食物濃度對來自不同營養水平水體盔型溞克隆生活史參數的影響
2023-04-29 23:52:38黃小娥郭詩元林秋奇
水生態學雜志 2023年5期
黃小娥 郭詩元 林秋奇

摘要:探討浮游動物對食物濃度變化的生態適應性,為了解環境中食物濃度對浮游動物生存、進化的影響提供理論依據。于2018年5月采集廣東流溪河水庫、廣西巖灘水庫和洪潮江水庫(營養水平分別為貧-中營養型、中營養型和富營養型)盔型溞克隆,比較分析了高食物濃度(以碳濃度計為2 mg/L)和低食物濃度(以碳濃度計為0.5 mg/L)的斜生柵藻(碳磷比為105:1)對這3個水庫的盔型溞克隆生活史參數的影響。在高食物濃度處理組,3個克隆幼溞生長速率、首窩和第二窩產仔量和內稟增長率均顯著高于低食物組,首次生殖時間和新生幼溞體長顯著小于低食物組,首次懷卵體長在2個處理間無顯著差異。食物的增加縮短了性成熟時間、增加產仔量,從而提高內稟增長率。首次生殖時間在3個克隆間沒有顯著差異,但來自中營養水體克隆的產仔量在2個食物處理組中均顯著低于其他2個克隆,導致中營養水體克隆內稟增長率小于其他2個克隆。貧-中營養水體克隆在2個食物處理組中內稟增長率均與富營養水體克隆沒有顯著差異。結果表明,3個盔型溞克隆對食物濃度變化均表現出較高的表型可塑性,但在生活史策略上并未表現出適應性進化。
關鍵詞:枝角類;生活史策略;食物數量;富營養化
中圖分類號:Q178.1? ? ? ? 文獻標志碼:A? ? ? ? 文章編號:1674-3075(2023)05-0084-08
富營養化是淡水水體普遍面臨的環境問題,它可以改變浮游植物的數量和質量,從而通過食物鏈對浮游動物產生強烈的選擇壓力(Hairston et al, 1999; Mao et al, 2020)。浮游植物質量變化包括化學計量比、大小可食性、不飽和脂肪酸含量等(Enache et al, 2019)。富營養化對浮游植物的影響主要包括:降低浮游植物碳磷比,提高浮游植物的化學計量學質量(De Senerpont Domis et al, 2014);增加浮游植物豐度,提高其總生物量(Van Meerssche & Pinckney, 2019);使浮游植物群落結構轉變為以藍藻為優勢類群,從而降低群落多不飽和脂肪酸含量,增加藍藻毒素含量(Sommer et al, 1986; Demott & Müller‐Navarra, 1997)等。如果環境中食物條件的變化是短期的,浮游動物可通過表型可塑性來調整生活史策略以應對(Stearns, 1989);如果食物變化是長期且持續的,浮游動物可通過微進化、改變種群在世代過程中等位基因的頻率來適應(Weider et al, 2005)。經自然選擇后生活于當前環境的個體,較之其他水體環境中的克隆而言對該水體有更高的適合度。如Hairston等(1999)在20世紀60-70年代對歐洲中部的康斯坦茨湖沉積物中的盔型溞(Daphnia galeata)休眠卵進行了孵化,發現富營養化期間產生的克隆比貧營養期產生的克隆對產毒藍藻具有更高的抗性。