鹽脅迫對甜櫻桃砧木生理特性及光合熒光參數的影響
2023-02-21 08:45:08李煥勇廖方舟劉景超王芝學楊麗芳
西北植物學報 2023年1期
關鍵詞:差異
李煥勇,廖方舟,劉景超,王芝學,楊麗芳
(天津市農業科學院 林業果樹研究所,天津 300384)
土壤鹽漬化已經成為全球重要的資源與環境問題,對農業生產和生態環境造成不同程度的影響,嚴重威脅著全球糧食安全。目前,全球土壤鹽漬化程度呈上升趨勢,中國各類鹽堿土總面積約為3.69×107hm2,占可用耕地面積的4.88%,主要分布在東北、西北、華北內陸以及沿海地區等,合理地開發利用鹽堿地資源,對中國農業生產以及生態環境可持續發展具有重要意義[1]。
鹽漬環境中的土壤水勢降低,會導致其中的植物吸收水分困難或水分外排,造成滲透脅迫[2]。但植物在鹽漬條件下可以通過有機滲透調節物質的代謝來降低自身水勢,進而減輕鹽漬環境對自身造成的損傷[1]。鹽脅迫還可造成植物膜脂過氧化現象,影響細胞膜通透性,進而妨礙植物生長發育和形態建成[3-4]。耐鹽植物在長期進化過程中形成了復雜且有效的抗氧化體系,主要包括以抗壞血酸(AsA)、類胡蘿卜素(Car)為主的非酶促抗氧化系統,以及超氧化物歧化酶(SOD)、過氧化氫酶(CAT)等為主的酶促抗氧化系統[5-7]。葉綠體作為植物對逆境最敏感的細胞器,在鹽脅迫條件下會受到一定程度的損傷,進而影響到植物的光合作用。鹽脅迫造成的滲透脅迫破壞了植物PSⅡ系統反應中心,降低了植物的光合電子傳遞效率和PSⅡ的光合作用活力[8-9]。
甜櫻桃(Cerasusavium)為薔薇科(Rosaceae)櫻桃屬(Cerasus)落葉果樹,在北方素有“春果第一枝”的美稱。甜櫻桃對鹽分敏感,通過嫁接可以提高其耐鹽性,選育優質抗逆砧木可以提高甜櫻桃的抗逆性。當前已對甜櫻桃砧木響應鹽脅迫的光合特性進行了一定研究[10-12],但對其在鹽脅迫下的滲透調節物質、抗氧化酶活性及葉綠素熒光特性等鮮見報道。本研究以栽培中常用甜櫻桃砧木‘吉塞拉6號’(Gisela 6)和‘考特’(Colt)為試驗材料,研究不同濃度NaCl處理對兩個甜櫻桃砧木葉片中主要滲透調節物質、抗氧化酶活性及光合熒光特性的影響,探討甜櫻桃砧木耐鹽生理機制,為甜櫻桃砧木耐鹽機制研究及優良砧木的開發利用提供理論依據。
1 材料和方法
1.1 試驗材料
供試材料為生長狀況一致且長勢良好的1年生‘吉塞拉6號’(G6)和‘考特’(KT)砧木苗,每個品種選擇30株定植于裝有混合基質(蛭石、珍珠巖和草炭土體積比1∶1∶3)的花盆中,在室外避雨棚下培養,苗木正常生長1個月后進行鹽脅迫處理。
1.2 材料處理
采用完全隨機區組設計,共設置0、50、100、150 mmol·L-1NaCl等4個濃度鹽脅迫處理水平,每個處理設置3次重復,每個重復10株幼苗。為防止鹽分流失,花盆底部放置塑料托盤,每次澆水后將流出的溶液及時倒回花盆內。鹽脅迫處理14 d后采集葉片樣品保存于液氮中用于生理指標檢測,并對植株進行光合氣體交換參數和葉綠素熒光參數的測定。
