寒地水稻龍粳31耐冷特性及油菜素內酯的耐冷調節作用
2023-05-26 01:43:40曾維康魏嘉琪過一馳程一珊姜樹坤趙明輝耿麗娟郭志富
種子 2023年3期
關鍵詞:水稻
高 洋, 曾維康, 魏嘉琪, 過一馳, 程一珊, 姜樹坤, 趙明輝, 耿麗娟, 郭志富
(1.沈陽農業大學生物科學技術學院, 沈陽 110866;2.黑龍江省農業科學院耕作栽培研究所, 哈爾濱 150086;3.沈陽農業大學水稻研究所, 沈陽 110866; 4.沈陽市食品藥品檢驗所, 沈陽 110000)
低溫冷害是常見的非生物逆境之一,對水稻(OryzasativaL.)的生長發育影響巨大,是限制水稻產量穩定和提高的主要環境因素[1]。寒地水稻因特殊的選育和生長環境,具備更好的低溫耐受潛力和基礎[2],其主要在東北稻區尤其是黑龍江稻區種植,而黑龍江稻區作為我國最大的商品粳稻生產基地,水稻種植面積和產量均為全國第一,近三年水稻種植面積約占全國粳稻面積的50%[3],對保障國家糧食安全具有重要的戰略意義。
多年來,針對高緯度低積溫以及冷害頻發等問題,東北地區水稻育種工作者選育出很多熟期早、抗性強、產量高的寒地水稻品種,為東北稻區水稻產量的穩定和提高奠定了基礎,同時為深入開展水稻耐冷基礎理論研究提供了理想的研究材料。龍粳、龍稻、綏粳、墾稻等系列品種,均在東北稻區有一定的種植面積[4]。在眾多寒地水稻品種中,龍粳31因其高產、優質、抗逆性強等顯著特點,多年來一直是全國粳稻種植面積最大的品種,已累計推廣超過130萬hm2。目前,針對龍粳31的基礎理論研究主要集中在栽培措施的優化上,如種植密度調整、肥料配比優化、旱直播條件摸索等方面,最近也出現了龍粳31遺傳基礎分析以及利用基因編輯技術對其進行品質改良探索的相關報道[5-8]。龍粳31作為種植面積最大的寒地水稻品種,有很強的耐冷性,但目前對于龍粳31低溫耐受表型特征及生理生化等方面的研究較為少見,限制了對寒地水稻耐冷機理的深入了解。
油菜素內酯(Brassinolides,BRs)是甾醇類植物激素中最具活性的一種,可有效提高植物的抗冷性。研究表明,外源噴施BRs對于水稻不同時期耐冷性的提高均有顯著的促進作用[9-12]。目前,BRs處理與寒地水稻耐冷性關系方面的研究尚未見報道。基于以上分析,本研究以耐冷性較強的寒地水稻龍粳31和冷敏感水稻9311為研究材料,在芽期和苗期分別鑒定其在冷處理以及低溫和BRs聯合處理下的耐冷表型性狀變化,并測定耐冷相關生理生化指標及內源激素含量的變化情況,為寒地水稻耐冷生理基礎及BRs途徑下水稻耐冷調控機制的明確提供參考。
1 材料與方法
1.1 試驗材料
本研究以寒地水稻龍粳31和冷敏感水稻9311為試驗材料。龍粳31由黑龍江農科院水稻研究所提供,9311由沈陽農業大學水稻研究所提供。
1.2 試驗方法
1.2.1低溫脅迫及BRs處理后低溫發芽率測定