隨后的研究發現,富營養化期間產生的克隆在產毒藍藻環境下始終具有較高的適合度(Hairston et al, 2001)。Frisch等(2014)發現富營養化導致的浮游植物化學計量比變化使美國明尼蘇達南中心湖多蚤溞(Daphnia pulicaria)的種群遺傳結構發生變化。沉積物中年輕基因型(2002-2008年)的多蚤溞對磷的利用效率和滯留率均低于古老基因型(1301-1418年);年輕基因型在高磷食物下的適合度高于古老基因型,而古老基因型在低磷食物下的適合度大于年輕基因型。
溞屬(Daphnia)是枝角類中的重要類群,具有分布范圍廣、濾食能力強、個體相對較大等特點;在環境條件良好的情況下進行孤雌生殖,因而也容易發生等位酶變異,是生態學和生物進化研究的重要對象和模式生物(Lampert, 2011)。以往研究已表明,在富營養化的過程中,溞屬種類對于食物中磷含量的變化以及藍藻毒素的變化產生了適應性進化(Frisch et al, 2014; Hairston et al, 1999),生活于富營養時期的溞類個體比貧營養時期的個體在高磷環境下有更高的生長速率,且對藍藻有更高的抗性。那么溞類在面臨富營養化所造成的食物數量的變化時是否也存在這種適應性進化?本研究以中國南亞熱帶水庫常見的大型枝角類盔型溞(Daphnia galeata)為研究對象,從空間尺度上,選取了3個來源地營養水平不同的盔型溞克隆,研究三者在不同的食物濃度下的生活史差異,探討食物數量在盔型溞對富營養化過程的適應中所產生的影響,為更好地了解環境中食物數量的變化對浮游動物生存、進化的影響提供理論依據。
1? ?材料與方法
1.1? ?實驗動物與食物的培養
實驗選用的流溪河水庫克隆(LXH克隆)、巖灘水庫克隆(YT克隆)及洪潮江水庫克隆(HCJ克隆)均于2018年5月分別采自于廣東流溪河水庫(113°47'E,23°45'N)、廣西巖灘水庫(107°30'E,24°3'N)和洪潮江水庫(109°7'E,21°50'N)。其營養水平分別為貧-中營養型、中營養型和富營養型(Lin et al, 2018; 周晴等, 2015; 唐匯娟等, 2021)。用孔徑113 μm的浮游生物網由水體底部至表層垂直拖取,從采集到的水樣中,隨機挑選出20只盔型溞并進行單獨培養。取它們的下一代個體進行線粒體COI基因測序,挑選出各個水體的優勢克隆。在實驗室,每個克隆只挑選出1個雌性個體,喂以1 mg/L(食物濃度均以碳濃度計) 的斜生柵藻(Scenedesmus obliquus)進行單克隆培養。培養用水為經孔徑 0.22 μm的濾膜過濾后的流溪河水庫水,流溪河水庫為貧-中營養型水庫,沒有藍藻毒素分布,經0.22 μm濾膜過濾后水的水質狀況不會對實驗結果造成干擾。培養條件為:光照強度1 200 lx,光周期 L∶D = 12 h: 12 h,溫度25℃。