1.3 測定指標及方法
1.3.1 生理指標葉片可溶性糖含量測定采用蒽酮法,可溶性蛋白含量測定采用考馬斯亮藍染料結合法,脯氨酸含量測定采用茚三酮法,丙二醛含量測定采用硫代巴比妥酸法,超氧化物歧化酶(SOD)活性測定采用氮藍四唑光還原法,過氧化物酶(POD)活性的測定采用愈創木酚還原法[13-14]。
1.3.2 光合氣體交換參數和綠葉素熒光參數采用Li-6800便攜式光合-熒光測量儀(Li-COR,USA)進行葉片的光合氣體交換參數和葉綠素熒光參數的測定,測定控制光強度為1500 μmol·m-2·s-1,CO2濃度為400 μmol·mol-1,相對濕度為65%,每個處理測量5次。將所測幼苗葉片在夜間進行充分暗適應后,測定暗適應初始熒光(Fo)和最大熒光(Fm);白天充分光照后,測定凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)、胞間CO2濃度(Ci)等;同步執行飽和脈沖測量記錄光適應下初始熒光(Fo′)、光適應下最大熒光(Fm′)和穩態熒光(Fs),并計算光系統Ⅱ最大光化學效率(Fv/Fm)、光化學淬滅系數[qP=(Fm′-Fs)/(Fm′-Fo)]和非光化學淬滅系數[NPQ=(Fm-Fm′)/Fm′]。
1.4 數據處理
利用Microsoft Excel 2016對原始數據進行整理、標準差分析及圖形的繪制,利用SPSS 19.0軟件進行單因素方差分析,采用LSD法進行多重比較,方差分析在P<0.05時認為差異顯著。
2 結果與分析
2.1 鹽脅迫對兩個甜櫻桃砧木幼苗表觀性狀的影響
隨著NaCl處理脅迫濃度的升高,兩個甜櫻桃砧木‘吉塞拉6號’(G6)和‘考特’(KT)的生長狀況均逐漸變差,側枝減少,葉片表現出焦邊、卷曲、落葉等癥狀,其受鹽害程度逐漸加重,不同NaCl處理之間性狀差異明顯(圖1)。
2.2 鹽脅迫對兩個甜櫻桃砧木葉片滲透調節物質含量的影響
2.2.1 可溶性糖含量隨著NaCl處理濃度的升高,甜櫻桃砧木葉片中可溶性糖含量在‘吉塞拉6號’中呈現先增加后減少的趨勢,而在‘考特’中卻呈逐漸增加趨勢(圖2,A);在鹽脅迫條件下,‘吉塞拉6號’葉片中可溶性糖含量迅速增加并維持在較高水平,在100 mmol·L-1NaCl處理時達到最大值(77.98 mg·g-1),并與其他處理之間差異顯著(P<0.05);‘考特’葉片可溶性糖含量在150 mmol·L-1NaCl處理達到最大(72.92 mg·g-1),也與其他處理存在顯著差異。
