秋播冬麥區矮稈基因 RhtB1b、 RhtD1b和 Rht8的分布頻率及其與產量性狀的關系
2023-04-25 09:10:18蘭素缺金京京張穎君彭曉慧李杏普張業倫
麥類作物學報 2023年4期
關鍵詞:檢測
張 凱,蘭素缺,金京京,張穎君,彭曉慧,李杏普,張業倫
(河北省農林科學院糧油作物研究所/河北省作物遺傳育種實驗室,河北石家莊 050035)
利用矮稈基因調節株高是培育高產小麥品種的重要途徑[1],同時,矮稈基因還可以提高小麥的抗倒伏能力、抗旱性和產量[2-3]。小麥矮稈基因大致分為三類:D-草叢型矮生基因、Us -單莖矮生基因和Rht基因[4]。目前,已被發掘并命名的矮稈基因有24個。其中,被廣泛應用的矮稈基因主要是來自農林10號和赤小麥的Rht1(Rht-B1b)、Rht2(Rht-D1b)和Rht8、Rht9[5-7]。Rht-B1b、Rht-D1b和Rht8基因分別位于4BS、4DS和2DL染色體上[8]。Rht-B1b和Rht-D1b基因編碼的DELLA蛋白,主要抑制植物對赤霉素(GA)的響應,但該蛋白并不結合GA受體蛋白,因此對赤霉素表現為不敏感[9]。Rht8是赤霉素敏感基因,通過調節赤霉素合成相關基因的表達,影響不同活性GA的含量比例,從而降低株高[10]。 Ellis等[11]研究表明,Rht-B1b和Rht-D1b是中等強度降稈基因,兩個基因同時存在時,存在較強的累加效應,降稈效應可達40%以上,這兩個基因的降稈效應均強于Rht8基因。
矮稈基因的定位與克隆為分子標記的開發奠定了基礎。Peng等[9]利用同源克隆的方法獲得小麥矮稈基因Rht-B1a和Rht-D1a。在此基礎上,Ellis等[11]開發了STS標記BF/MR1、BF/WR1、DF1/MR2和DF2/WR2,分別用于檢測矮稈基因Rht-B1b、Rht-B1a、Rht-D1b和Rht-D1a。為了提高檢測效率,Rasheed等[12]和Wu等[13]分別開發了KASP和STARP標記,用于檢測Rht-B1a和Rht-D1a基因型。Korzun等[14]和Debora等[10]分別開發了Rht8基因連鎖的微衛星標記Xgwm261和側翼標記DG279、DG371,可用于檢測Rht8基因。熊宏春等[15]將Rht8基因精細定位于分子標記 INDEL-2005 和 SSR-2650 之間,并開發了分子標記CAPS-Rht8。分子標記的開發為分子標記輔助選擇育種以及矮稈基因的高效應用提供了便利的檢測方法。