本實驗所選用的食物為斜生柵藻,在光周期L : D = 12 h: 12 h和25℃條件下用標準CMOBO培養基培養(Kilham et al, 1998)。在對數生長期,將斜生柵藻藻液以3 000 r/ min 的速度離心10 min,去掉上清液,加無菌蒸餾水再次離心去掉上清液,隨后加入到適量的無氮、無磷的CMOBO培養基中,于4℃條件下避光保存。取少量藻液在孔徑為0.22 μm的玻璃纖維濾膜上過濾,在60℃烘箱中烘干24 h,稱重,用元素分析儀(vario EL cube, 德國)測定柵藻碳含量。用鉬酸銨分光光度法測定藻細胞磷含量,可知該時期藻細胞的碳磷原子比為105:1,可確保3個盔型溞克隆不受食物磷限制,以便比較分析食物數量對盔型溞克隆的影響。
1.2? ?實驗處理
實驗開始的前1 d,將即將孵化出幼溞的母溞轉移到培養用水中,獲取足夠數量的齡期小于24 h的幼溞,每個克隆隨機挑出20只新生幼溞測量它們的體長。實驗設計3個動物處理組:LXH克隆、HCJ克隆和YT克隆;2個食物濃度處理組:0.5 mg/L和2 mg/L。參考Pajk等(2018)的方法,采用單個體培養實驗,培養容器為50 mL的燒杯,實驗用水為經孔徑0.22 μm的濾膜過濾后的流溪河水庫水,體積為30 mL。每只燒杯投放1只齡期小于24 h的幼溞,每個處理組設置20個平行。培養條件與母體相同。每天觀察盔型溞的生長和懷卵情況,并將其轉移至新配制的含有相應食物濃度的過濾流溪河水庫水中,記錄母溞的存活情況及所產幼仔數,并將幼溞移除;測量母溞首次懷卵體長、首窩和第二窩幼溞體長。
1.3? ?數據處理與分析
編制生命表,計算首次懷卵時間,特定齡期存活率(lx)、特定齡期生殖率(mx)、第一窩產仔量和第二窩產仔量等參數。種群內稟增長率(r)根據歐拉方程計算獲得:
式中:lt為實驗動物首次懷卵體長,l0為實驗動物的初始體長,t為首次生殖時間。
在R4.0.3中用arm程序包進行統計分析。對數據進行lg x對數轉換,使各處理組數據呈正態分布且方差齊性。采用雙因素方差分析和Tukey多重比較來比較食物濃度和克隆對生活史參數的影響。數據均為mean[±]SD,以不同字母表示差異顯著(P<0.05),大小按字母排序,即a>b>c,在圖中顯示方差分析中克隆(c)、食物(F)和它們的相互作用(c[×]F)影響的F值和P值。
2? ?結果與分析
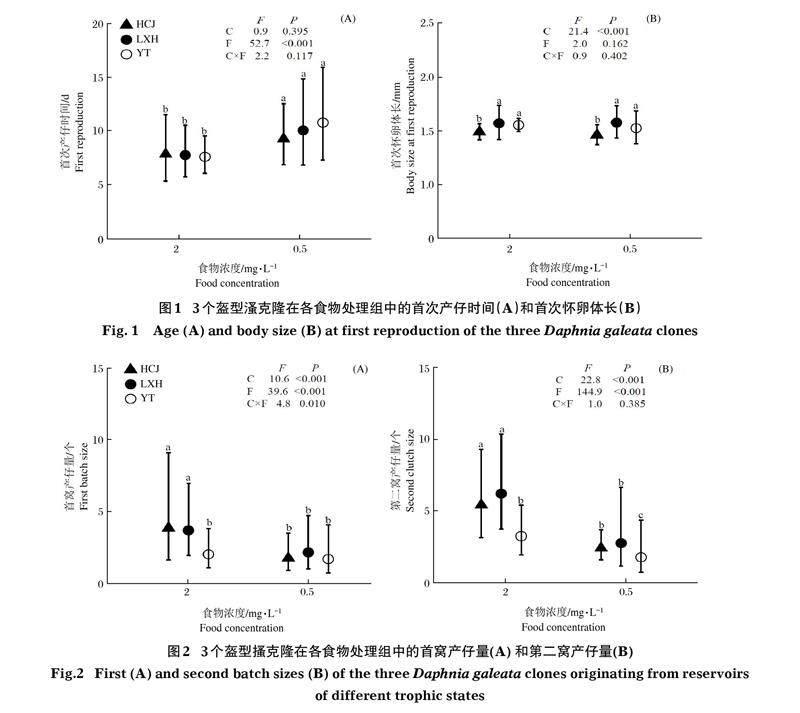
HCJ、LXH和YT克隆的首次產仔時間分別為8.4~9.7、8.3~10.7和7.4~10.7 d。3個克隆在低食物濃度處理組首次產仔時間均顯著長于高食物濃度處理組(P<0.001);在相同食物處理組,首次產仔時間在3個克隆間沒有顯著差異(圖1A)。克隆與食物數量對首次產仔時間的影響不存在顯著的交互作用。HCJ、LXH和YT克隆的首次懷卵母溞體長分別為1.45~1.49、1.56~1.57 和1.53~1.54 mm。3個克隆的首次懷卵母溞體長在2個食物濃度處理組間沒有顯著差異;在同一食物處理組,LXH克隆和YT克隆的首次懷卵母體體長沒有顯著差異,而HCJ克隆首次懷卵母溞體長顯著小于LXH和YT克隆(P<0.001)(圖1B)。克隆與食物數量對首次懷卵母溞體長的影響不存在顯著的交互作用。
