外源鈣對蜜橘抗日灼生理病害的分子與生理機制研究
2023-02-21 08:04:54高志鍵李春俠周秋蓉李江波
西北植物學報 2023年1期
高志鍵,李春俠,周秋蓉,李江波,李 延,王 平*
(1 福建農林大學 園藝學院,園藝植物遺傳育種研究所,福州 350002;2 撫州市南豐柑桔研究所,江西撫州 344100;3 福建農林大學 資源與環境學院,福州 350002)
日灼病是由溫度過高和強直射太陽光所造成的一種生理病害,尤其是突發性的高溫與強光更容易導致日灼發生[1]。柑橘果實發生日灼后,果實畸形,灼傷部位果皮呈現黃色或深褐色,果肉呈現海綿狀,果汁變淡風味差,失去食用價值[2]。柑橘適宜種植在溫暖且潮濕的氣候下,若陽光長時間直射植株,使其表面溫度過高,會導致果實的灼傷,尤其是樹冠外圍結果較多的南豐蜜橘,其日灼果實發生率可達40%以上[3]。而樹冠內膛果實由于枝葉的遮擋,一般不會發生日灼。
在強光和高溫脅迫下,果實表皮的生理代謝水平會失衡,產生大量的ROS分子,導致膜損傷和結構改變,進而引起酚類物質的酶促和非酶促褐變,造成果皮日灼[4]。正常情況下,細胞內產生ROS和抗氧化酶活性升高是處于一種動態平衡,不會使植物受到傷害,但當植物處于逆境中,這種平衡被打破,植物體自身的保護防御機制也隨之不斷變化[5]。由此可見,抗氧化酶防御機制對植物本身非常重要。張蓓等[6]研究發現,噴施螯合鈣可以減緩蜜廣橘果皮ROS的積累,增強抗氧化酶的活性,保持了抗氧化系統的動態平衡。Ca2+作為胞內的第二信使,在植物對各種逆境信號的轉導中起著重要作用[7]。Ca2+信號通過與鈣調素(calmodulin,CaM)、依賴鈣激活的蛋白激酶(calcium-dependent protein kinases,CDPKs或CPKs)等相關靶蛋白結合而調節下游靶酶的活性,從而調節ROS的代謝[8],提高植物對逆境的適應性。前人研究發現CaM 拮抗劑處理可以明顯降低抗氧化酶超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX)等酶活性,說明抗氧化酶活性受到鈣信號轉導系統的調節[9-10]。已有研究表明,Ca2+參與多種逆境脅迫過程如低溫、鹽脅迫、氧脅迫等,施用外源鈣能提高植物的抗逆性[11-12]。而有關鈣信號轉導基因對蜜橘果實抗日灼作用的研究未見報道,為探討鈣信號系統對蜜橘抗氧化基因表達的影響,本研究以南豐蜜橘為試驗材料,研究鈣處理下不同發育階段鈣信號轉導途徑相關基因和抗氧化酶基因的變化以及果皮抗氧化酶活性和ROS含量的變化,以期為生產上用鈣處理預防蜜橘日灼生理病害問題提供理論支持。
1 材料和方法
1.1 材料與處理
試驗分別于2017和2018年的6月1日到11月10日在江西省撫州市南豐縣明誠果園進行,以中熟南豐蜜橘(CitrusreticulataBlanco cv.Nanfeng tangerine) 20年結果樹為試驗材料。選擇掛果量、樹勢相當,朝向一致無遮擋的果樹作為試驗樹,每個處理6株,常規管理。分別于6月1日和7月1日17:00后對不同處理試驗果樹的果實進行噴施處理,以果面滴水為度。2017年預試驗使用螯合鈣(美國布蘭特股份有限公司) 噴施濃度梯度為0(清水對照)、0.10、0.14、0.18和0.22 g·L-1;2018年選擇預試驗效果最好的螯合鈣濃度0.14 g·L-1噴施樹冠外圍頂部果實,以噴施清水的樹冠外圍頂部果實為對照。
7-11月的每月5日采樣1次,共計5次,每次每株在南和西南方向隨機采10個日灼果實和10個正常果實,每次每處理共采120個果實運至實驗室后,40個果實為一個重復,共3個生物學重復,日灼果取日灼處果皮,正常果取相同部位同等大小的果皮,稱取適當重量的果皮,分別混合均勻,用錫箔紙分裝,液氮速凍,隨后置于-80 ℃冰箱保存備用。
1.2 方 法
1.2.1 日灼果發生率統計高溫結束后(11月10日),分別統計鈣處理和對照的果實總數和日灼果數,計算果實日灼發生率。
果實日灼發生率(%)=日灼果數/果實總數×100%
1.2.2 果皮RNA提取及相關基因表達分析基因表達量測定:參照東盛生物通用RNA試劑盒進行RNA的提取,參照諾唯贊HiScriptⅡ Q RT SperMix for qPCR試劑盒合成cDNA。
基因表達分析采用實時熒光定量PCR方法,在羅氏熒光定量PCR分析儀上完成。本研究選擇了柑橘Actin基因作為內參基因,根據citrus HarvEST database(http://harvest.ucr.edu/)已知EST序列分別設計了內參基因Actin、CsNADK、CsMPK8、CsCPK等特異性引物(表1)。