目前,中國北部冬麥區大部分小麥品種株高為75~80 cm,南部冬麥區為85~95 cm,已接近最適株高[16]。然而,矮稈基因在不同麥區分布多樣化且無規律性,不同麥區間生態環境和栽培條件也差異較大,小麥品種的矮稈基因類型及產量性狀結構特點也不明確,給育種親本選配和矮稈基因的高效利用造成困擾。因此,本研究利用STS分子標記BF/MR1、BF/WR1、DF1/MR2和DF2/WR2以及微衛星標記Xgwm261,對不同生態區近十年來育成的237份秋播冬小麥品種進行Rht-B1b、Rht-D1b和Rht8基因檢測,并分析三個基因的分布情況以及與株高和產量相關性狀的關系,以期為育種親本選配提供參考依據。
1 材料與方法
1.1 試驗材料
供試材料共237份,其中黃淮冬麥區121份、北部冬麥區57份、西南冬麥區41份、長江中下游冬麥區17份、新疆冬春麥區1份,大部分為2010年后育成的品種。分別于2019-2020和2020-2021年度在河北省農林科學院糧油作物研究所堤上試驗站,按照隨機區組設計,每份材料種植5行,每行4 m,株距5 cm,行距25 cm。
1.2 基因組DNA的提取
每份材料選取兩粒具有代表性的種子,用鋼珠打碎后,參照TPS法[17]提取基因組DNA。用核酸定量儀(Nano Drop 2000)檢測DNA濃度,將DNA原液稀釋至100 ng·μL-1后,保存于 -20 ℃備用。
1.3 矮稈基因的分子標記檢測
利用Ellis 等[11]設計的四對STS分子標記BF/MR1、BF/WR1、DF1/MR2和DF2/WR2檢測Rht-B1和Rht-D1基因(表1),其中BF/MR1與BF/WR1為一對互補引物,分別用于檢測Rht-B1b和Rht-B1a;DF1/MR2與DF2/WR2也為一對互補引物,分別用于檢測Rht-D1b和Rht-D1a,以石4185(含有Rht-B1b基因)和鄭麥7698(含有Rht-D1b基因)為陽性對照。