試驗分為三個處理,常溫對照為25 ℃(ck)、低溫處理溫度為17 ℃、聯合處理為BRs(0.01 μmol/L)+17 ℃低溫處理。選取50粒健康飽滿的水稻種子,在45 ℃烘箱處理2 d打破休眠后,分別使用蒸餾水和BRs(0.01 μmol/L)進行24 h浸種處理。將浸種后的種子置于放有濕潤濾紙的培養皿中分別放入25 ℃和17 ℃光照培養箱中處理7 d。
1.2.2低溫脅迫及BRs處理后苗期表型的測定
試驗分為3個處理,常溫25 ℃、4 ℃低溫處理、聯合處理為BRs(0.1 μmol/L)+4 ℃低溫處理。選取健康飽滿的種子,在45 ℃烘箱里放置3 d打破種子休眠后,放在墊有濕潤濾紙的玻璃培養皿里,于35 ℃恒溫箱中黑暗催芽72 h,種在混合基質土(V基質土∶V蛭石∶V珍珠巖=2∶2∶1)的栽培盆(7 cm×7 cm×7 cm),在25 ℃培養室內生長21 d,長至三葉期,將4 ℃、4 ℃+BRs(處理前24 h,用0.1 μmol/L BRs進行根際灌溉和葉面噴施)兩種處理,置于10 ℃低溫培養箱中處理7 d后,常溫恢復7 d,將表型與ck進行比較。
1.2.3低溫脅迫及BR處理后苗期葉片生理生化指標測定
將前述苗期低溫和聯合處理的水稻幼苗按0、1、3、5、7 d進行葉片取樣,分別進行脯氨酸(Pro)含量、丙二醛(MDA)含量、相對電導率、超氧化物歧化酶(SOD)活性測定[14-15]。
1.2.4低溫脅迫及BR處理后苗期葉片生理生化指標測定
苗期低溫和聯合處理3 d后,對幼苗葉片進行取樣,使用預冷卻的研缽將0.5 g樣品勻漿,然后在4 mL提取溶液中進行超聲輔助提取45 min。12 000 r/min離心10 min后,收集上清液,重復抽提一次。用2 mL甲醇和2 mL甲酸(1 mol/L)預活化HLB和MCX柱。然后,將500 μL溶液通過0.22 μm過濾器過濾,并轉移到2 mL的LC-MS瓶中進行UPLC-MS/MS分析[16]。
2 結果與分析
2.1 低溫脅迫及BRs處理對水稻發芽率的影響
由圖1可知,龍粳31的ck、低溫處理和BRs+低溫處理的發芽率分別為100.00%、93.33%、97.33%,而9311的ck、低溫處理和BR+低溫處理的發芽率分別為100.00%、82.67%、86.00%,冷處理的龍粳31和9311發芽率分別降低6.67%和17.33%,證明龍粳31的低溫發芽率要高于9311,具備更好的芽期耐冷基礎。BRs浸泡后進行冷處理的龍粳31和9311發芽率分別降低4.29%和4.03%,證明低溫條件下,BRs對水稻種子發芽率有一定的抑制作用。
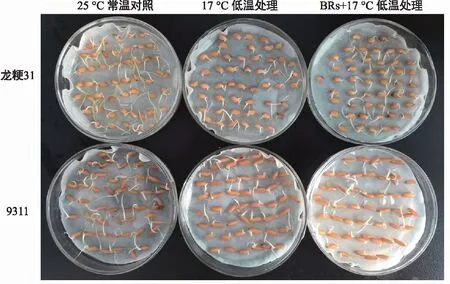
圖1 低溫發芽率表型Fig.1 Germination rate phenotype under cold temperature

注:每組材料從左到右分別為25 ℃常溫對照,4 ℃冷處理,BRs+4 ℃冷處理。A為冷處理3 d;B為冷處理7 d;C為恢復生長3 d;D為恢復生長7 d。圖2 苗期耐冷表型Fig.2 Phenotype with cold tolerance at seedling stage
2.2 低溫脅迫及BRs處理后苗 期表型變化