表1 熒光定量PCR引物Table 1 Fluorescent quantitative PCR primers
基因擴增體系為:10 μL 2 × ChamQ Universal SYBR qPCR Master Mix,0.4 μL上/下游引物,2 μL cDNA,7.2 μL ddH2O。基因擴增體系體積20 μL。PCR反應程序:95 ℃預變性30 s;循環反應:95 ℃ 10 s,60 ℃ 30 s,45個循環;溶解曲線:95 ℃ 30 s,60 ℃ 60 s,95 ℃ 15 s。試驗重復3次,按照2-ΔΔCt法計算出基因的相對表達量。
1.2.3 果皮活性氧熒光染色果皮活性氧的分布:每月每個處理分別取6個果實,用冷凍切片機對日灼處果皮(2 mm×2 mm)進行切片,切片厚度為70μm,立即用氯甲基二氯二氫熒光素二乙酯(CM-H2DCFDA)對其染色,在黑暗環境下孵育20 min,放在載玻片上,用激光共聚焦顯微鏡進行觀察。

1.2.5 抗氧化酶活性測定酶液粗提取參照曹建康等[15]方法;SOD活性測定采用氮藍四唑光化還原法;CAT活性測定采用 H2O2紫外吸收法;POD活性測定采用愈創木酚法;APX活性的測定參考Yu等[16]的方法;谷胱甘肽還原酶(GR)活性測定參考Hasanuzzaman等[17]的方法;谷胱甘肽過氧化物酶(GPX)活性測定參考Yu等[18]方法。
1.3 數據分析
數據采用Microsoft excel 2019處理計算平均值和標準差,采用SPSS 19.0進行差異顯著性分析(P<0.05),利用Excel 2019軟件作圖。
2 結果與分析
2.1 鈣處理后南豐蜜橘果實日灼發生率的變化
在高溫結束后,對果實日灼發生率進行統計分析發現,鈣處理的果實日灼發生率顯著低于清水處理,日灼發生率降低了13.76%(圖1)。結果表明鈣處理能顯著降低南豐蜜橘果實的日灼發生率。

不同小寫字母表示處理與對照間在0.05水平差異顯著,下同圖1 鈣處理后南豐蜜橘果實日灼發生率變化Different normal letters indicate significant difference between treatment and control at P<0.05,the same as belowFig.1 Change of the incidence of sunburn fruits in Nanfeng tangerine after calcium treatment
2.2 鈣處理后南豐蜜橘果皮鈣信號轉導途徑相關基因的表達
圖2顯示了蜜橘果皮的鈣信號途徑相關基因相對表達量。從7月日灼病害發生至11月果實完熟,鈣處理的果皮CsNADK相對表達量總體顯著高于對照,在9月達到峰值;鈣處理的果皮CsMPK8的相對表達量在整體上顯著高于對照。從8月日灼病發生前期開始至11月果實完熟,鈣處理的果皮CsCPK相對表達量始終顯著高于對照。在最易發生日灼時期的7-10月,鈣處理降低了蜜橘果皮CsRBOH的相對表達量。8-10月份,鈣處理的果皮CsCAMK1的相對表達量均顯著高于對照,在10月達到峰值。鈣處理的果皮CsGAD的相對表達量從整體上顯著高于對照,其中在7和8月鈣處理的果皮CsGAD的相對表達量最高。說明鈣處理可以顯著提高蜜橘果皮鈣信號途徑相關基因CsNADK、CsMPK8、CsCPK、CsCAMK1和CsGAD的相對表達水平,并抑制了調控ROS產生的CsRBOH相對表達。