2.2.2 可溶性蛋白含量圖2,B顯示,‘吉塞拉6號’和‘考特’葉片中可溶性蛋白的含量均隨著NaCl處理濃度的升高而逐漸增加,在150 mmol·L-1NaCl處理時達到最大,分別達到2.93 mg·g-1和3.06 mg·g-1,分別比對照顯著增加了51.7%和19.1%,其余濃度NaCl處理也與對照之間均存在顯著差異(P<0.05)。隨著NaCl處理濃度的升高,‘吉塞拉6號’葉片可溶性蛋白的增加量明顯高于‘考特’。

A.吉塞拉6號(G6);B.考特(KT);0、50、100、150分別表示0、50、100、150 mmol·L-1NaCl處理圖1 鹽脅迫下兩個甜櫻桃砧木植株形態和葉片表型變化A.Gisela 6 (G6);B.Colt (KT);0,50,100 and 150 indicate 0,50,100,and 150 mmol·L-1NaCl treatment,respectivelyFig.1 Plant morphological changes and leaf phenotypic changes of two sweet cherry rootstocks under salt stress
2.2.3 脯氨酸含量由圖2,C可知,兩個甜櫻桃砧木葉片中脯氨酸含量也均隨著NaCl處理濃度的升高而逐漸增加,且各濃度鹽脅迫處理之間及其與對照之間均存在顯著性差異(P<0.05);在相同鹽脅迫濃度下,‘考特’葉片的脯氨酸含量均不同程度高于‘吉塞拉6號’,它比對照的增加幅度也大于‘吉塞拉6號’。其中,在150 mmol·L-1NaCl處理濃度下,‘吉塞拉6號’和‘考特’葉片的脯氨酸含量與對照相比分別增加了2.14倍和2.87倍。
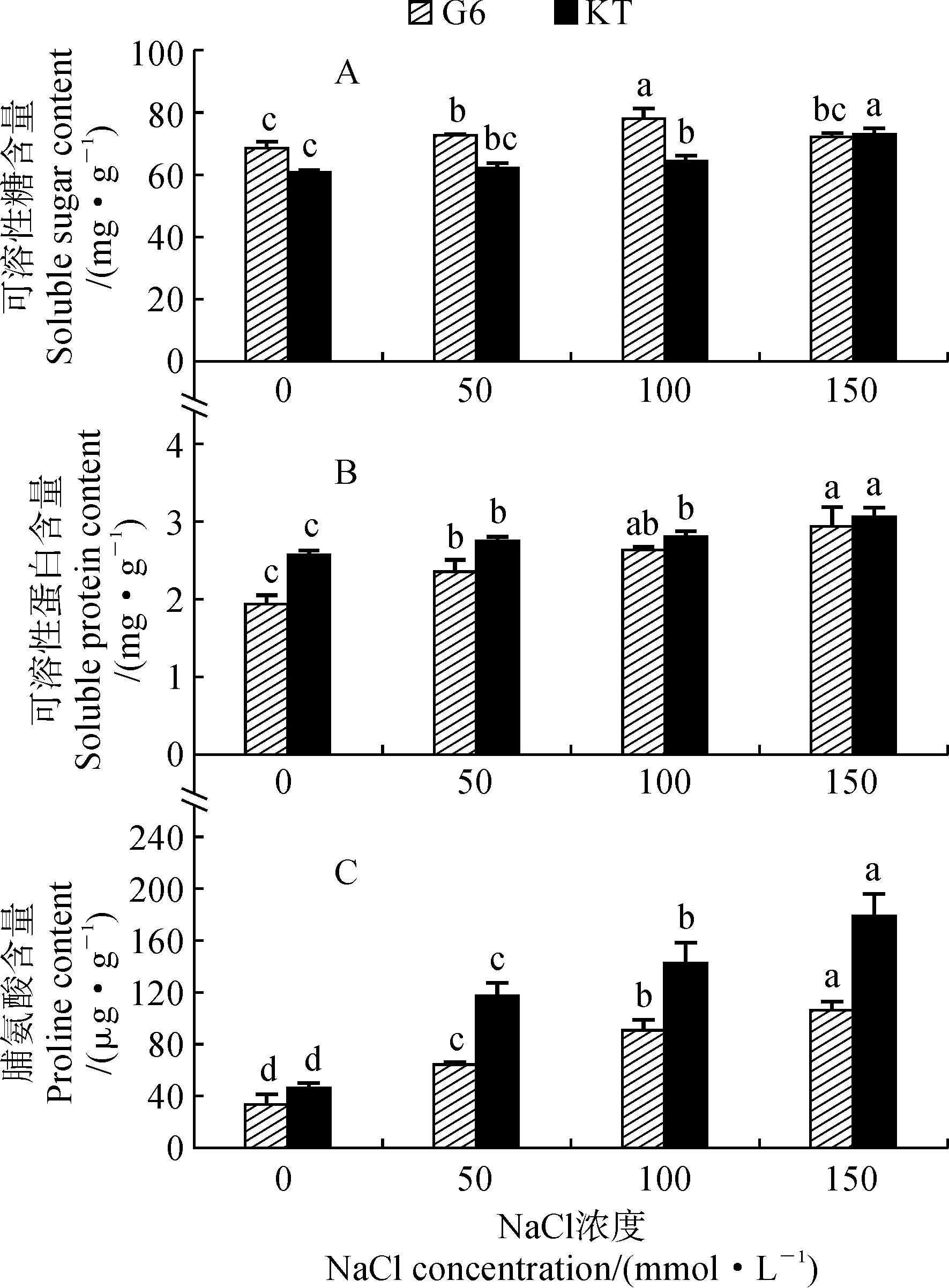
G6.吉塞拉6號;KT.考特;不同小寫字母表示同一品種不同處理間差異顯著(P<0.05),下同圖2 鹽脅迫下兩個甜櫻桃砧木葉片滲透調節物質含量的變化G6.Gisela 6;KT.Colt;Different normal letters indicate significant differences among different treatments of the same variety at the level of 0.05 (P<0.05);the same as below Fig.2 The contents of osmotic adjustment substance in leaves of two sweet cherry rootstocks under salt stress
2.3 鹽脅迫對兩個甜櫻桃砧木葉片MDA含量及SOD和POD活性的影響