用Korzun等[10]設計的微衛星標記Xgwm261檢測Rht8基因,以石4185為陽性對照。PCR反應體系均為10 μL,包括模板DNA 2 μL,2×SanTaq PCR Mix 預混液5 μL,上、下游引物(濃度為0.025 nmol·μL-1)各0.3 μL,用滅菌雙蒸水補至 10 μL。PCR反應程序:94 ℃預變性5 min;94 ℃變性30 s,55~63 ℃退火30 s,72 ℃延伸30 s,35個循環;72 ℃延伸5 min,4 ℃保存。Rht-B1和Rht-D1的PCR擴增產物用1.5%的瓊脂糖凝膠(含Gold View染料)進行檢測,在凝膠成像儀下觀察并拍照記錄。Rht8基因的PCR擴增產物用8%的非變性聚丙烯酰胺凝膠進行檢測,銀染顯色,拍照記錄。

表1 本研究所用的分子標記信息
1.4 小麥株高和產量性狀調查
于2020和2021年小麥灌漿中后期,每份材料選取10個單株測量株高,同時隨機調查10個麥穗的穗長。于2020和2021年小麥成熟期,用萬深SC-G自動考種儀統計千粒重、粒長和粒寬;人工調查單位面積穗數,收獲后稱重測產;隨機選取10個麥穗,調查穗粒數。所有測量指標均取2年的平均值作為最終調查結果。
2 結果與分析
2.1 Rht-B1b、 Rht-D1b和 Rht8基因的分子標記檢測結果及其分布頻率
檢測結果表明,發現237份供試材料中,僅含Rht-B1b(突變型)的材料有17份,占比7.2%;含Rht-B1b并同時含其他矮稈基因的材料有22份,占比9.3%(圖1和表2);僅含Rht-D1b(突變型)的材料有63份,占比26.6%;含Rht-D1b并同時含其他矮稈基因的材料有50份,占比21.1%(圖2和表2);僅含Rht8的材料有39份,含Rht8并同時含其他矮稈基因的材料有65份,分別占 16.5%和27.4%(圖3和表2)。
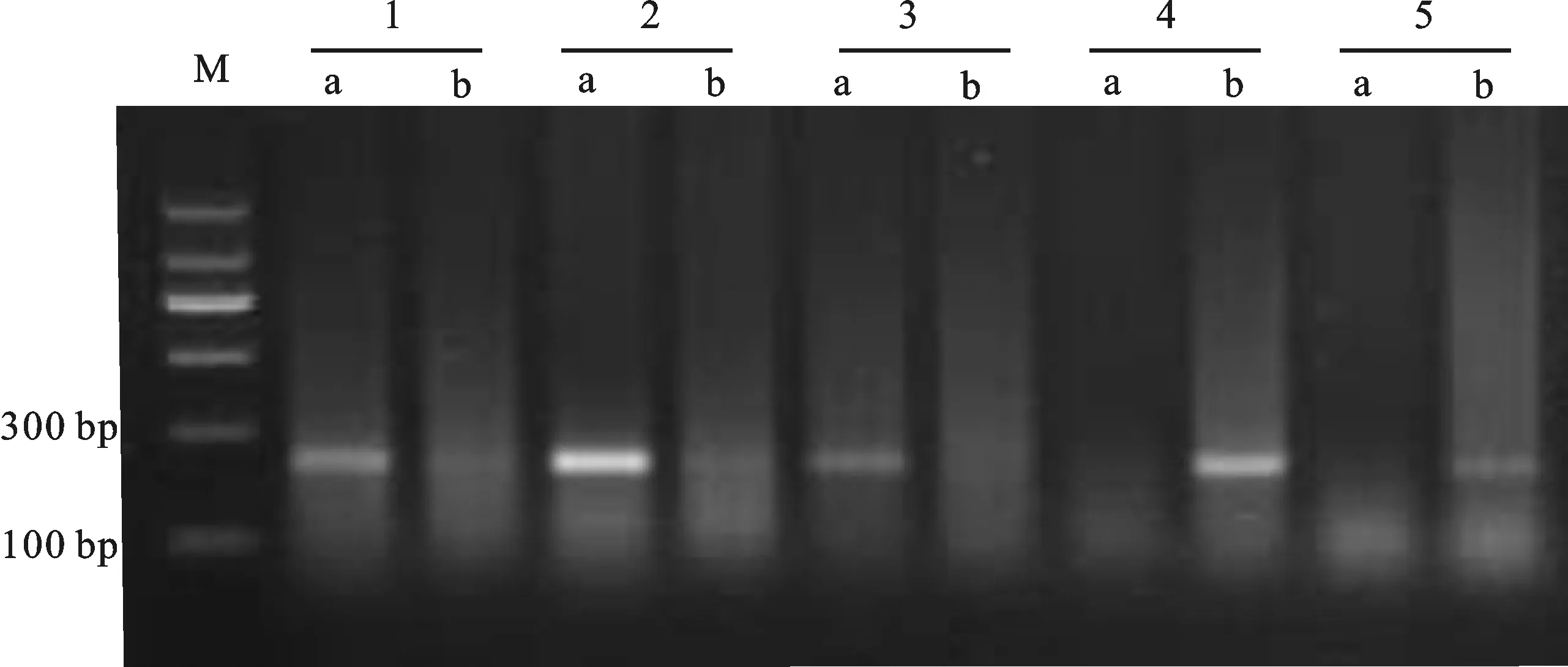
M:DL1200;a: Rht-B1a;b: Rht-B1b;1:冀麥449;2:農大5695;3:濟麥32;4:榮華901;5:石4185(CK)。
