連作年限影響芹菜根際土壤微生物群落結構及功能類群*
2023-05-26 10:18:38吳宏亮馮海萍張海宇康建宏
中國農業氣象 2023年5期
劉 素, 吳宏亮**,陳 倬,馮海萍,張海宇,康建宏
連作年限影響芹菜根際土壤微生物群落結構及功能類群*
劉 素1, 吳宏亮1**,陳 倬1,馮海萍2,張海宇1,康建宏1
(1.寧夏大學農學院,銀川 750021;2.寧夏農林科學院園藝研究所,銀川 750000)
芹菜是寧夏冷涼區特色蔬菜,大規模種植不可避免地導致連作障礙。為了揭示芹菜根際土壤微生物的多樣性特征和群落結構隨種植年限變化的差異,本研究采用Illumina Novaseq?PE250測序平臺對芹菜種植一年(CA)、連作兩年(CB)和連作三年(CC)時根際土樣的16S V3V4區和ITS1區進行雙末端Paired-end測序,并進行生物信息學分析。結果表明,土壤細菌多樣性隨連作年限延長呈逐漸降低趨勢,Chao 1指數第一年8509.41,第三年降低至6314.09。CA香農指數最高,為11.48,CC最低,僅11.08。隨著連作年限的增加,土壤細菌中的δ-變形菌綱(Deltaproteobacteria)和芽單胞菌綱(Gemmatimonadetes)等優勢菌綱及其中的羅庫菌屬()、芽球菌屬()、紅桿菌屬()等有益菌豐度減少。土壤真菌的多樣性指數、豐富度指數等均隨連作年限增加呈先下降后上升趨勢,表現為CA、CB的Chao 1指數分別為505.44和264.66,而CC為443.92,CA、CB的香農指數分別為5.98和4.93,而CC為5.61,且連作年限間均存在顯著差異。同時,土壤真菌中的糞殼菌綱(Sordariomycetes)和瓶毛殼屬()、黑孢殼屬()等優勢菌綱屬具有顯著變化差異。微生物主坐標成分分析結果表明,CC與其他處理間存在強異質性。PICRUSt2功能預測分析表明,芹菜根際土壤細菌主要有7大類一級功能層,氨基酸生物合成相對豐度占比最高,真菌有5大類一級功能層,核苷和核苷酸生物合成相對豐度占比最高,同時芹菜連作不同年限時多條代謝通路間具有極顯著差異。綜上研究結果,芹菜連作會降低根際土壤菌群的豐富度及多樣性,改變菌群結構,導致根際微生態環境失衡,從而引發相關連作障礙病害。其中,以芹菜連作三年時微生物各項指數變幅最大,故建議寧夏南部山區芹菜產業將連作不超過兩年作為田間種植模式。
芹菜;連作年限;根際土壤微生物;高通量測序;PICRUSt2功能預測
芹菜(L.)為傘形科植物[1?2],富含大量芹菜素[3],是一種優質、高產、高營養的蔬菜,但芹菜不耐連作。寧夏西吉縣地處海拔1688-2633m,為典型的溫帶大陸性季風氣候,海拔高、晝夜溫差大、氣候冷涼、光熱資源豐富,為當地芹菜產業的發展創造了有利條件[4],素來具有“中國西芹之鄉”美稱。但由于當地耕地面積的有限,加之種植條件的制約及經濟利益驅動等因素,致使芹菜連作成為農戶在農業生產中常采用的種植方式。而長期連作會引發作物嚴重的病害問題,如根莖腐病、放線蟲害等,使其品質及產量顯著下降,嚴重影響寧夏南部山區芹菜產業的健康可持續發展。
根際土壤微生物作為土壤生態系統的重要組成部分[5],與土壤環境中的能量流動及物質轉化息息相關,并參與到許多重要的生化反應中[6]。而連作會對根際土壤微生物群落結構造成破壞,抑制有益生物的存活,同時促進病原菌的生長,加劇土傳病害的發生,進而對作物的生長造成影響,導致減產[7]。高葦等[8]研究表明,芹菜連作后形成的根腐病會嚴重影響土壤真菌、細菌的種類和數量。鄭立偉等[9]認為,甜瓜連作年限的增加致使土壤細菌多樣性降低,細菌中多數優勢菌門的有益菌豐度減少,土壤真菌的豐度增加。岳思君等[9]通過冗余分析表明,引起硒砂瓜連作障礙發生的主要原因是隨著連作時間的增加,放線菌門、變形菌門等有益微生物豐度下降,而與土壤理化性質變化呈不顯著關系。總之,連作種植在一定程度上會引起土壤微生物多樣性的改變,但有關芹菜連作對微生物多樣性影響尚缺乏系統研究,特別是對不同連作年限的芹菜種植土壤的微生物群落結構分析未見報道。因此,從土壤根際微生物菌群的群落結構穩定性與功能多樣性的微生態系統角度研究連作障礙,對作物生產具有重要的理論和實踐意義[10]。本研究通過Illumina Novaseq-PE250高通量測序技術對芹菜連作不同年限時的根際土壤菌群結構與多樣性進行分析,旨在探明芹菜連作時根際土壤微生物群落變化情況,以期為探究造成芹菜連作障礙的核心病菌提供土壤環境微生物生態層次的理論依據。
1 材料與方法
1.1 土樣收集
樣品于2020年8月中旬,在寧夏回族自治區固原市西吉縣隆堡村(N 35°81′E106°11′)不同種植年限芹菜地芹菜收獲期采集,分別命名為芹菜一年根際土壤(用CA表示);芹菜連作兩年根際土壤(用CB表示);芹菜連作三年根際土壤(用CC表示)。選擇采樣單元相對中心位置的典型地塊用五點取樣法收集樣品,用鏟子去除地表植被和其他雜質,在地面距根部5cm處挖出植株,再使用抖落法去除未富集根系土壤,將主根與毛根附著發達區域300~500g土壤作為根際土壤。五處根際土壤充分混勻后,挑出根系、秸稈、石塊和蟲體等雜物,在田間用四分法棄去多余部分,最后每個處理保留1kg左右,裝入17cm×25cm自封袋,掛好內外標簽,置入簡易冷藏箱進行冷藏,隨后將樣品凍存于-80℃備用,準備送樣檢測,每組樣品設4個重復,樣品測序與分析委托上海某公司完成。