2.3.1 丙二醛含量由圖3,A可知,兩個甜櫻桃砧木葉片丙二醛含量均隨著NaCl處理濃度的升高而增加,且在不同濃度NaCl處理間存在顯著差異;與對照相比,‘吉塞拉6號’和‘考特’葉片丙二醛含量分別在50和100 mmol·L-1濃度開始存在顯著差異(P<0.05),在150 mmol·L-1NaCl處理時分別顯著增加了26.3%和15.4%。在相同濃度NaCl處理下,‘吉塞拉6號’葉片丙二醛含量均明顯高于相應‘考特’,且比對照增幅也高于‘考特’。
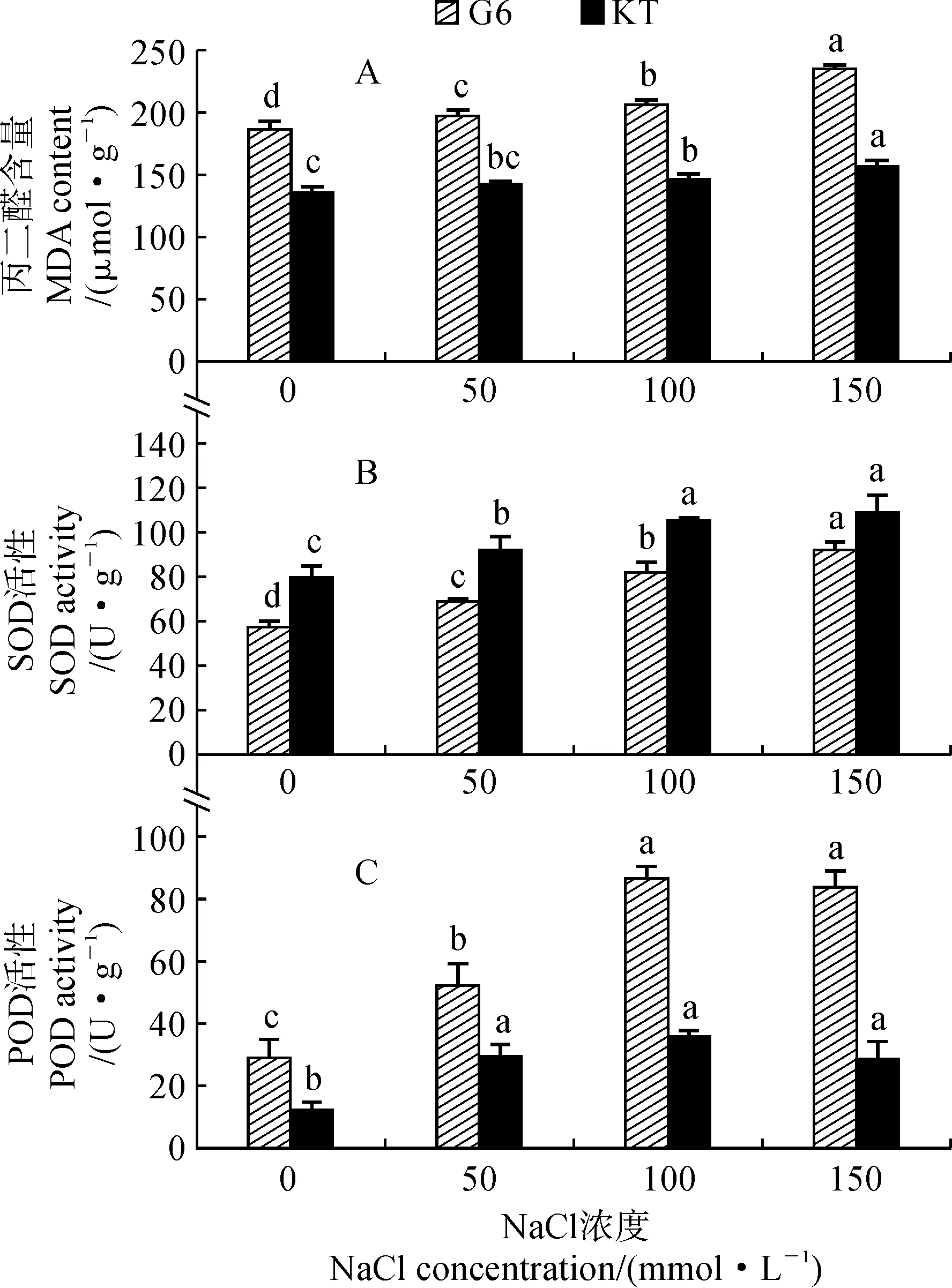
圖3 鹽脅迫下兩個甜櫻桃砧木葉片丙二醛含量及SOD和POD活性的變化Fig.3 The MDA content and SOD and POD activities in leaves of two sweet cherry rootstocks under salt stress
2.3.2 SOD活性兩個甜櫻桃砧木葉片中SOD活性隨著NaCl處理濃度的增加均呈現持續增加的變化趨勢,在150 mmol·L-1NaCl處理時分別達到92.11和109.01 U·g-1,分別比對照顯著增加了60.5%和36.3%,‘吉塞拉6號’的增加幅度明顯大于‘考特’(圖3,B)。‘吉塞拉6號’葉片中SOD活性在不同濃度處理之間均存在顯著性差異(P<0.05),而‘考特’葉片中SOD活性在50 mmol·L-1NaCl處理時與對照相比無顯著變化,在100和150 mmol·L-1NaCl處理時顯著高于對照和50 mmol·L-1NaCl處理,但它們之間沒有顯著性差異。
2.3.3 POD活性隨著NaCl處理濃度的增加,兩個甜櫻桃砧木葉片中POD活性均呈現出先升高后下降的變化趨勢,并均顯著高于相應對照,且均在100 mmol·L-1NaCl處理時達到最大值,此時分別比對照增加了2倍和1.97倍(圖3,C)。在相同處理條件下,‘吉塞拉6號’葉片的POD活性均明顯高于‘考特’;‘吉塞拉6號’葉片的POD活性表現為100和150 mmol·L-1濃度NaCl處理均顯著高于50 mmol·L-1NaCl處理(P<0.05),而‘考特’在3個濃度處理之間不存在顯著差異。
2.4 鹽脅迫對兩個甜櫻桃砧木葉片光合氣體交換參數的影響
2.4.1 凈光合速率兩個甜櫻桃砧木葉片凈光合速率(Pn)隨著NaCl處理濃度的升高而逐漸降低,且各鹽脅迫處理均與對照差異顯著;在150 mmol·L-1NaCl處理時,‘吉塞拉6號’和‘考特’的Pn與對照相比分別降低了85.7%和74.2%,‘吉塞拉6號’的下降幅度顯著大于‘考特’(圖4,A)。在相同處理條件下,‘吉塞拉6號’的Pn均明顯高于相應處理的‘考特’。‘吉塞拉6號’的Pn在各處理之間均存在顯著性差異(P<0.05),而‘考特’的Pn在100與150 mmol·L-1NaCl處理之間差異不顯著。