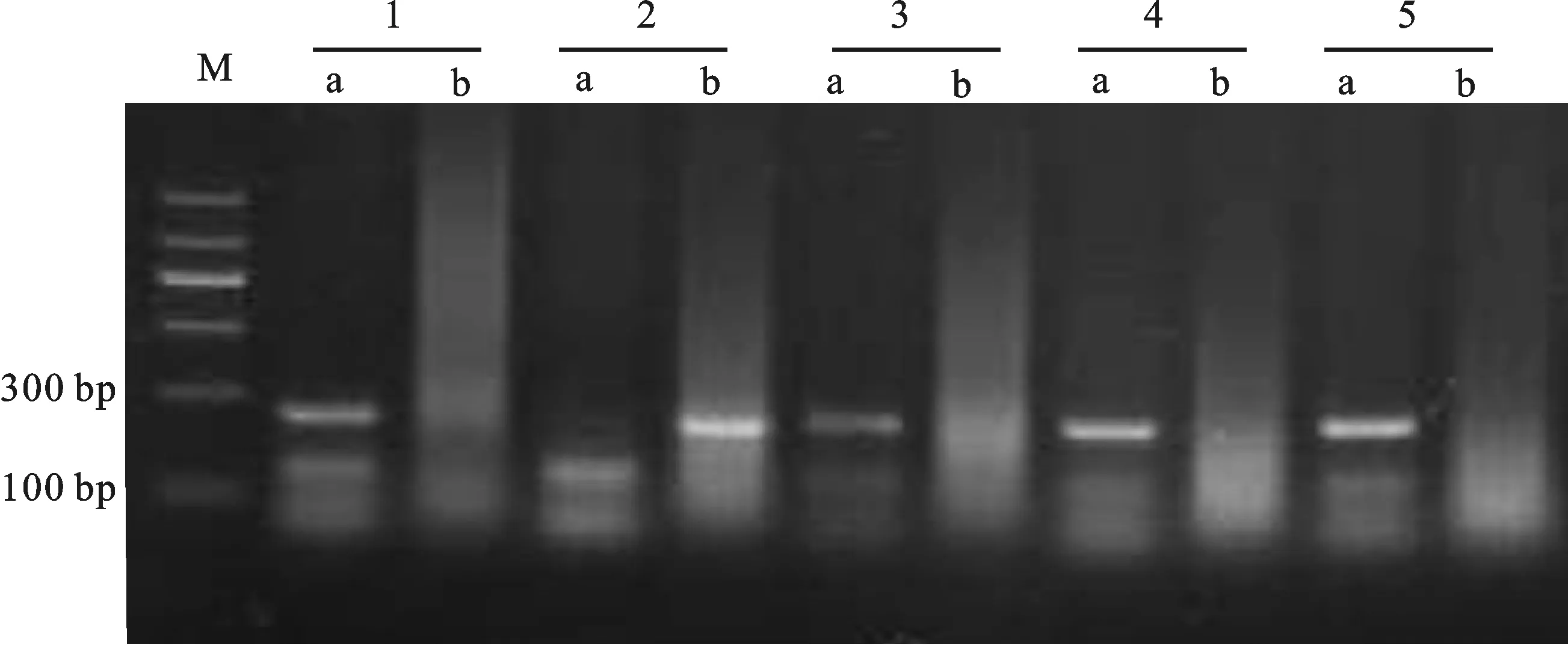
M:DL1200;a: Rht-D1a;b: Rht-D1b;1:新麥36;2:鄭麥7698(CK);3:洛麥27;4:蜀麥1628;5:華1168。
從矮稈基因在不同冬麥區的分布頻率來看,Rht-B1b在西南冬麥區的分布頻率最高,其次是在北部冬麥區和長江中下游冬麥區,在黃淮冬麥區分布頻率最低;Rht-D1b在長江中下游冬麥區的分布頻率最高,其次是在黃淮冬麥區和北部冬麥區,在西南冬麥區分布頻率最低;Rht8在各冬麥區均有分布,在長江中下游冬麥區分布頻率最高,其次是在北部冬麥區和黃淮冬麥區,西南冬麥區分布頻率最低(表2和表3)。
(續表2 Continued table 2)
矮稈基因及其組合在所有麥區(由于新疆冬春麥區只有1份小麥材料,未進行統計)的分布頻率由高到低依次為Rht-D1b(26.7%);Rht-D1b+Rht8(18.2%);Rht8(16.5%);Rht-B1b(7.2%);Rht-B1b+Rht8(6.8%);Rht-B1b+Rht-D1b+Rht8(2.1%);Rht-B1b+Rht-D1b(0.4%)。Rht-B1b在西南冬麥區的分布頻率最高,為 24.4%;Rht-D1b在黃淮冬麥區的分布頻率最高,為32.2%;Rht8在各麥區分布頻率相當,在14.9%~19.5%之間;Rht-B1b+Rht8在北部冬麥區分布頻率最高,為12.3%;Rht-B1b+Rht-D1b僅在黃淮冬麥區有分布,在其他麥區未檢測到該聚合類型。Rht-D1b+Rht8在長江中下游冬麥區和黃淮冬麥區分布頻率較高,分別為 23.5%和 22.3%;Rht-B1b+Rht-D1b+Rht8在長江中下游冬麥區分布頻率最高,為5.9%,在西南冬麥區未檢測到該聚合類型。此外,供試材料中有22.0%的材料均未檢測到Rht-B1b、Rht-D1b和Rht8,西南麥區的分布頻率最高,達 31.7%(表3)。
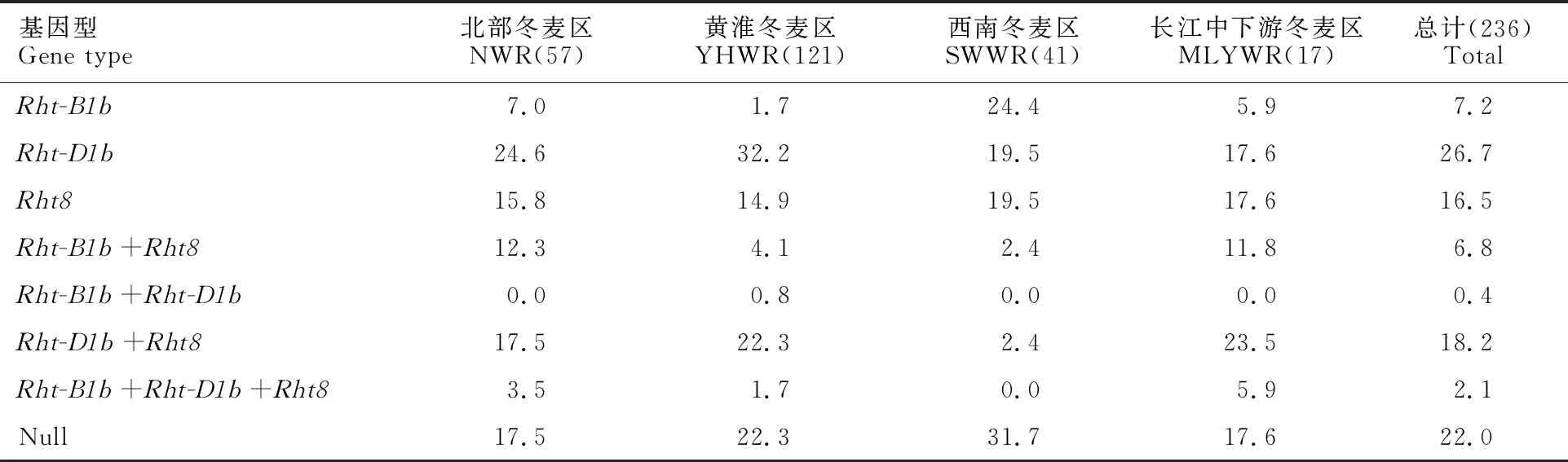
表3 矮稈基因 Rht-B1b、 Rht-D1b和 Rht8及其組合在不同麥區的分布頻率