1.2 觀測項目及分析
1.2.1 基因組DNA的提取和PCR擴增
使用OMEGA Soil DNA Kit試劑盒通過間接法提取樣品根際土壤中的宏基因組DNA,再將土壤樣品中的DNA進行0.8%瓊脂糖凝膠電泳,進行分子大小判斷,利用紫外分光光度計對DNA進行定量。選擇用于細菌鑒定的16S rRNA V3V4區引物,真菌鑒定的ITS 1區引物進行 PCR 擴增,以通過瓊脂糖凝膠電泳檢測且檢測合格的PCR擴增產物為模板進行測序文庫的制備,最后將合格文庫在Novaseq-PE250測序平臺上機測序。
1.2.2 分析流程
(1)對高通量測序的原始下機數據根據序列質量進行初步篩查,對問題樣本進行重測、補測。
(2)通過質量初篩的原始序列按照index和Barcode信息,進行文庫和樣本劃分,并去除barcode序列。
(3)按照QIIME2 dada2[11?12]分析流程進行OTU聚類。
(4)對各樣本(組)在不同物種分類學水平的具體組成進行展示,了解整體概況。
(5)在ASV/OTU層面,計算各樣本的距離矩陣,并通過多種非監督的排序、聚類手段,結合相應統計學檢驗方法,衡量不同樣本(組)間的Beta多樣性差異及差異顯著性。
(6)在物種分類學組成層面,通過各種非監督、監督的排序、聚類和建模手段,結合相應統計學檢驗方法,進一步衡量不同樣本(組)間的物種豐度組成差異,并嘗試尋找標志物種。
(7)根據16S rRNA和ITS基因測序結果,做預測樣本的PICRUSt2功能預測分析[13]菌群代謝功能。
1.3 統計分析
用SPSS 22.0統計分析軟件對數據進行單因素方差分析,處理間差異顯著性檢驗使用鄧肯法(Duncan,P<0.05)。微生物群落結構變化分析圖和功能預測圖形使用QIIME2 軟件和R語言繪制。
2 結果與分析
2.1 連作年限對芹菜根區土壤微生物群落多樣性指數的影響
不同連作年限田間芹菜根區土壤樣品的細菌群落α多樣性指數觀測結果見表1。由表可見,Good’s coverage覆蓋率為95%以上,即該土壤深度已經基本覆蓋樣品中所有的物種。對3個不同連作年限土壤的細菌α多樣性進行統計分析可見,隨著連作時間延長,土壤的Chao1指數、Shannon指數和Pielou’s evenness指數均有一定程度的降低。同時,CC處理與CA和CB處理的土壤中均呈現指標間顯著差異,說明土壤進行芹菜的多年連續種植會降低其作物根際土壤細菌群落的豐富度及多樣性。
真菌群落α多樣性指數結果表明(表1),各樣品文庫的覆蓋率為100%,觀測到樣品中序列沒有被測出的概率很低。各處理多樣性指數(Shannon指數)、豐富度指數(Chao1指數)和均勻度指數(Pielou’s evenness指數)變化趨勢一致,均隨連作年限增加呈下降后再回升的趨勢,組間呈現顯著性差異。其中CA數值最高,分別為505.44、5.98和0.6671,CB數值最低,分別為264.66、4.93和0.6131,說明芹菜的種植年限也會影響其根部土壤真菌菌群群落,連作年限不同,多樣性、豐富度和均勻度不同。
2.2 連作年限對芹菜根區土壤細菌群落的影響
2.2.1 群落組成及豐度分析
選取豐度排名前8的物種,生成柱形圖。由圖1可見,主要細菌優勢菌綱γ-變形菌綱(Gammaproteo- bacteria)、α-變形菌綱(Alphaproteobacteria)、放線菌綱(Actinobacteria)和擬桿菌綱(Bacteroidia)的相對豐度變化趨勢一致,由高到低排序為CC>CB>CA。其中,CC土壤中的上述4個菌綱豐度較CA的增幅分別為71.07%、44.40%、67.52%和182.52%。δ-變形菌綱(Deltaproteobacteria)、Subgroup_6菌綱、芽單胞菌綱(Gemmatimonadetes)和嗜熱性菌綱(Thermoleophilia)的相對豐度隨連作年限增加而呈現下降趨勢。

表1 三種連作處理芹菜根際土壤樣品中微生物多樣性指數比較(平均值±均方差)
注:CA為芹菜種植一年根際土壤,前茬是休閑土壤,CB為芹菜連作兩年根際土壤,CC為芹菜連作三年根際土壤。小寫字母表示處理間在0.05水平上的差異顯著性。下同。
Note: CA is the rhizosphere soil for celery planting for one year, which previous crop is the fallow soil, CB is the rhizosphere soil for celery continuous cropping for two years, and CC is the rhizosphere soil for celery continuous cropping for three years. The lower case letters indicate the significant difference between treatments at 0.05 level. The same as below.