2.4.2 氣孔導度隨著NaCl處理濃度的增加,兩個甜櫻桃砧木葉片的氣孔導度(Gs)變化趨勢與Pn趨勢相似。兩個甜櫻桃砧木葉片的Gs在不同濃度NaCl處理下均明顯低于對照;‘吉塞拉6號’和‘考特’的Gs在150 mmol·L-1NaCl處理下分別比對照顯著降低了41.7%和63.7%,‘考特’的Gs下降幅度顯著大于‘吉塞拉6號’,且‘考特’的Gs在各處理條件下均低于相應的‘吉塞拉6號’(圖4,B)。‘考特’的Gs在各鹽濃度處理下與對照相比均存在顯著性差異(P<0.05),而‘吉塞拉6號’的Gs僅在100和150 mmol·L-1NaCl處理時與對照有顯著性差異。
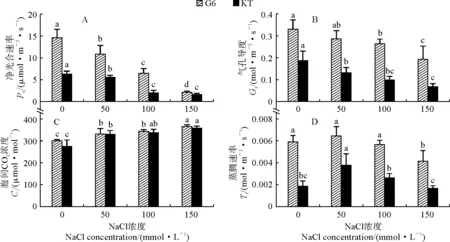
圖4 鹽脅迫下兩個甜櫻桃砧木葉片光合氣體交換參數的變化Fig.4 The photosynthetic gas exchange parameters in leaves of two sweet cherry rootstocks under salt stress
2.4.3 胞間CO2濃度圖4,C顯示,隨著NaCl處理濃度的增加,兩個甜櫻桃砧木葉片的胞間CO2濃度(Ci)表現出逐漸升高的趨勢,‘吉塞拉6號’和‘考特’在150 mmol·L-1NaCl處理下分別比對照顯著增加了21.7%和30.6%;兩個砧木葉片的Ci在鹽脅迫處理下增加幅度均不大,但各處理與對照之間均存在顯著性差異(P<0.05)。
2.4.4 蒸騰速率由圖4,D可知,兩個甜櫻桃砧木葉片的蒸騰速率(Tr)均隨著NaCl處理濃度的增加呈現出先升高后降低的變化趨勢,并均在50 mmol·L-1NaCl處理時達到最大值,期間‘吉塞拉6號’始終明顯高于相同條件下的‘考特’;與對照相比,僅‘吉塞拉6號’的150 mmol·L-1NaCl處理、‘考特’的50 mmol·L-1NaCl處理有顯著差異,兩品種其他鹽脅迫處理與對照之間均不存在顯著性差異。
2.5 鹽脅迫對兩個甜櫻桃砧木葉片葉綠素熒光參數的影響
首先,初始熒光(Fo)表示的是光系統Ⅱ(PSⅡ)反應中心處于完全開放時的熒光水平。由圖5,A可知,兩個甜櫻桃砧木葉片的Fo隨著NaCl處理濃度的升高整體均呈現上升的趨勢,但‘吉塞拉6號’僅在150 mmol·L-1NaCl時比對照增加顯著,‘考特’在100和150 mmol·L-1NaCl時增加顯著;與對照相比,‘吉塞拉6號’和‘考特’的Fo在150 mmol·L-1NaCl時分別顯著增加29.0%和68.7%,‘考特’增幅明顯大于‘吉塞拉6號’。兩個甜櫻桃砧木的Fo在對照處理和50 mmol·L-1NaCl處理時表現為‘吉塞拉6號’高于‘考特’,而在其余處理下則表現相反。最大熒光(Fm)反映了PSⅡ反應中心處于完全關閉時的熒光水平。兩個甜櫻桃砧木葉片的Fm隨著NaCl處理濃度的增加呈現不規律的變化趨勢,但各濃度鹽處理之間及其與對照之間均無顯著差異(圖5,B)。