HCJ、LXH和YT克隆的首窩產仔量分別為1~6、1~6和1~3個。YT克隆首窩產仔量在2個食物濃度處理組間無顯著差異,LXH和HCJ克隆首窩產仔量高食物濃度處理組顯著高于低濃度處理組(P<0.001)(圖2)。克隆與食物數量對首窩產仔量的影響存在顯著的交互作用(P<0.05)。在高食物濃度處理組中,HCJ和LXH克隆首窩產仔量差異不顯著,YT克隆首窩產仔量顯著少于HCJ和LXH克隆(P<0.001)。在低食物濃度處理組中,3個克隆間的首窩仔量無顯著差異。HCJ、LXH和YT克隆的第二窩產仔量分別為1~8、1~9和1~6個。3個克隆的第二窩產仔量在高食物濃度處理組均顯著高于低食物濃度處理組(P<0.001);在不同食物濃度處理組中,HCJ克隆和LXH克隆第二窩產仔量均無顯著差異,而YT克隆第二窩產仔量均顯著少于HCJ和LXH克隆(P<0.001)。克隆與食物數量對第二窩產仔量的影響不存在顯著的交互作用。
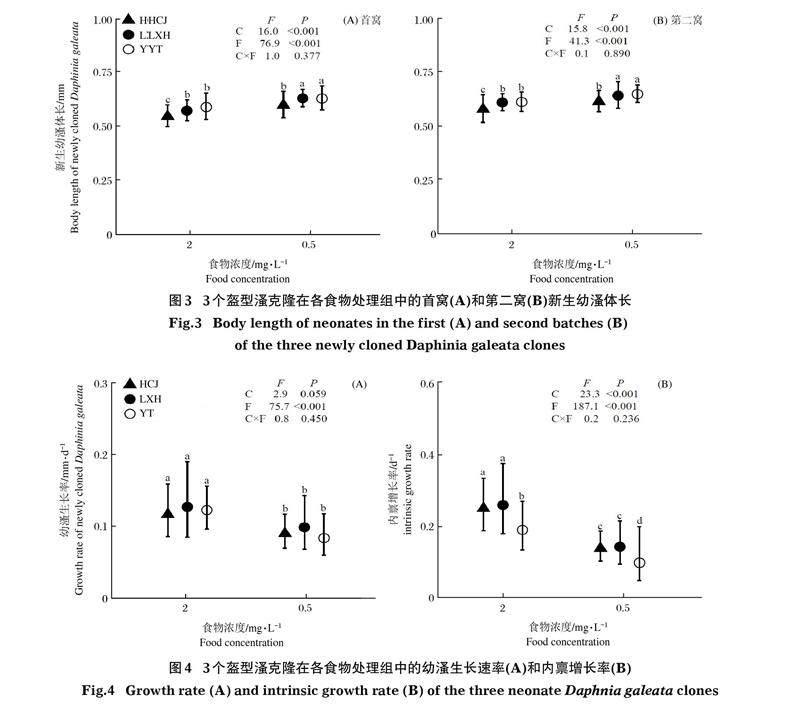
HCJ、LXH和YT克隆的首窩新生幼溞體長分別為0.55~0.60、0.57~0.63和0.59~0.63 mm。3個克隆的首窩新生幼溞體長在高食物濃度處理組均顯著小于低食物濃度處理組(P<0.001);在同一食物處理組中,LXH克隆和YT克隆之間沒有顯著差異,而HCJ克隆顯著小于LXH和YT克隆(P<0.001)(圖3A)。克隆與食物濃度對首窩產新生幼溞體長的影響不存在顯著的交互作用。HCJ、LXH和YT克隆第二窩新生幼溞體長分別為0.58~0.61、0.61~0.64和0.61~0.65 mm。3個克隆的第二窩新生幼溞體長在高食物濃度處理組顯著小于低食物濃度處理組(P<0.001);在同一食物處理組中,LXH和YT克隆之間沒有顯著差異,而HCJ克隆顯著小于LXH和YT克隆(P<0.001)(圖3B)。克隆與食物濃度對第二窩產新生幼溞體長的影響不存在顯著的交互作用。
HCJ、LXH和YT克隆的幼體生長速率分別為0.18~0.54、0.21~0.53和0.19~0.47 mm/d。3個克隆的幼體生長速率在高濃度處理組顯著高于低濃度處理組(P<0.001);在同一食物處理組中,3個克隆的幼體生長速率沒有顯著差異(圖4A)。克隆與食物數量對幼溞體生長速率的影響不存在顯著的交互作用。HCJ、LXH和YT克隆的種群內稟增長率分別為0.08~0.36、0.05~0.37和0.05~0.29 d-1。3個克隆的種群內稟增長率在高濃度處理組均顯著高于低食物濃度處理組(P<0.001);在同一食物濃度處理組中,LXH克隆和HCJ克隆間內稟增長率無顯著差異,二者均顯著高于YT克隆(P<0.001)(圖4B)。克隆與食物數量對內稟增長率的影響不存在顯著的交互作用。