選取屬水平排名前 35 的屬進行聚類分析,繪制熱圖。由圖2可見,在CA處理中,細菌相對豐度占主要優勢的(相對豐度>1%)有羅庫菌屬()、菌屬、芽球菌屬()、菌屬和紅桿菌屬()共5個屬。在CB處理中,纖維弧菌屬()和溶桿菌屬()兩種菌屬的相對豐度相較CA的土壤有明顯升高。而CC處理中,以上各屬的相對豐度明顯降低,而黃桿菌屬()、菌屬、假單胞菌屬()、異根瘤菌屬-新根瘤菌屬-副根瘤菌屬-根瘤菌屬()、砂單胞菌屬()、氣微菌屬()和類諾卡氏菌屬()的豐度較高。以上結果說明,芹菜連作不同年限時土壤根際菌群在屬分類水平上存在顯著差異。
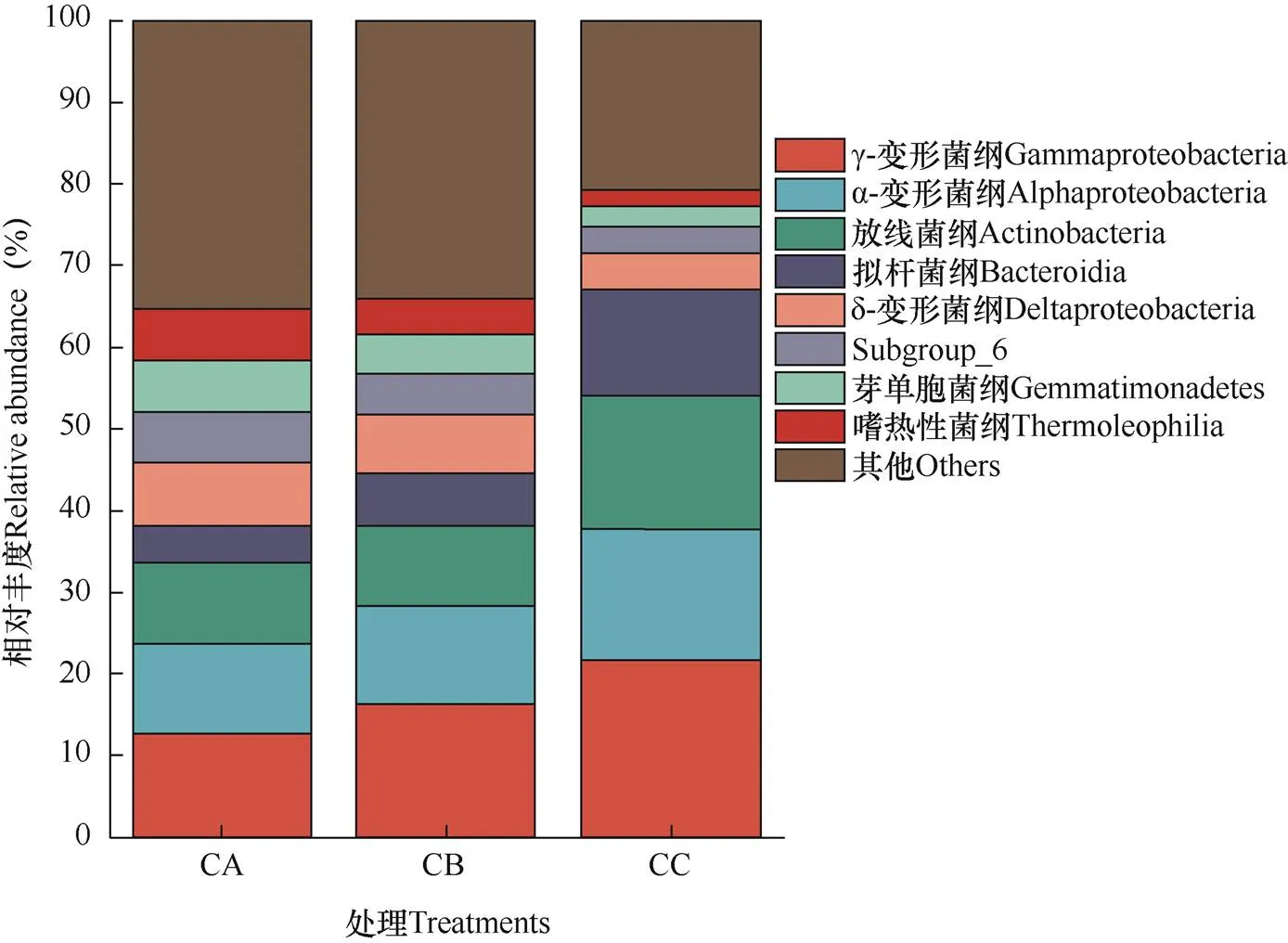
圖1 三種連作處理中芹菜根際土壤樣品在細菌綱水平上的組成和相對豐度

圖2 三種連作處理中芹菜根際土壤樣品在細菌屬水平上的物種豐度聚類熱圖
圖3表明,PCo1和PCo2能解釋3組樣品中64.5%的樣品信息。其中,CA和CB分布在PCo1負值區域,說明細菌群落結構相似性較高,CC分布在正值區域,距離較遠,說明相似性較低且組間差異大于組內差異,進一步表明芹菜連作不同年限時其根際土壤細菌菌群結構存在顯著差異。
2.2.2 PICRUSt2細菌菌群功能預測分析
2.2.2.1 代謝通路統計
基于MetaCyc數據庫對芹菜連作土壤進行細菌菌群功能預測分析(圖4),結果表明,第一層次代謝通路共有7大類,每一類代謝通路又被進一步劃分為多個等級。在芹菜根際土壤細菌的一級功能層中,生物合成為其主要組成。同時預測基因的第二層次代謝通路分析結果表明細菌菌群在多方面具有功能潛能,但只有在9種生物合成方面其相對豐度大于18000,占比最高的為氨基酸生物合成。

圖3 三種連作處理中芹菜根際土壤樣品細菌群落組成主坐標分析

圖4 基于MetaCyc數據庫對芹菜根際土壤不同細菌菌群PICRUSt2功能預測結果
2.2.2.2 代謝通路差異分析