苗期耐冷表型鑒定結果表明,冷處理7 d后,龍粳31和9311的生長均受到明顯的抑制。當恢復生長3 d時,9311出現枯萎現象,而龍粳31長勢明顯好于9311,當恢復生長7 d時,9311已經干枯死亡,龍粳31仍然長勢良好,說明龍粳31具有更強的苗期耐冷性。與單獨冷處理相比,BRs+冷處理后明顯緩解了低溫對龍粳31的生長限制,而對于9311的緩解作用并不明顯(圖2)。
2.3 低溫脅迫及BRs處理對水稻生理指標的影響
為了進一步探究寒地水稻耐冷生理變化,對低溫及BRs處理下的SOD活性、Pro和MDA含量和相對電導率進行鑒定。常溫下,龍粳31和9311幼苗的SOD活性相近,隨著低溫處理時間的延長,兩種材料的SOD活性均呈現先升高后降低的趨勢,且龍粳31的上升幅度明顯高于9311。0.1 μmol/L的BR處理后,龍粳31和9311在低溫脅迫后SOD活性同樣呈先上升再下降的趨勢,而上升幅度大于單獨低溫處理條件(圖3 A);常溫下,龍粳31幼苗的Pro含量始終高于9311;在低溫處理后兩材料的Pro含量均呈上升趨勢,相比而言,龍粳31上升幅度更大,說明在龍粳31受低溫脅迫后Pro有更多的積累,可能比9311更具耐冷生理基礎。與單獨低溫處理相比,BRs+低溫處理下龍粳31的Pro含量整體變化不大,但9311的Pro含量顯著提高(圖3 B);常溫下,龍粳31和9311幼苗的MDA含量相近;隨低溫處理時間的延長,龍粳31和9311的MDA含量累積逐漸增多,且龍粳31上升程度明顯低于9311,說明龍粳31可能具有更強的清除膜脂過氧化物的能力;與單獨低溫處理相比,BRs+低溫處理后MDA含量均呈下降趨勢,且9311下降幅度更為明顯(圖3 C);常溫下,龍粳31幼苗的相對電導率和9311近似;隨著低溫處理時間延長,龍粳31和9311的相對電導率均顯著上升,且9311較龍粳31的相對電導率上升幅度更大,說明9311的質膜對低溫脅迫可能更敏感。與單獨低溫處理相比,BRs+低溫處理可以降低龍粳31和9311的相對電導率,且對9311的作用更為明顯(圖3 D)。
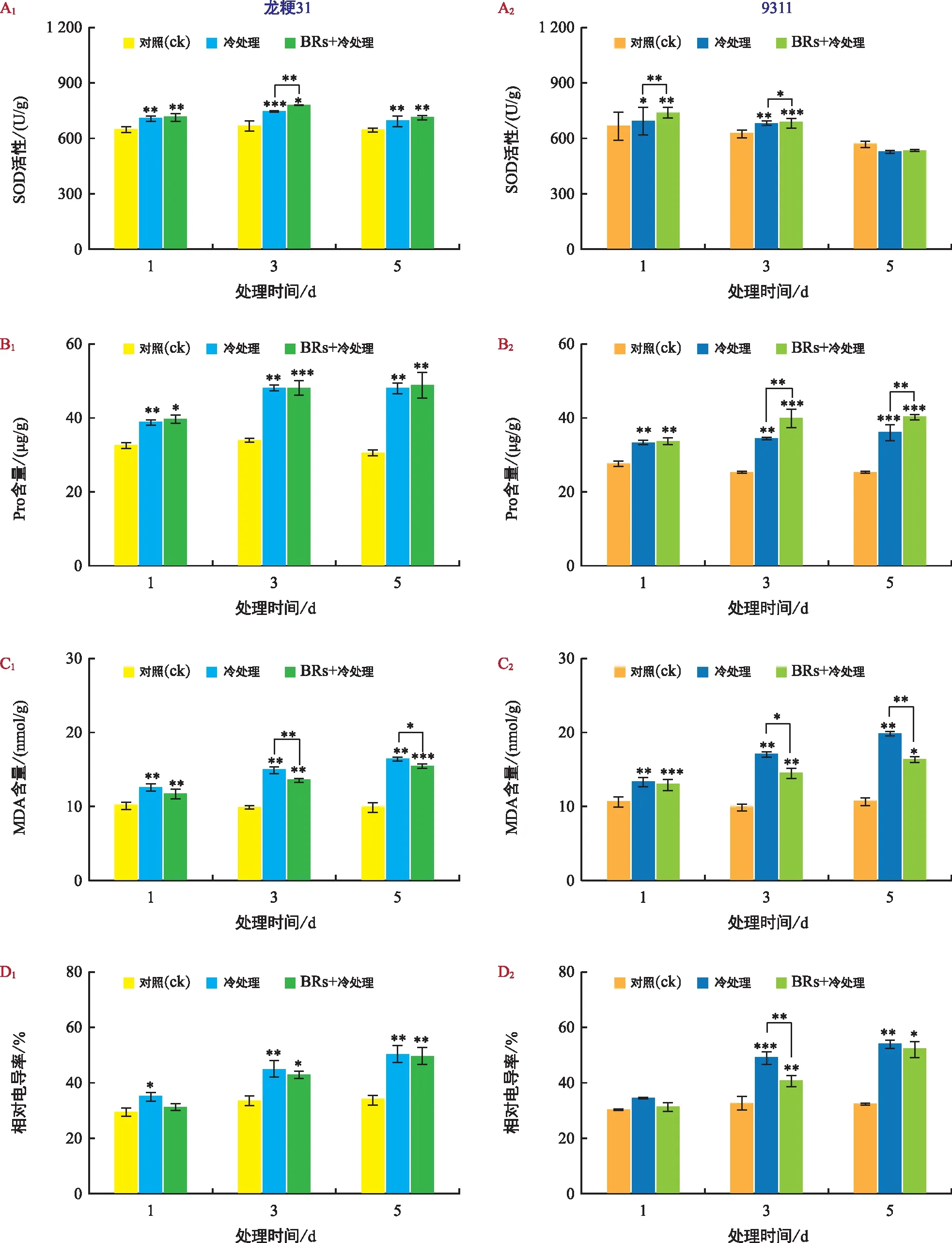
注:A為SOD活性;B為Pro含量;C為MDA含量;D為相對電導率。“*”表示p<0.05,“**”表示p<0.01,“***”表示p<0.001。下同。圖3 不同處理下生理指標變化Fig.3 Changes of physiological indicators under different treatments
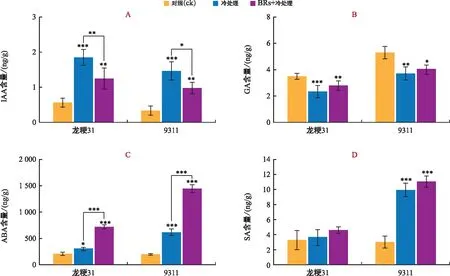
注:A為IAA含量;B為GA含量;C為ABA含量;D為SA含量;“*”表示p<0.05,“**”表示p<0.01,“***”表示p<0.001。圖4 不同處理下植物激素含量變化Fig.4 Changes of the plant hormone content under different treatments
2.4 低溫脅迫及BRs處理對水稻內源激素的影響
