上表達更高[3]。在體外DC分化過程中,檢測到DPP4表達明顯增加,這表明DPP4表達與DC細胞成熟有關。DPP4陽性DC有助于獲得性免疫,特別是Th1樣反應。具體的機制:一方面,在DC上表達的DPP4賦予其修飾巨噬細胞來源的趨化因子的能力,這可能趨化Th1細胞;另一方面,DC細胞上DPP4可以通過與腺苷脫氨酶(adenosine deaminase,ADA)結合調節VAT微環境中腺苷濃度,從而促進DC介導的T細胞的活化及分化[3]。
在高脂飲食誘導和遺傳性肥胖小鼠內臟脂肪組織的巨噬細胞上也檢測到DPP4表達,并且隨著巨噬細胞功能成熟,表達增加[3]。DPP4的長期抑制會下調脂肪組織巨噬細胞中的促炎基因,減少VAT中的炎癥,并防止單核細胞遷移和肌動蛋白聚合。同樣,通過DPP4抑制劑阿格列汀和shRNA沉默抑制DPP4會抑制巨噬細胞的浸潤和聚集。此外,與DC相似,巨噬細胞上DPP4的表達能夠通過調節微環境中的腺苷濃度來促進T細胞增殖[3]。
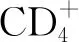
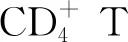
1.1.3DPP4調控趨化因子和細胞因子 如前所述,DPP4能夠通過切割N端二肽降解或改變多種細胞因子和趨化因子的活性。基質細胞衍生因子-1(stromal cell-derived factor-1,SDF-1),也稱為CXCL12,是T細胞、造血祖細胞和脂肪來源再生細胞的趨化因子。SDF-1可被DPP4進行蛋白水解裂解并轉化為CXCL12(3-68)。CXCL12(3-68)不能誘導CXCR4介導的β抑制蛋白的募集和IP3,Akt或ERK1/2的下游活化,從而失去其對淋巴細胞的趨化特性。DPP4抑制劑西他列汀顯著增強全長SDF-1[而不是CXCL12(3-68)]誘導關節內淋巴細胞浸潤的能力[6]。除了滅活SDF-1外,DPP4還通過與其受體CXCR4的直接相互作用調節SDF-1介導的淋巴細胞遷移。DPP4在T淋巴細胞上與CXCR4結合,SDF-1能夠誘導CXCR4/DPP4復合物的內化。內化的CXCR4可迅速回收到細胞膜,而DPP4則留在細胞內囊泡中,這表明SDF-1在減少DPP4依賴性失活方面具有自調節機制。其他證實可被DPP4酶解的趨化因子包括IP10、MIP、MIG、I-TAC、MDC、RANTES等。
DPP4還可降解多種細胞因子,包括成纖維細胞生長因子2(FGF2)、IL-3、粒細胞-巨噬細胞集落刺激因子(GM-CSF)、粒細胞集落刺激因子(G-CSF)、IL-3和促紅細胞生成素(EPO)。許多IL家族成員,如IL-2/-5/-10/-13/-17/-22/-23/-27/-28,也具有DPP4的潛在酶解位點。但是,這些位點是否為DPP4的真正截斷位點,仍需進一步的生化和生物學研究證實。
1.2DPP4促進纖維化 成纖維細胞是紡錘形細胞,負責結締組織中細胞外基質和膠原蛋白的合成。成纖維細胞不是傳統的免疫細胞,但它們在免疫調節和風濕疾病中起著重要作用。成纖維細胞也可以作為抗原提呈細胞,通過直接的細胞間相互作用和間接的細胞因子分泌促進適應性免疫細胞的增殖、激活和募集。