2.2 Rht-B1b、 Rht-D1b和 Rht8與株高及產量性狀的相關性
不含目標基因的小麥株高最高,與Rht-D1b+Rht8和Rht-B1b+Rht-D1b+Rht8基因型小麥差異顯著;Rht-B1b+Rht-D1b+Rht8基因型小麥的株高最低,與Rht-B1b+Rht8和Rht-D1b+Rht8基因型小麥差異不顯著。不同矮稈基因型的降稈效果表現為Rht-B1b+Rht-D1b+Rht8;Rht-D1b+Rht8;Rht-B1b+Rht8;Rht-D1b;Rht-B1b;Rht8(表4和表5)。Rht-D1b+Rht8基因型小麥的穗長最短,與其他基因型以及不含目標基因的小麥差異顯著。Rht-B1b+Rht-D1b+Rht8基因型小麥的粒寬最大,顯著大于Rht-B1b、Rht-B1b+Rht8基因型以及不含目標基因的小麥。Rht-B1b+(Rht-D1b+)Rht8基因型小麥的千粒重最大,顯著高于Rht-B1b、Rht-B1b+Rht8基因型小麥。所有基因型及不含目標基因的小麥的粒長和穗粒數均無顯著差異(表4)。由于Rht-B1b+Rht-(D1b+Rht8)種質只有5份,結果還需進一步驗證。
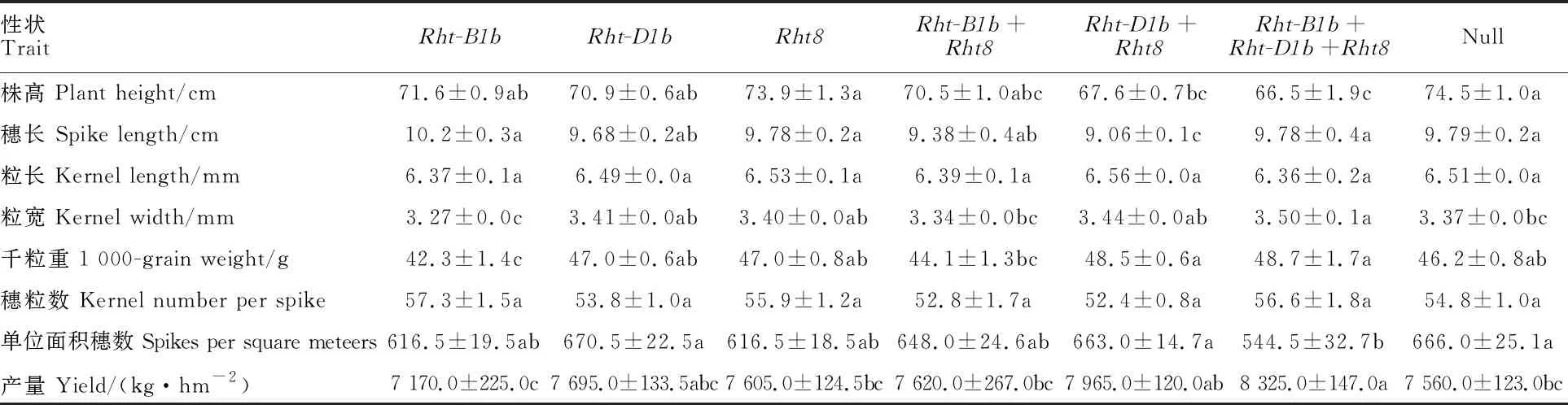
表4 Rht-B1b、 Rht-D1b和 Rht8及其組合對株高及產量相關性狀的影響
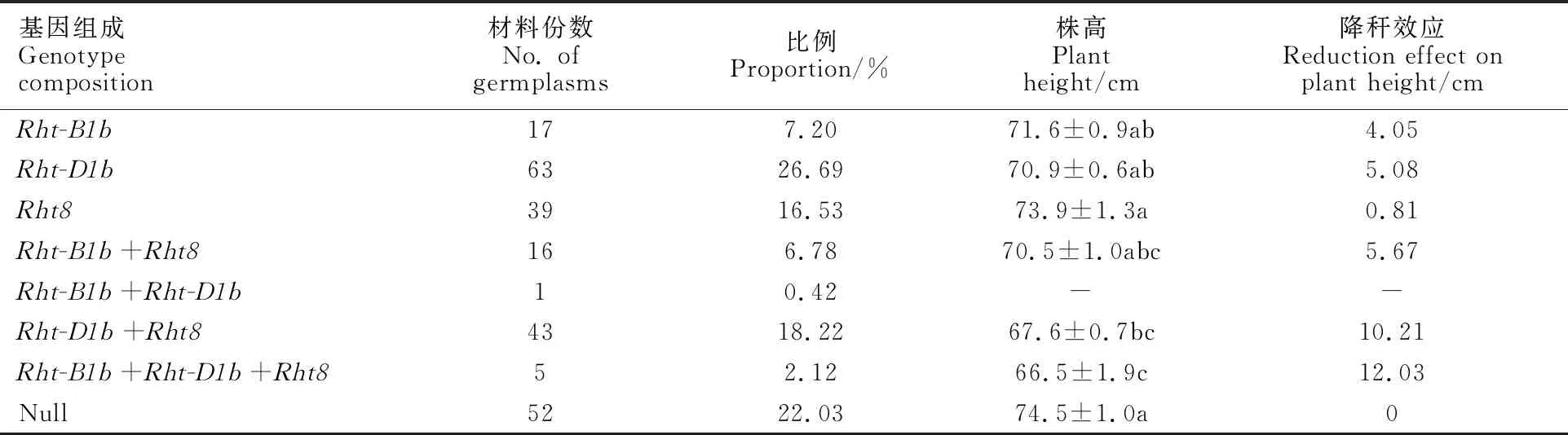
表5 矮稈基因 Rht-B1b、 Rht-D1b和 Rht8 及其組合的降稈效應
3 討 論
3.1 矮稈基因的分布頻率
本研究供試237份材料中,含有Rht-D1b、Rht-B1b和Rht8基因的材料占所有供試材料的 78.0%,可見,Rht-D1b、Rht-B1b和Rht8雖占據中國冬小麥矮稈基因的主導地位,但仍有一些其他矮稈基因在生產中得到一定規模應用。在本研究矮稈基因不明確的材料中,多數植株株高低于75 cm,屬于矮稈種質,說明含有其他矮稈基因。陳向東等[18]發現117份國內推廣小麥品種及中間種質中,矮稈基因的分布頻率為27.4%。周曉變[19]發現黃淮麥區主栽品種、歷史品種和農家種中,Rht4、Rht9和Rht12基因的分布頻率分別為 66.7%、40.7%和34.6%。可見,在中國主要冬麥區中,除了Rht-B1b、Rht-D1b和Rht8外, 其他矮稈基因分布也較為廣泛。