在獲得代謝通路的豐度數據后,對CA與CB、CB與CC、CA與CC分別進行組間代謝通路差異分析。在CA和CB中,以CA為對照組,CB為上調組,圖5a顯示,CB相對CA有34條代謝通路上調,其中25條通路之間具有極顯著差異(P<0.001),分別是PWY-6565通路superpathway of polyamine biosynthesis III和ECASYN-PWY通路enterobacterial common antigen biosynthesis等。在CB和CC中(圖5b),以CB為對照組、CC為上調組進行代謝通路的差異性分析,結果表明,CC相對CB有24條通路上調,26條通路下調,其中38條通路之間呈現極顯著差異(P<0.001),分別是PWY-7385通路1,3-propanediol biosynthesis(engineered)和PWY- 7198通路pyrimidine deoxyribonucleotides de novo biosynthesis IV等。在CA和CC中(圖5c),CA為對照組,CC為上調組,結果表明,CC相對CA有27條通路上調,23條通路下調,其中37條通路之間呈現極顯著差異(P<0.001),分別是PWY-5499通路Pvitamin B6 degradation和PWY-7209通路superpathway of pyrimidine ribonucleosides degradation等。結果說明芹菜的連作年限對土壤微生物代謝有十分重要的影響,連作年限越長通路上下調變化越明顯,通路之間差異越顯著。
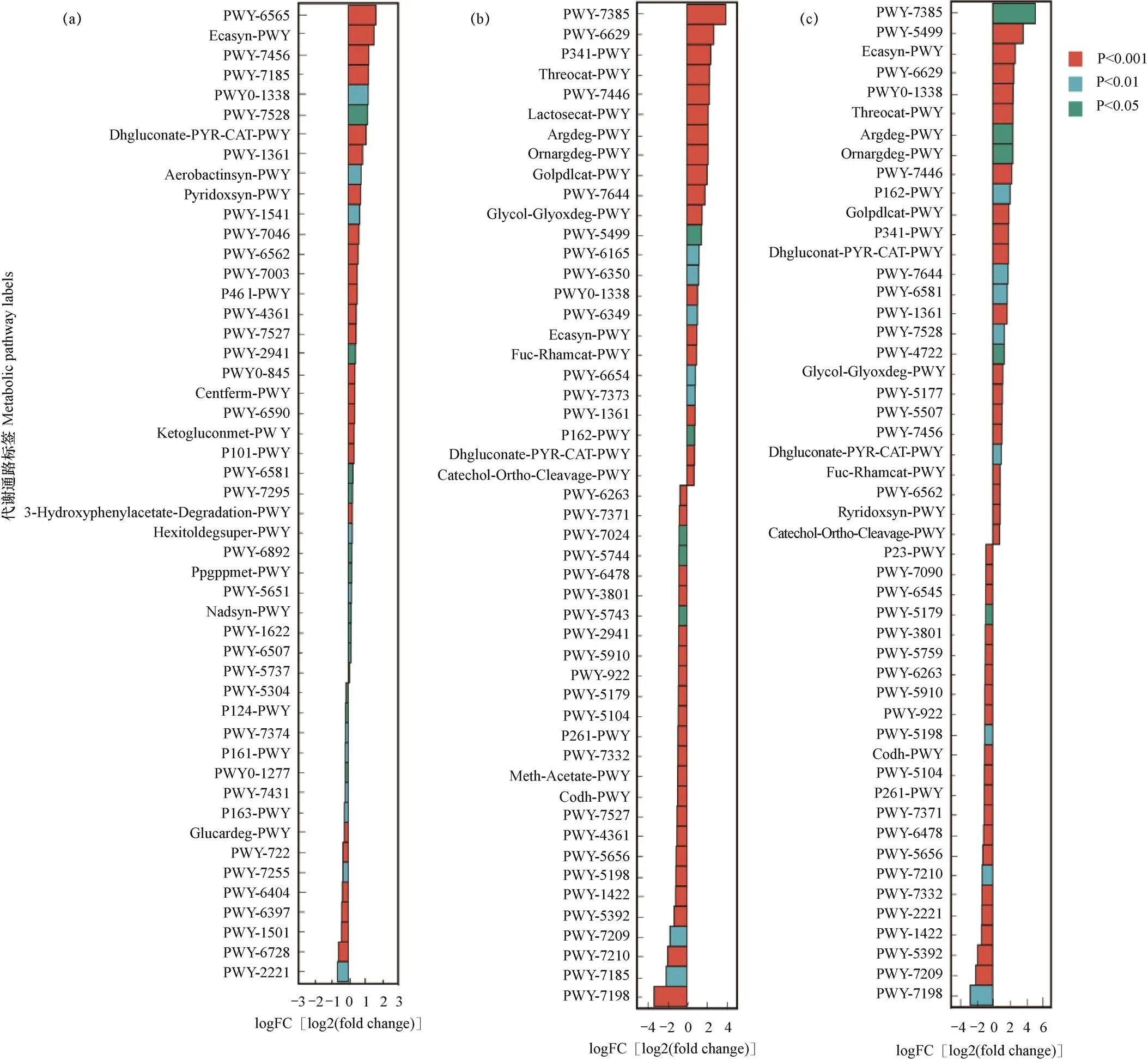
圖5 CA與CB(a)、CB與CC(b)以及CA與CC(c)處理中土壤細菌菌群代謝通路的差異分析
注:logFC[log2(fold change)]為正值代表上調組相對對照組上調,負值為下調。下同。
Note: A positive value of logFC[log2(fold change)] represents upregulation in the upregulation group relative to the control group, and a negative value is downregulation. The same as below.