與小鼠胸腺髓質成纖維細胞比較,DPP4在被膜成纖維細胞中的表達水平更高,這表明DPP4可能作為區分不同成纖維細胞亞群的標記物。人皮膚中的主要成纖維細胞群被定義為SFRP2/DPP4。DPP4的表達可分離出纖維化疤痕形成譜系,并且抑制DPP4可減少傷口愈合過程中的皮膚瘢痕[7]。另一項研究表明,與健康人比較,系統性硬化癥患者的DPP4在皮膚成纖維細胞中的表達上調。DPP4在皮膚纖維化中調節TGF-β誘導的成纖維細胞活化,并表征以膠原和肌成纖維細胞標志物表達為特征的活化成纖維細胞群。DPP4的遺傳缺失或抑制可抑制成纖維細胞的增殖、遷移和膠原蛋白的產生[8]。LEE等[9]報道,DPP4可通過PAR2和下游NF-κB/SMAD信號轉導激活皮膚成纖維細胞。研究發現DPP4在葡萄糖酸氯己定誘導的大鼠腹膜纖維化模型增厚腹膜中表達增加。DPP4在糖尿病中的表達增加可促進上皮-間充質轉變和腹膜纖維化,且這些作用可以被DPP4抑制劑所抑制[10]。此外,DPP4與細胞外基質蛋白(如膠原和纖連蛋白)和細胞外基質降解酶(如纖溶酶原和鏈球激酶)的結合可能與細胞擴散和轉移相關。鏈激酶,纖溶酶原及其代謝物纖溶蛋白與DPP4富含半胱氨酸的區域結合,導致細胞內鈣離子(Ca2+)反應的快速增加以及隨后的成纖維細胞活化[11]。DPP4與纖溶酶原的結合還通過促進基質金屬蛋白酶的分泌和纖溶酶原向纖溶酶的轉化來調節細胞外基質的穩態。此外,纖溶酶的產生能夠降解BP180,BP180是一種在自身免疫性皮膚病大皰性類天皰瘡中發現的自身抗原。因此,DPP4可能參與維持BP180免疫耐受性和預防BP自身抗體的產生[12]。此外,有研究認為DPP4抑制劑可能具有抑制新型冠狀病毒感染(COVID-19)患者炎癥反應,減輕 COVID-19患者肺部損傷和纖維化的作用,然而尚需要臨床試驗研究加以證實[13]。
2 DPP4抑制劑在風濕免疫病中的潛在作用
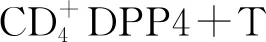
DPP4抑制劑西他列汀和維格列汀在博來霉素誘導的皮膚纖維化的小鼠模型中顯示出顯著的抗纖維化作用,可通過抑制TGF-β誘導的ERK信號通路改善皮膚厚度、羥脯氨酸含量和肌成纖維細胞累積[8]。維格列汀還有效減輕博來霉素誘導的肺纖維化中的纖維化和炎癥[15]。肌成纖維細胞是組織纖維化中產生膠原蛋白的主要細胞。在SSc患者皮膚中,肌成纖維細胞會導致組織緊張和皮膚/關節攣縮。而DPP4對于成纖維細胞的激活至關重要,DPP4陽性成纖維細胞高表達促纖維化基因,如:Ⅰ型膠原、Ⅲ型膠原、纖連蛋白等。此外,重組人DPP4刺激可促進肺成纖維細胞中纖連蛋白的產生,表明DPP4在成纖維細胞活化和組織重塑中發揮作用。最近的一項研究發現,表達DPP4的成纖維細胞參與皮膚瘢痕中的膠原沉積,抑制DPP4可以減少小鼠傷口愈合模型中瘢痕的形成[14]。后續研究表明,SFRP2/DPP4成纖維細胞亞群是SSc皮膚中促纖維化成纖維細胞的祖細胞,DPP4通過PAR2激活NF-κB和SMAD信號轉導,導致真皮成纖維細胞的激活[9]。因此,DPP4抑制劑可能通過調控成纖維細胞功能而參與SSc。盡管動物研究證明DPP4抑制劑在SSc中的前瞻性應用,但DPP4抑制劑在SSc患者中作用的臨床試驗研究有限,其在SSc中的潛在作用仍有待大樣本臨床研究進一步證實。
2.2DPP4抑制劑與系統性紅斑狼瘡(systemic lupus erythematosus,SLE) SLE是一種以典型皮膚紅斑表現為特征的慢性多系統自身免疫性疾病。最近一項研究發現,與健康小鼠比較,患有紅斑狼瘡樣綜合征的小鼠,血清DPP4活性顯著降低,表明DPP4可能參與SLE的發病機制。同樣,多個臨床證據表明,與健康對照組比較,SLE患者的血清和外周血單個核細胞中DPP4表達水平較低[16]。最近來自中國臺灣地區和韓國的人群隊列研究表明,DPP4抑制劑與自身免疫病患病風險降低有關,但分層分析未發現DPP4抑制劑使用是否與SLE發病風險相關[17]。然而,由于這些隊列中包含的SLE病例數較為有限,DPP4抑制劑與SLE的潛在關聯仍需進一步研究證實。