圖2 南豐蜜橘不同發育時期果皮鈣信號途徑相關基因的表達Fig.2 Expressions of calcium signaling pathway related genes in peels of Nanfeng tangerine during different development stages
2.3 鈣處理后南豐蜜橘果皮抗氧化酶相關基因的表達
如圖3所示,對照處理和鈣處理的果皮CsSOD(Cu-Zn)的相對表達量在高溫和強直射光脅迫的初期(7-8月)整體較高,而后期(9-11月)顯著降低,但鈣處理的果皮CsSOD(Cu-Zn)相對表達量在7-11月均顯著高于對照。在7-10月,鈣處理果皮CsCAT的相對表達量始終高于對照。對照處理和鈣處理的果皮CsPOD的相對表達量呈現出先下降后上升的趨勢,但鈣處理果皮CsPOD的相對表達量在整個果實發育過程中一直顯著高于對照,在8月達到峰值。在蜜橘果實發育期間,對照處理和鈣處理的果皮CsAPX的相對表達量呈現出先上升后下降的趨勢,但鈣處理果皮CsAPX的相對表達量始終顯著高于對照,并于8月達到峰值。在蜜橘發育過程中,對照處理和鈣處理的果皮CsGSR的表達趨勢與CsSOD(Cu-Zn)的相似,鈣處理的果皮CsGSR的相對表達量整體上顯著高于對照。從7-9月日灼病害發生最嚴重時期,鈣處理果皮CsGPX的相對表達量顯著高于對照,在8月達到峰值。以上結果說明鈣處理顯著提高了蜜橘果皮抗氧化酶相關基因的表達。

圖3 南豐蜜橘不同發育時期果皮抗氧化酶相關基因的表達Fig.3 Expression of antioxidant enzyme related genes in peels of Nanfeng tangerine during different development stages
2.4 鈣處理南豐蜜橘果皮ROS和丙二醛含量的變化


圖4 南豐蜜橘不同發育時期果皮過氧化物和MDA含量的變化Fig.4 Changes of peroxide and MDA contents in peels of Nanfeng tangerine during different development stages
2.5 鈣處理南豐蜜橘果皮ROS熒光強度分布的變化
由圖5可以看出,在果實遭受高溫脅迫初期(7月),對照處理和鈣處理的外果皮和白皮層開始出現綠色熒光;在8-9月高溫期,對照處理和鈣處理的果皮的ROS綠色熒光已經從外果皮漫延到中果皮,但鈣處理果皮ROS熒光分布擴散面積較小;10月,對照處理和鈣處理的外果皮ROS熒光繼續積累,鈣處理果皮白皮層綠色熒光分布面積逐漸減少;11月份對照處理和鈣處理的外果皮、中果皮細胞間隙急劇增大,白皮層細胞部分崩潰,ROS熒光信號部分大范圍消失。在蜜橘不同的發育時期,2個處理的ROS熒光強度均呈現先上升后下降的趨勢(圖5)。圖6表明,7-10月果實遭受高溫脅迫期,對照的外果皮ROS熒光強度最高,鈣處理的ROS熒光強度均顯著弱于對照,二者差異顯著。11月份果實進入成熟期,對照處理和鈣處理果皮熒光強度急劇下降(圖6)。說明果皮細胞在經過長時間ROS的毒害作用,產生ROS的細胞器(線粒體和葉綠體)嚴重損傷,生成ROS能力下降,導致對照處理和鈣處理的果皮ROS熒光強度低于其他月份。

圖5 南豐蜜橘不同發育時期果皮ROS的熒光分布Fig.5 Distribution of ROS in the peels of Nanfeng tangerine during different developmental stages

圖6 南豐蜜橘不同發育時期果皮ROS的熒光強度Fig.6 Fluorescence intensity of ROS in the peels of Nanfeng tangerine during different developmental stages
2.6 鈣處理南豐蜜橘果皮抗氧化酶活性變化
圖7顯示,在7月日灼病害開始發生到11月果實完熟,對照處理和鈣處理果皮中的抗氧化酶活性均呈現出先上升后下降的趨勢。說明在果實日灼病害發生初期,高溫和強直射光脅迫激發了抗氧化酶活性,病害發生后期,隨著ROS的積累,膜脂過氧化,使抗氧化酶的活性受到抑制。在不同的發育時期,鈣處理果皮的抗氧化酶活性均高于對照。在8月份鈣處理果皮的SOD、CAT、POD、APX、GR、GPX活性最高,與對照相比分別增加了35.82%、47.96%、31.48%、31.01%、47.65%和123.19%。隨著時間延長,鈣處理對抗氧化酶活性影響愈小。說明噴鈣能在一定程度上提高抗氧化酶的活性。