2.3 連作年限對芹菜根區土壤真菌群落的影響
2.3.1 群落組成及豐度分析
選取豐度排名前8的物種,生成柱形圖。由圖6可見,主要真菌優勢菌綱糞殼菌綱(Sordariomycetes)、銀耳綱(Tremellomycetes)、盤菌綱(Pezizomycetes)、錘舌菌綱(Leotiomycetes)和傘菌綱(Agaricomycetes)的相對豐度在三組樣本中均呈先下降后上升趨勢,由大到小排序為CC>CA>CB。其中,糞殼菌綱(Sordariomycetes)具有顯著變化差異,相對豐度分別為55.56%、51.98%和44.52%。座囊菌綱(Dothideomycetes)和被孢霉綱(Mortierellomycetes)的變化趨勢一致,由高到低分別為CA>CB>CC。其中,座囊菌綱(Dothideom- ycetes)的相對豐度百分比依次為15.56%、13.31%和10.72%。
選取屬水平排名前20的屬進行聚類,繪制熱圖。由圖7可見,CA土壤中,前20中相對豐度大于1% 的有7個屬,分別為菌屬、刺毛四枝孢菌屬()、彎孢菌屬()、炭疽菌屬()、被孢霉菌屬()、赤霉菌屬()和zyma菌屬。CB中以上菌屬的相對豐度呈下降趨勢,而鏈格孢菌屬()的相對豐度大于1%。CC中6個屬的相對豐度與CA和CB差異顯著,分別為瓶毛殼屬()、黑孢殼屬()、木拉克酵母屬()、絲殼屬()、假裸囊菌屬()和土赤殼屬()。
圖8表明,PCo1和PCo2能解釋3組樣品82.3%的菌群多樣性。其中,CA主要分布于PCo1的負端區域,CB分布于PCo1的0值附近,CC位于PCo1的正端區域,3個樣品組間相似性較小,差異顯著,說明芹菜連作后根際土壤真菌群落譜系間發生了改變。
2.3.2 PICRUSt2真菌菌群功能預測分析
2.3.2.1 代謝通路統計
基于MetaCyc數據庫的預測分析見圖9。由圖可見,芹菜根際土壤真菌菌群的一級功能層包括5大類,其中生物合成、前體代謝和能量生成功能層的真菌菌群相對豐度較高。在第二層次代謝通路中,9種生物合成方面和2種前體代謝和能量生成方面真菌菌群的相對豐度大于3000,其中,占比最高的為核苷和核苷酸生物合成。

圖6 三種連作處理中芹菜根際土壤樣品在真菌綱水平上的組成和相對豐度
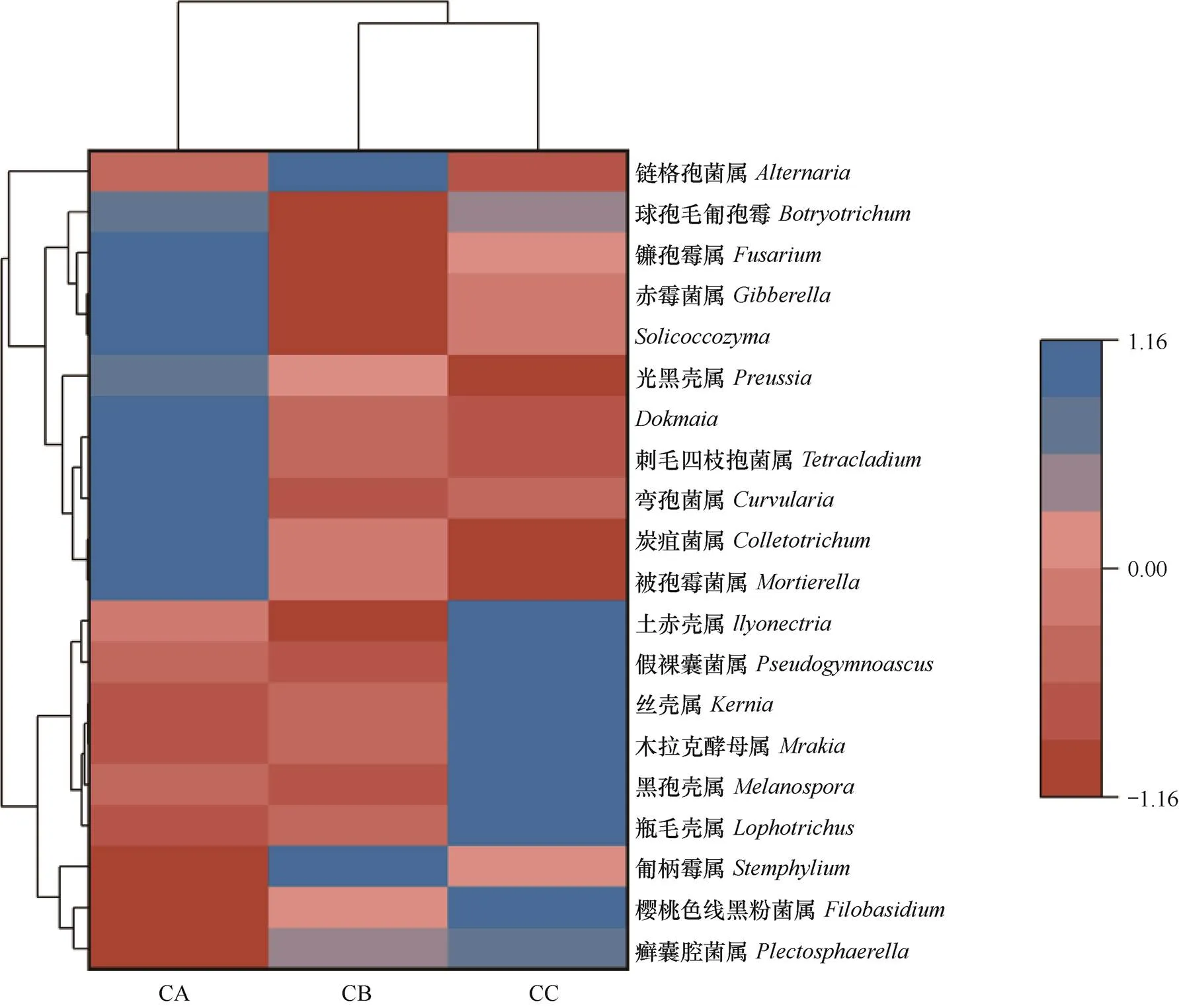
圖7 三種連作處理芹菜根際土壤樣品在真菌屬水平的物種豐度聚類熱圖
2.3.2.2 代謝通路差異分析
代謝通路差異分析可以找出組間具有顯著差異的代謝通路(圖10)。在CA和CB的土壤樣品中(圖10a),以CA為對照組,CB為上調組,結果顯示,CB相對CA有6條代謝通路上調,10條通路下調,其中4條通路之間具有極顯著差異(P<0.001),下調通路中PWY-7409通路phospholipid remodeling和P221-PWY通路octane oxidation等變化幅度較大。CB和CC中(圖10b)以CB為對照組、CC為上調組進行代謝通路的差異性分析,結果表明,CC相對CB有4條通路上調,14條通路下調,其中9條通路之間呈現極顯著差異(P<0.001),上調通路中P221-PWY通路octane oxidation和SO4ASSIM- PWY通路sulfate reduction I (assimilatory)等具較大變幅。在CA和CC的土壤樣品中(圖10c),CA為對照組,CC為上調組,結果表明,CC相對CA有14條通路均下調,其中3條通路之間呈現極顯著差異(P<0.001),分別是PWY4FS-8通路phosphatidylglycerol biosynthesis II (non-plastidic)和PWY4FS-7通路phosphatidylglycerol biosynthesis I (plastidic)等。結果進一步說明芹菜的連作年限對其微生物代謝有十分重要的影響,連作年限越長通路上下調變化越明顯,通路之間差異越顯著。