本研究中Rht-D1b和Rht8基因的分布頻率分別為47.9%和44.1%,而Rht-B1b的分布頻率僅為16.5%,說明Rht-D1b和Rht8應用更廣泛。楊松杰[4]和周 陽等[20]研究表明,Rht-D1b和Rht8基因在中國小麥的分布頻率分別為47.5%和 43.9%,與本研究研究結果基本一致。
3.2 不同麥區矮稈基因型的分布及其與產量性狀的關系
不同麥區間矮稈基因型的分布頻率存在明顯的差異。黃淮冬麥區主要以Rht-D1b(32.2%)、Rht8(14.9%)和Rht-D1b+Rht8(22.3%)為主。可見,黃淮冬麥區更偏好使用Rht-D1b,其次是Rht8。周 陽等[21]和楊松杰[4]也發現該麥區Rht-D1b的分布頻率高。曹廷杰等[21]和慕美財等[22]認為,這是由于Rht-D1b在黃淮冬麥區的小麥種質中已被固定所致。北部冬麥區矮稈基因型主要以Rht-D1b(24.6%)、Rht8(15.8%)和Rht-D1b+Rht8(17.5%)為主,西南冬麥區主要以Rht-B1b(24.4%)、Rht-D1b(19.5%)和Rht8(19.5%)為主,長江中下游冬麥區主要以Rht-D1b(17.6%)、Rht8(17.6%)和Rht-D1b+Rht8(23.5%)為主。其中,西南冬麥區是所有麥區中種植密度最低的生態區,植株個體發育健壯不易倒伏,不需要聚合多個矮稈基因降低株高來規避倒伏風險,并且以此來獲得更高生物量和產量。此外,Rht-B1b基因在西南冬麥區的分布頻率明顯高于其他麥區,且具有比其他基因型更高的穗粒數和較低的單位面積穗數,這也正是西南冬麥區小麥群體的產量結構特征。說明矮稈基因在不同生態區的分布與矮源產量結構特征和生態環境適應性有關。