其次,Fv/Fm反映了暗適應下植物PS Ⅱ 潛在最大光化學效率。隨著NaCl處理濃度的升高,‘吉塞拉6號’和‘考特’葉片的Fv/Fm基本上均呈現降低的趨勢,但均僅在150 mmol·L-1NaCl處理時與對照達到顯著差異水平(P<0.05),降幅分別為9.0%和15.0%,且‘吉塞拉6號’高于‘考特’(圖5,C)。
再次,非光化學淬滅系數(NPQ)主要反映天線色素所吸收光能中以熱能形式耗散掉的能量部分,而光化學淬滅系數(qP)反映了天線色素捕獲的光能中用于光化學反應的能量部分。隨著NaCl處理濃度的升高,兩個甜櫻桃砧木的NPQ呈現逐漸升高的趨勢,而qP呈現逐漸降低的趨勢;與對照相比,‘吉塞拉6號’的NPQ和qP在各濃度鹽脅迫下均差異顯著,而‘考特’則分別僅在100~150 mmol·L-1和150 mmol·L-1NaCl處理時差異顯著;在150 mmol·L-1NaCl處理時,‘吉塞拉6號’和‘考特’的NPQ與對照相比分別顯著增加了73.2%和117.9%,而兩品種的qP則比對照分別顯著減少了61.1%和62.0%,即‘考特’的NPQ變化幅度明顯大于‘吉塞拉6號’(圖5,D、E)。
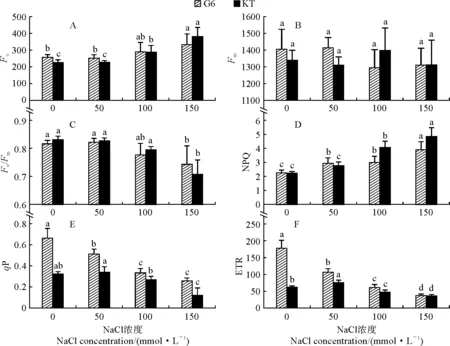
圖5 鹽脅迫下兩個甜櫻桃砧木葉片葉綠素熒光參數的變化Fig.5 The chlorophyll fluorescence parameters in leaves of two sweet cherry rootstocks under salt stress
另外,光合電子傳遞效率(ETR)主要反映了實際光強條件下的表觀電子傳遞效率。圖5,F顯示,隨著NaCl處理濃度的升高,‘吉塞拉6號’的ETR呈現逐漸降低的趨勢,且在各處理及對照間均存在顯著差異,其在150 mmol·L-1NaCl時比對照顯著減少了79.1%(P<0.05);而‘考特’的ETR隨著NaCl處理濃度的升高呈現出先升高后降低的變化趨勢,且在各處理及對照間均存在顯著差異,其在50 mmol·L-1NaCl時最大并顯著高于對照,而在其余濃度鹽脅迫下顯著低于對照(P<0.05)。
可見,隨著NaCl處理濃度的升高,兩個甜櫻桃砧木葉片的Fo、NPQ均呈現上升的變化趨勢,而Fv/Fm、qP和ETR則表現出逐漸降低的變化趨勢。
3 討 論
植物在鹽漬條件下可以通過調整體內各種新陳代謝途徑,控制各種有機代謝產物的合成和降解及氧化酶活性等來維持細胞內水勢平衡,進而可以調控自身生產及滲透調節。在鹽脅迫條件下,植物體內可溶性糖、可溶性蛋白及脯氨酸等物質參與滲透調節維持滲透平衡[1]。可溶性糖在鹽脅迫條件下還可作為能量物質,并對維持膜的穩定具有重要作用[15-16]。研究表明,鹽脅迫條件下小果白刺[1]、流蘇[17]、耐鹽花生[18]幼苗中可溶性糖含量均逐漸增加。