圖10 CA與CB(a)、CB與CC(b)以及CA與CC(c)處理中土壤真菌菌群代謝通路的差異分析
3 討論與結論
3.1 討論
本研究采用Illumina平臺對群落DNA片段進行雙端(Paired-end)測序,通過將16S rRNA的V3?V4區和ITS V1區測序,結果表明,序列覆蓋率在95%及以上的測序結果可以確切反映土壤環境中的細菌群落結構。
根際土壤是植物能量和物質代謝最活躍的部位之一[14],該微環境代表了植物-微生物和微生物-微生物相互作用的主要生態位,其中細菌在根際土壤生態系統中扮演著重要角色,對于植物生長發育和環境適應至關重要[15?16]。例如能直接參與土壤微生態物質循環和能量交換過程的固氮菌和纖維素分解菌,其分別在土壤氮元素循環和碳元素循環中起到重大作用[17?18],能將亞硝酸根氧化成硝酸根離子的硝化細菌,是幫助作物特別是旱作物順利吸收利用所需氮素的主要細菌類群[19],能作為生防菌的芽孢桿菌[20]、假單胞桿菌[21]和巴氏桿菌[22],可通過競爭作用和拮抗作用誘導作物抗性并促進作物生長,間接提高作物抗性等一種或多種聯合機制來抑制引起植物病害的某些病原菌的存活和活動[23?24]。因此,土壤菌群的結構和多樣性對改善土壤質量和作物健康起著決定性的作用。
本研究通過對不同年限連作的芹菜根際土壤細菌進行比較分析,探究作物不同種植年限與其根際土壤菌群結構及多樣性之間的關系。結果表明,芹菜連作兩年時,纖維素分解性細菌的纖維弧菌屬()相對豐度有所增加,它是具有解磷、固氮等促生功能的有益內生菌屬[25]。同時黃單胞科的溶桿菌屬()也比一年生芹菜的溶桿菌屬相對豐度較高,該菌是一種重要的生防革蘭氏陰性細菌,能產生多種胞外水解酶,包括幾丁質酶、纖維素酶和蛋白酶,以及小分子次生抗菌代謝產物,對植物病原真菌、卵菌和陽性細菌均有較好的抗菌活性[26]。而在芹菜連作三年的根際土壤中,以上兩種有益細菌菌屬豐度反而降低,且己科河菌門的病原菌屬豐度急劇減少,黃桿菌門的屬相對豐度增加。有研究報道,羅庫細菌可以構成一種單物理、植物水平譜系,對廣泛的化學刺激做出反應,同時編碼多種碳傳遞蛋白,包括專門生產脂質、肽和糖的蛋白質[27],而黃桿菌屬為條件致病菌,在高鹽堿土壤中會明顯增多,與作物連作模式中植物病害的發生率升高呈顯著相關性[28?29]。這一結果說明,芹菜種植連作年限越長,其根際土壤中原有的(具有潛在的纖維素降解功能[30])(根際促生菌[31])等功能性細菌量都會持續下降,對土壤環境中的土壤營養元素循環、植物對營養元素的吸收利用產生一定負面作用,從而影響植物的生長發育,降低作物產量與抗病能力。
對土壤真菌群落結構變化分析發現,芹菜連作三年時,瓶毛殼菌屬(s)、黑孢殼屬()、木拉克酵母菌屬() 、絲殼菌屬()、假裸囊菌屬()和土赤殼菌屬()的相對豐度急劇增加。其中,Farag Mf[32]通過真菌的形態學特征、病癥癥狀和致病性試驗,將確定為番茄粉紅色根腐病的致病因子。同時,張進強等[33]對天麻連作土壤研究表明,的豐富度與天麻土傳真菌病害發生率呈顯著正相關。當地芹菜連作障礙主要表現為易感染根腐病,有可能是因為以黑孢殼屬和土赤殼屬真菌為主,其他侵襲性真菌為輔過多積累導致,但本試驗結果還需在田間進一步驗證,以具體明確影響連作植物生長的微生物種群,為連作土壤微生物菌群的定向改造提供更加充分的理論依據。