本課題組研究也發現,黃淮冬麥區、西南冬麥區和長江中下游冬麥區矮稈基因Rht-B1b、Rht-D1b和Rht8野生型與突變型之間的產量因子差異均不顯著(數據未提供),表明不同矮稈基因與產量因子間并不存在相關性。顯著性分析發現,黃淮冬麥區單位面積穗數最高,北部冬麥區和長江中下游冬麥區千粒重較高,而這三個麥區Rht-D1b基因的分布頻率分別為57.0%、45.6%和47.0%,Rht8基因的分布頻率分布為43.0%、54.4%和58.8%;西南冬麥區的穗粒數最高,Rht-B1b基因的在該麥區的分布頻率為26.8%。可見,小麥植株具有所在生態區的產量結構特征,與矮稈基因本身無關。因此,各生態區矮稈基因的使用偏好性可能與矮稈基因被固定的先后順序有關,越早在某個生態區被固定的矮稈基因,其攜帶者越早具有該生態區的產量結構特征,故被利用的機率和分布頻率也就越高,從而造成矮稈基因與產量性狀關聯的遺傳假象。
3.3 Rht-B1b、 Rht-B1b和 Rht8及其組合對株高的影響
不同矮稈基因型的降稈效果從強到弱依次為:Rht-B1b+Rht-D1b+Rht8;Rht-D1b+Rht8;Rht-B1b+Rht8;Rht-D1b;Rht-B1b;Rht8。這與Tang 等[23]和馮 潔等[24]的研究結果一致。但Rht-B1b、Rht-B1b和Rht8單獨應用時,降稈效果不顯著。Rht8只有與Rht-D1b聚合(Rht-D1b+Rht8和Rht-B1b+Rht-D1b+Rht8)株高才顯著降低,而與Rht-B1b聚合時差異不顯著。同時,三個矮稈基因聚合類型(Rht-B1b+Rht-D1b+Rht8)的株高顯著低于僅含單個基因的株高,表明不同矮稈基因聚合時,降稈效果存在累加效應,并且Rht-D1b的降稈效果優于Rht-B1b,這與Sheoran等[25]和Robbins等[26]的研究結果基本一致。
總之,Rht-B1b、Rht-D1b和Rht8是中國小麥矮稈基因的主要類型。雖然Rht-D1b和Rht8基因型小麥總體上具有較高的千粒重和單位面積穗數,Rht-B1b基因型小麥具有較高的穗粒數,但矮稈基因本身與產量因子間無顯著相關性。矮稈基因聚合時降稈效果存在累加效應,且Rht-D1b的降稈效果優于Rht-B1b。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