在不同處理下,對植物低溫響應相關的激素IAA、GA、ABA和SA的含量分別進行測定。結果表明,常溫下龍粳31幼苗IAA含量要高于9311。低溫處理后,龍粳31和9311的IAA含量均顯著上升,分別提高了228.44%和338.62%,9311的上升幅度更大。與單獨低溫處理相比,BRs+低溫處理下龍粳31和9311的IAA含量分別降低了32.58%、33.56%(圖4 A);GA含量的變化情況顯示,常溫下9311幼苗GA含量高于龍粳31。低溫處理后,龍粳31和9311的GA含量均顯著下降,分別下降了39.20%和30.23%,龍粳31的下降幅度更大。與單獨低溫處理相比,BRs+低溫處理下龍粳31和9311的GA含量分別升高了20.76%和11.43%,且龍粳31上升幅度更大(圖4 B);針對ABA含量而言,常溫下龍粳31和9311幼苗ABA含量相近。低溫處理后,龍粳31和9311的ABA含量均呈上升趨勢,分別提高了44.78%和214.99%,9311上升幅度更大。與單獨低溫處理相比,BRs+低溫處理下龍粳31和9311的ABA含量分別升高了140.40%和135.87%(圖4 C);而SA含量在常溫下龍粳31和9311幼苗相近。低溫處理后,龍粳31和9311中SA含量均有所上升,分別提高了12.61%、229.12%,9311的上升幅度顯著高于龍粳31。與單獨低溫處理相比,BRs+低溫處理下龍粳31和9311中SA含量分別升高了10.48%和25.52%,9311上升幅度更大(圖4 D)。
綜合來看,IAA、ABA和SA含量均在低溫處理后有所提升,且9311上升幅度更為顯著。相反地,GA含量在低溫處理后呈下降趨勢,且龍粳31下降幅度更大。與單獨低溫處理相比,BRs+低溫處理使ABA、SA和GA含量有所提升,9311提升的幅度更大,而BRs+低溫處理后IAA呈下降趨勢。這些結果表明,龍粳31內源激素受低溫影響的敏感程度要低于9311,而外源施加BR會對大部分植物激素含量產生影響。
3 討論與結論
3.1 寒地水稻表型性狀與耐冷性的關系
近年來,研究人員多以低溫處理后的發芽率、成苗率、發芽系數、芽期根長、苗長和第一葉長等表型指標評價寒地水稻的芽期耐冷性;以死葉率、葉片赤枯度、葉片卷曲度、根長、苗長、側根數、干重及鮮重等表型指標進行水稻苗期耐冷性鑒定[17-19]。在本研究中,龍粳31與9311的低溫發芽率有所降低,但龍粳31的降低幅度要小于9311。對于苗期而言,9311低溫處理后恢復,出現了瀕臨枯死的狀態,但龍粳31低溫處理恢復后植株依舊長勢良好。這些結果表明,寒地水稻龍粳31在芽期和苗期均具有更好的耐冷能力。外源施加BRs能緩解低溫對水稻芽期和苗期生長的不利影響,進一步說明BRs處理對于水稻耐冷表型有一定的緩解作用。
3.2 外源BRs及低溫脅迫對水稻幼苗生理生化指標的影響