PICRUSt2軟件是基于PICRUSt軟件升級后的細菌群落功能預測平臺,在土壤微生物群落功能預測方面多見應用[34],它可以適配OTU序列或ASV序列和豐度數據,將微生物的變化情況和生物功能聯系起來[35]。王峰等[36]通過對茶園土壤細菌群落進行PICRUSt2功能預測,得出其在代謝、遺傳信息處理和環境信息方面功能活躍。呂錫斌等[37]運用PICRUSt2預測軟件分析得出,赤水河流域微生物群落的功能主要涉及代謝等46個子功能。本研究通過將芹菜根際土壤的16S rRNA序列和ITS序列進行PICRUSt2功能預測分析,發現土壤細菌共涉及7個主代謝通路,真菌共涉及5個主代謝通路,其子通路均表現出功能上的豐富性,同時通路之間也表現出極顯著差異。但鑒于PICRUSt2功能預測分析具有一定的局限性,所以后續研究需要結合宏基因組測序等技術進一步分析。
3.2 結論
芹菜連作時其根際土壤的微生物群落結構與多樣性之間存在顯著差異。連作年限增加,益生菌比例緩慢下降,致病病原菌比例呈先下降后急劇增長趨勢,此微生物群落結構的變化不利于作物的生長發育,最終導致芹菜出現嚴重的連作問題。因此,本研究認為芹菜連作兩年是寧夏南部山區較好的田間種植方式,其連作障礙問題可能是因為連作兩年后,根際土壤部分致病真菌菌群如黑孢殼屬、土赤殼屬真菌豐度急劇增加、結構劇烈變化引起。
[1] 汪勁武.傘形科植物觀(上)[J].植物雜志, 1989(6):22-24.
Wang J W.Apiaceae plant view(Part I)[J].Chinese Journal of Botany,1989(6):22-24.(in Chinese)
[2] 汪勁武.傘形科植物觀(下)[J].植物雜志, 1990(1):30-32.
Wang J W.Apiaceae plant view (Part II)[J].Chinese Journal of Botany, 1990(1):30-32.(in Chinese)
[3] 李榮華,夏薇,吳坤.蔬菜中芹菜素高效液相色譜測定[J].中國公共衛生, 2007(7):894-895.
Li R H,Xia W,Wu K.Determination of apigenin in vegetables by high performance liquid chromatography[J]. Chinese Journal of Public Health,2007(7):894-895.(in Chinese)
[4] 陳麗生.西吉縣特色冷涼蔬菜產業發展現狀分析[J].新農業,2019(15):84-85.
Chen L S.Analysis of the current situation of the development of special cold vegetable industry in Xiji county[J].New Agriculture,2019(15):84-85.(in Chinese)
[5] Yang R P,Mo Y L,Liu C M,et al.The effects of cattle manure and garlic rotation on soil under continuous cropping of watermelon(L)[J].PloS One, 2016,11(6):e0156515.
[6] Li W,Liu Q,Chen P.Effect of long-term continuous cropping of strawberry on soil bacterial community structure and diversity[J].Journal of Integrative Agriculture, 2018,17(11):2570-2582.