2.3DPP4抑制劑導致類風濕關節炎(rheumatoid arthritis,RA)發病風險增加 RA是一種以多關節疼痛、腫脹、僵硬和功能喪失為特征的慢性進行性自身免疫性疾病。雖然其確切的原因和發病機制尚不清楚,但人們普遍認為T細胞活化在RA炎癥的發生和維持中起重要作用。參與T細胞活化的各種因素在RA患者中表達增加,包括CD28、CD40、CTLA4、ILRA / IL2、IL-21等。而作為T細胞的活化標志物,DPP4在RA中的作用自然受到越來越多的關注。
如前所述,SDF-1是DPP4的底物,可通過與其受體CXCR4的相互作用在炎癥細胞募集中起核心作用。在抗原誘發關節炎的小鼠模型中,DPP4缺乏導致血清活性SDF-1的持續存在和關節炎關節中CXCR4陽性炎性細胞的浸潤增加。此模型血漿DPP4水平低于骨關節炎,與炎癥標志物C反應蛋白呈負相關。SDF-1的水平與RA患者滑液中DPP4+ T細胞的數量呈負相關。由于滑膜中SDF-1水平與疾病活動評分(DAS28)和炎癥標志物(血清C反應蛋白和IL-6)密切相關[21],因此RA中滑膜DPP4的降低可能導致SDF-1/CXCR4軸的滑膜炎癥。這些結果表明DPP4/SDF1/CXCR4在RA滑膜炎癥中起著關鍵作用。
滑膜細胞過度增生、破骨細胞過度活化導致的骨侵蝕是RA進展的嚴重后果。最近的一項研究表明,滑膜成纖維細胞中外源性DPP4或DPP4的過表達會減少成纖維細胞中促炎細胞因子(如IL-1β、IL-6和IL-3)的產生[18]。而在最近的一項研究中發現,DPP4在破骨細胞中高表達,骨吸收抑制劑地諾單抗可在抑制破骨細胞的同時,降低循環DPP4水平和增加GLP-1水平,顯著降低HbA1c水平,這表明破骨細胞來源的DPP4可能是糖代謝與骨重塑之間的重要聯系[22]。在小鼠RA模型中研究表明,DPP4的抑制增強滑膜成纖維細胞侵入軟骨。然而,在鏈脲霉素誘導的大鼠糖尿病模型中監測其骨骼效應后發現,DPP4抑制劑可能通過減少CTX-I依賴性骨吸收,從而減輕骨質流失并改善機械骨強度[23]。
對使用DPP4抑制劑患者的臨床觀察可能為DPP4酶活性在RA中的作用提供關鍵信息。已有多個群體報告DPP4抑制劑相關的新發RA病例[24]。然而,與其他抗糖尿病治療比較,更大規模的人群研究未能確定RA與DPP4抑制劑使用之間的關聯[25]。未來需要更多的隨機對照試驗研究DPP4抑制劑對RA發展的確切影響。
2.4DPP4抑制劑與炎癥性腸病(inflammatory bowel disease,IBD) IBD是一種典型的自身免疫介導的消化系統疾病,包括克羅恩病(Crohn's disease,CD)和潰瘍性結腸炎(ulcerative colitis,UC),與風濕免疫病相似,都存在異常的免疫應答,且炎性腸病性關節炎亦屬于風濕性疾病。與健康對照組或其他疾病組患者比較,IBD患者血清DPP4水平和酶活性顯著降低,且降低程度與疾病活動度相關。此外,治療應答者的血清DPP4水平相對無反應者更高[26]。與緩解期患者相比,糞便中DPP4活性在活動性潰瘍性結腸炎患者中降低,但在克羅恩病患者中增加[27]。