本研究也發現,NaCl處理促進了兩個甜櫻桃砧木葉片中可溶性糖含量的積累,而‘吉塞拉6號’在高濃度NaCl處理時出現下降趨勢,可能是高鹽脅迫下能量消耗加劇所致,這與胡愛雙等對八棱海棠的研究結果相似[19]。可溶性蛋白作為一種親水性物質,鹽脅迫促進可溶性蛋白的積累可以增加水分的束縛,降低細胞滲透勢[20]。本研究中隨著NaCl處理濃度的升高兩個甜櫻桃砧木葉片中可溶性蛋白含量均逐漸增加,可能是由于鹽脅迫促進了甜櫻桃砧木的蛋白質合成代謝,造成可溶性蛋白的逐漸積累,從而降低細胞滲透勢來適應鹽脅迫環境。這與張婭等對小麥的耐鹽性研究結果相一致[21]。鹽脅迫下植物體內脯氨酸含量的增加可以調節體內滲透勢,防止因鹽脅迫造成過度失水而對細胞造成傷害。本研究發現,兩個甜櫻桃砧木葉片中脯氨酸含量隨著NaCl處理濃度的升高而呈現出逐漸增加趨勢,脯氨酸的積累量與NaCl處理濃度之間呈現正相關的關系,這與小果白刺[1]及八棱海棠等[19]研究結果一致,這是由于在鹽脅迫下甜櫻桃砧木體內通過脯氨酸積累來調節體內微環境,從而可增強對鹽脅迫的適應能力。
同時,在鹽脅迫條件下,植物體內活性氧積累,會對植物造成氧化損傷[22],而植物體內SOD、POD等抗氧化酶可以通過清除活性氧來減少膜系統的傷害,降低其對植物體造成的損傷[23]。丙二醛作為膜脂過氧化的產物之一,其含量的多少反映了植物對鹽脅迫抵抗能力的強弱[24]。本研究表明,隨著NaCl處理濃度的升高,甜櫻桃砧木葉片中丙二醛含量逐漸增加,這是由于隨著脅迫濃度的加強,甜櫻桃砧木發生膜脂過氧化作用,破壞了膜結構的完整性,從而導致丙二醛含量增加。這與鹽脅迫下葡萄、榆樹等丙二醛含量變化趨勢相一致[25-26]。同時,甜櫻桃砧木葉片中SOD活性隨著處理濃度的升高而增加,這說明在鹽脅迫下甜櫻桃砧木的抗氧化能力增強,可以清除大量過剩的活性氧,降低活性氧對甜櫻桃砧木細胞膜的損傷;甜櫻桃砧木葉片中POD活性隨著NaCl處理濃度的升高而表現出先升高后降低的變化趨勢,且‘吉塞拉6號’的POD活性明顯高于‘考特’,這與對蘋果砧木及梅花的研究結果一致[27-28],這可能是由于‘吉塞拉6號’具有較強的抗鹽性,可以通過提高POD的活性來減輕鹽脅迫對植物的傷害,而鹽脅迫濃度過高時已超出了甜櫻桃砧木的耐受能力,其POD活性隨之降低,自身調節能力也同時降低。
光合作用為植物獲取能量的重要途徑,鹽脅迫對植物自身光合作用效率會產生顯著影響。許多研究發現,鹽脅迫條件下植物葉片Pn、Gs和Tr會發生不同程度的降低[29-30]。鹽脅迫影響植物葉片Pn下降的主要原因是受氣孔或非氣孔因素的限制,如果Pn和Ci均降低時,認為氣孔開度變小為Pn降低的主要原因,光合作用受氣孔限制因素影響;當Pn降低而Ci升高時,認為光合作用下降受非氣孔因素影響,是由于鹽脅迫破壞了光合機構,影響了電子傳遞速率[31]。