圖7 南豐蜜橘不同發育時期果皮抗氧化酶活性的變化Fig.7 Changes of antioxidant enzyme activities in peels of Nanfeng tangerine during different developmental stages
3 討 論
果皮日灼生理病害一直是蜜橘生產上的常見問題,尤其是近幾年全球氣溫的上升逐漸使日灼病害加重。目前生產上的防控措施主要是噴水降溫、果實套袋、遮陽網、噴石灰水和高嶺土以及果園生草等方式[2,19],但這些方法不同程度上存在低效、可操縱性差和成本高等問題。本研究中,高溫來臨前期在果實表面噴施螯合鈣,是一種簡便有效的預防蜜橘日灼病害的生產措施。
鈣是植物生長發育必需的大量營養元素之一,在植物細胞生理活動中,它不僅能維持細胞壁、細胞膜和膜結合蛋白的穩定性,還通過鈣信號轉導途徑相關基因參與調控許多重要的生理生化過程,從而影響植物對外界環境變化的響應與適應,以增強植物對逆境的抵抗能力[9,20]。本研究通過蜜橘果皮活性氧測定和活性氧的高質熒光染色發現,在7—10月高溫和強直射光脅迫下,果皮細胞產生了過量的ROS,從而產生過多的丙二醛,細胞膜脂過氧化,細胞結構破壞,細胞防御系統崩潰,導致了日灼果實的發生。前人研究表明,鈣信號轉導系統可以調節ROS的生成和清除,使細胞氧化-還原平衡,增加植物的抗逆性[8]。鈣信號可以通過CaM控制NAD激酶 (nicotinamide asenine dinucleotide kinase,NADK)的活性,調節ROS的產生[21]。水稻中AtNADK2的過表達提高了NADP/NAD比值,從而提高植物對氧化損傷的耐受性[22]。本研究結果表明,鈣處理能明顯刺激CsNADK基因,增加其表達量,增強植物對高溫和強直射光脅迫的抵抗力,這與前人研究結果一致。鈣信號轉導系統還可以通過調節植物呼吸爆發氧化酶(respiratory burst oxidase homologue,Rboh,又稱NADPH氧化酶)來間接影響ROS的產生,Rboh是參與植物ROS產生的關鍵酶,在調節NADPH氧化酶活性產生ROS中起到重要作用[23]。有絲分裂原激活的蛋白激酶(mitogen-activated protein kinase,MAPK)是真核生物中的信號分子,參與許多生物或非生物的脅迫反應的過程[24],MPK8是目前研究較多的MAPK激酶。前人發現擬南芥中 CAM 會激活 MPK8,被激活的MPK8 通過轉錄控制 RbohD 表達來負調控 ROS 積累,揭示了 CAM 介導的信號轉導在脅迫下微調 ROS 穩態的新機制[25]。另外CPKs的磷酸化作用也可以調節RBOH的活性[26],水稻中的OsCPK12通過誘導ROS清除基因OsAPX2/OsAPX8的上調,抑制OsRBOHI的表達,調節細胞內ROS的平衡,增加水稻的耐鹽性[27]。本研究發現,鈣處理顯著增加CSMPK8和CSCPK基因的表達,進而顯著降低CSRBOH的相對表達量,從而降低了ROS的積累,提高了蜜橘抗日灼的能力。
鈣/鈣調素依賴性蛋白激酶(calcium/calmodulin-dependent protein kinases,CaMKs)是真核生物鈣信號途徑中鈣調素下游的一類重要靶蛋白,能夠引起眾多代謝的關鍵酶或轉錄因子磷酸化,從而完成對細胞代謝活動或某些基因表達的調節[28]。在植物中發現CaMKs有廣泛的作用。如玉米的ZmCCaMK基因可以激活抗氧化防御[29]。在本研究中,鈣處理使CsCAMK1基因顯著上調,且在逆境中后期表達量最強,說明CsCAMK1可能參與了蜜橘抗日灼的過程。
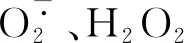
在強光和高溫脅迫下,鈣處理降低了南豐蜜橘果實的日灼發生率。本研究發現,鈣處理可促進蜜橘果皮細胞鈣信號轉導途徑相關基因的表達,并通過調控抗氧化系統相關基因表達以及抗氧化酶活性,清除細胞中過量的ROS,維持果皮細胞ROS代謝相對平衡,防止或降低了細胞膜脂過氧化,從而提高蜜橘果實對高溫和強光的耐受性。本研究證明了果面噴施螯合鈣防控日灼病害的可行性,為生產上預防和減輕蜜橘果實的日灼傷害提供了理論依據。