已有研究發現,DPP4抑制劑在IBD動物模型中有保護作用。給予DPP4抑制劑維達列汀后,乙酸誘導的結腸炎模型大鼠的結腸縮短和體質量減輕得到改善[28]。同樣,阿格列汀治療也有助于恢復C57BL/6小鼠DSS誘導的結腸炎的腸黏膜損傷。然而,關于DPP4抑制劑對IBD影響的臨床證據有限且不一致。多個研究表明,DPP4抑制劑的使用與IBD患病風險增加有關[29]。而另一項真實世界調查納入895例接受DPP4抑制治療及747例其他抗糖尿病治療的患者,結果顯示,使用DPP4抑制劑與IBD風險增加無關[30]。還有研究認為,使用DPP4抑制劑可縮短IBD的病程[31]。
總而言之,雖然DPP4在IBD患者血清和糞便中活性降低,且DPP4抑制劑雖然在動物模型中顯示出確切的對IBD的保護作用,但在臨床應用中與IBD的關系并不確定,需要更多的臨床研究提供真實世界數據。
2.5DPP4抑制劑可能改善自身免疫性糖尿病 1型糖尿病(type 1 diabetes mellitus,T1DM),也稱為自身免疫性糖尿病,其特征為免疫介導的胰腺β細胞破壞和早期胰島素分泌不足,其本質是一種自免疫紊亂,與風濕免疫病發病機制存在很大的相似性,因此稍作贅述。有研究發現T1DM患者血清DPP4活性增加,并且升高與糖尿病的持續時間相關[32]。然而,DPP4抑制對T1DM患者降低HbA1c的作用在臨床試驗中并不一致。
2.6DPP4抑制劑與其他風濕性疾病 除上述疾病外,DPP4還可能參與多種風濕性疾病。一項病例對照研究發現,干燥綜合征(Sjogren's syndrome,SS)患者的唾液DPP4活性增加,且DPP4活性與MMP9水平呈正相關。一系列研究報告DPP4與軟骨細胞生理學有關,抑制DPP4可抑制細胞外基質的降解,這被認為有助于骨關節炎的改善[34]。此外,據報道,DPP4抑制劑還可以改善銀屑病[35],可能是通過抑制T細胞活化實現。
綜上所述,目前的研究表明,DPP4抑制劑的使用可能導致部分風濕免疫病的發病風險增加或部分風濕免疫病發病風險降低。DPP4抑制劑對其各個疾病應用效果總結見表1。

表1 DPP4抑制劑對風濕性疾病的影響
3 結論與展望
DPP4的廣泛表達限制其作為精確生物標志物的潛力,目前并不是風濕免疫病的特異性標志物。但DPP4的表達水平在不同風濕免疫病中有顯著改變,且與疾病嚴重程度相關,因此在判斷疾病狀態及疾病活動度乃至臨床治療方面具有重要意義。實驗和臨床研究表明,DPP4可能在風濕性疾病的發病機制中起多種作用,DPP4抑制劑會在不同疾病條件下產生不同的結果。一個可能的原因是DPP4作為一種兼職蛋白,具有多種功能,包括各種底物的酶降解以及多種蛋白質-蛋白質非酶學相互作用。此外,DPP4抑制劑在不同細胞群中也可能具有不同的功能。為了剖析DPP4抑制劑對風濕性疾病的確切作用,未來的工作可能集中在DPP4在不同類型細胞中的作用,DPP4表達的時空特征(特別是在疾病的不同階段),DPP4的未識別配體以及針對DPP4非酶學作用的研究中。總之,DPP4在不同疾病中的具體作用及機制仍有待闡明。DPP4抑制劑在不同的風濕性疾病可能具有不同的作用,其作為降糖藥物,應視患者風濕病種類或易感性情況而作不同的考量。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
學苑創造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
汽車工程學報(2017年2期)2017-07-05 08:13:02
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