本研究發現,鹽脅迫對兩個甜櫻桃砧木葉片的Pn、Gs、Ci和Tr均具有顯著影響,隨著NaCl處理濃度的升高,甜櫻桃砧木葉片的Pn、Gs逐漸降低,而Ci逐漸升高。據此認為,非氣孔限制可能是鹽脅迫下兩種甜櫻桃砧木葉片Pn下降的主要因素,可能是由于鹽脅迫導致甜櫻桃砧木葉片光合機構受損,影響了RuBP羧化酶效率降低和光合作用表觀量子效率[32]。已有研究發現,鹽脅迫降低了黃連木及葡萄砧木葉片的Gs[33-34],同時本研究中甜櫻桃砧木葉片Gs也受鹽脅迫影響而下降,這可能是由于鹽脅迫造成滲透脅迫,水分向地上部分運輸受阻,葉片保衛細胞失水導致氣孔逐漸閉合。
葉綠素熒光參數包含豐富的光合信息,可反映植物在逆境條件下光系統對光能的吸收、分配、傳遞和耗散等情況,逆境脅迫對光合作用的影響程度都可依據葉綠素熒光參數的變化來顯示,是檢測鹽脅迫對植物光合機構傷害程度的重要指標[35-38]。Fv/Fm能夠反映植物PSⅡ潛在最大光化學效率,表示植物葉片的光抑制程度。研究發現正常生長植物葉片Fv/Fm經過充分暗適應后比較恒定,一般處于0.80~0.85之間,當Fv/Fm<0.8時,PSⅡ反應中心可能由于脅迫造成一定損傷[37-40]。本研究發現,甜櫻桃砧木葉片的Fv/Fm經NaCl處理后總體呈現降低的變化趨勢,但在低濃度(≤50 mmol·L-1NaCl)處理時Fv/Fm值在恒定范圍內,在高濃度(≥100 mmol·L-1NaCl)處理時Fv/Fm<0.80,這表明低濃度NaCl處理對甜櫻桃葉片的PSⅡ反應機構影響不大,而在高濃度NaCl處理導致植物細胞結構受損,破壞了細胞的光合機構,電子傳遞速率和光化學量子效率遭到光抑制,降低了植株對光能的利用率,進一步證明了光合作用效率下降是受非氣孔因素影響,這與甜櫻桃、葡萄等相關研究結果一致[41-42]。隨著NaCl處理濃度的升高,甜櫻桃砧木葉片中NPQ表現出升高的變化趨勢,而qP和ETR均呈現下降的變化趨勢,說明鹽脅迫下甜櫻桃砧木通過降低PSⅡ原初光能轉換效率和抑制電子傳遞效率,以減少用于光化學電子傳遞的份額,減輕光合器官受到損傷程度;同時PSⅡ反應中心會啟動保護性調節機制,通過增加熱耗散來消耗過多激發能,從而實現自我保護[43],鹽脅迫下辣椒[34]、葡萄砧木[44]及八棱海棠等[45]均表現出相似的保護機制。
綜上所述,鹽脅迫條件下,甜櫻桃砧木通過增加葉片可溶性糖、可溶性蛋白及脯氨酸等滲透調節物質含量,緩解滲透脅迫對其造成的傷害;另外還通過提高SOD和POD活性,提高抗氧化能力,清除體內過剩活性氧,減輕鹽脅迫造成的氧化損傷。同時,鹽脅迫抑制了甜櫻桃砧木的光合作用,其主要受限制因素為非氣孔因素;鹽脅迫下甜櫻桃砧木葉片細胞光合機構受損,電子傳遞速率和光化學量子效率受到光抑制,降低了植株對光能的利用率,導致光合速率降低。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