研究表明,低溫逆境會減少水稻葉綠素的合成,增加ROS、MDA和可溶性糖含量以及電解質滲漏率及細胞膜透性等,同時也可以顯著提高SOD、POD和CAT等抗氧化酶活性,進而賦予植物對低溫脅迫的抵抗力[1,14-15]。外源施加BRs可顯著改善低溫誘導的氧化脅迫,降低MDA含量,提高POD、SOD的活性[9-12]。本研究中,龍粳31和9311在遭遇低溫脅迫期間,均可通過增加滲透調節物質含量來提高自身耐冷性。隨著低溫處理時間的延長,在一定時間內,Pro含量繼續呈上升趨勢,隨著冷脅迫時間不斷加長,Pro含量開始逐漸減少。BRs+低溫處理條件下,0.1 μmol/L BRs處理可以影響低溫下龍粳31和9311的Pro含量,但影響并不顯著。低溫脅迫下龍粳31和9311的MDA累積逐漸增多,相對電導率顯著上升,但相對而言龍粳31的上升幅度更小,說明龍粳31清除膜脂過氧化物的能力可能更強。針對BRs+低溫處理條件來分析,0.1 μmol/L的BRs處理初期,龍粳31和9311在低溫下MDA的含量和相對電導率均有所降低,這也為BRs提高水稻耐冷性提供了生理層面的證據。低溫脅迫初期水稻可以通過增加抗氧化酶活性來提高自身耐冷性,隨著低溫處理時間延長,龍粳31和9311的SOD活性均呈現先上升再下降趨勢,說明植物體內酶活性存在動態平衡,超出植物承受范圍,平衡遭到破壞。綜合來看,龍粳31與9311相比,更具耐冷生理基礎,而外源BRs處理可使水稻生理指標產生利于耐冷性提高的變化趨勢。
3.3 外源BRs以及低溫脅迫對水稻幼苗內源激素含量的影響
近年來,激素在植物低溫脅迫應答中的作用逐漸受到重視。外源施加BRs可顯著影響低溫脅迫下內源激素的含量變化,有研究表明,植物激素參與植物對低溫的響應,包括ABA、JA、IAA、GA、BRs、SA、C2H4等[9-12]。低溫脅迫后,冬小麥(TriticumaestivumL.)ABA的含量快速增加,GA水平升高,隨著冷處理時間延長,植物適應低溫,ABA、CTK和GA的含量降低,SA和JA的含量開始增加[21]。然而,Du等[22]研究發現,水稻經過1 d的低溫處理后,IAA含量連續5 d保持在較高水平。本研究結果與之一致。IAA的濃度易受低溫脅迫的影響,其差異取決于植物種類、發育背景和生理環境。例如,低溫脅迫后冬小麥中IAA含量增加,而春小麥中IAA含量保持不變[23]。低溫處理小麥后,在小麥冠狀組織中IAA濃度顯著增加,而小麥葉片中的IAA濃度卻無明顯變化[24]。本研究中,龍粳31內源激素受低溫影響的敏感程度要低于9311,這與表型和生理指標方面的結論相對應。
綜上所述,通過對龍粳31和9311耐冷特性鑒定及BRs調節其耐冷表型和生理生化指標變化的研究,初步證明龍粳31具備更強的芽期和苗期的低溫抗性,同時具備更好的耐冷生理基礎。BRs處理,可對水稻芽期和苗期的多個表型指標產生傾向于耐冷性提高的正向影響,對龍粳31的影響可能更顯著。但是,水稻耐冷性是一個復雜的數量性狀,關于寒地水稻耐冷性與激素之間的關系需要進一步研究,外源施加BRs會對大部分植物激素含量產生影響,但BRs與水稻內源激素之間的關系仍需要進一步探討。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