[7] Dong L,Xu J,Feng G,et al.Soil bacterial and fungal community dynamics in relation todeath rate in a continuous cropping system[J].Scientific Reports,2016,6(1):31802.
[8] 高葦,吳峰,王勇,等.棉隆與生物有機肥協同防治芹菜根腐病及其對根際土壤微生物數量的影響[J].中國農學通報,2018,34(2):65-68.
Gao W,Wu F,Wang Y,et al.Synergetic effect of dazomet and biological fertilizer on celery root rot and soil rhizosphere microorganisms[J].Chinese Agricultural Science Bulletin, 2018,34(2):65-68.(in Chinese)
[9] 鄭立偉,趙陽陽,王一冰,等.不同連作年限甜瓜種植土壤性質和微生物多樣性[J].微生物學通報,2022,49(1):101- 114.
Zheng L W,Zhao Y Y,Wang Y B,et al.Soil properties and microbial diversity in the muskmelon fields after continuous cropping for different years[J].Microbiology China, 2022, 49(1):101-114.(in Chinese)
[10] 岳思君,馮翠娥,楊彥研,等.不同連作年限硒砂瓜土壤細菌群落結構特征[J].干旱地區農業研究,2020,38(1):230- 236.
Yue S J,Feng C E,Yang Y Y,et al.Analysis of microbial community structure and diversity in selenium-sand melon soil under different continuous cropping years[J].Agricultural Research in the Arid Areas, 2020,38(1):230-236.(in Chinese)
[11] Callahan B J,Mcmurdie P J,Rosen M J,et al.DADA2:High resolution sample inference from Illumina amplicon data[J]. Nature Methods,2016,13(7):581-583.
[12] Bokulich N A,Kaehler B D,Ram R J,et al.Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2's q2-feature-classifier plugin[J]. Microbiome, 2018,6(1):90.
[13] Gavin M D,Vincent J M,Jesse R Z,et al.PICRUSt2 for prediction of metagenome functions[J].Nature Biotechnology: the Science and Business of Biotechnology,2020,38(6): 685-688.
[14] 李夢潔,李占彪,周思含,等.植物根際微生物調控根系構型研究[J].中國農業氣象, 2021,42(11):895-904.
Li M J,Li Z B,Zhou S H,et al.Advances in the root system architecture regulated by plant rhizosphere microorganisms[J]. Chinese Journal of Agrometeorology,2021,42(11):895-904. (in Chinese)
[15] Wang X,Feng H,Wang Y,et al.Mycorrhizal symbiosis modulates the rhizosphere microbiota to promote rhizobia- legume symbiosis[J].Molecular Plant,2021,14(3):503-516.
[16] 岳衡,李閃閃,段雅欣,等.深松耕對寧南山區馬鈴薯田土壤細菌多樣性的影響[J].中國農業氣象,2021,42 (12):998- 1008.
Yue H,Li S S,Duan Y X,et al.Effect of subsoil plowing on soil bacterial diversity in potato fields in mountainous areas of southern Ningxia[J].Chinese Journal of Agrometeorology, 2021,42(12):998-1008.(in Chinese)
[17] 劉亞軍,馬琨,李越,等.馬鈴薯間作栽培對土壤微生物群落結構與功能的影響[J].核農學報, 2018,32(6):1186- 1194.
Liu Y J,Ma K,Li Y,et al.Effect of different Intercropping cultivation patterns of potato on the structure and function of soil microorganism community[J].Journal of Nuclear Agricultural Sciences,2018,32(6):1186-1194.(in Chinese)
[18] 殷繼忠,李亮,接偉光,等.連作對大豆根際土壤細菌菌群結構的影響[J].生物技術通報, 2018,34(1):230-238.
Yin J Z,Li L,Jie W G,et al.Effects of continuous cropping on bacterial flora structure in soybean rhizosphere soil[J]. Biotechnology Bulletin, 2018,34(1):230-238.(in Chinese)
[19] 張星,林煒鐵,朱雅楠.硝化細菌中亞硝酸鹽氧化還原酶的研究進展[J].微生物學通報, 2008,35(11):1806-1810.
Zhang X,Lin W T,Zhu Y N.Research progress of nitrite oxidoreductase in nitrobacteria[J].Microbiology China,2008, 35(11):1806-1810.(in Chinese)
[20] 王蕊,王騰,李二峰.生防芽孢桿菌在植物病害領域的研究進展[J].天津農學院學報,2021,28(4):71-77.
Wang R,Wang T,Li E F.Research advances of biocontrol Bacillus in the field of plant disease[J].Journal of Tianjin Agricultural University,2021,28(4):71-77.
[21] 高蕓.生防芽孢桿菌及假單胞菌拮抗植物微生物病害研究進展[J].北方園藝,2021(2):131-136.
Gao Y.Research progress in biocontrol of plant microbial diseases by Bacillus and Pseudomonas.[J].Northern Horticulture, 2021(02):131-136.(in Chinese)
[22] 董煒博,石延茂,遲玉成,等.穿刺巴氏桿菌防治植物根結線蟲病的研究現狀及其應用前景[J].中國生物防治, 1999(2):42-46.
Dong W B,Shi Y M,Chi Y C,et al.Status and prospect of biocontrol of plant root-knot nematode diseases by pasteuria penetrans[J].Chinese Journal of Biological Control,1999(2):42-46.(in Chinese)
[23] Li P,Ma L,Feng Y L,et al.Diversity and chemotaxis of soil bacteria with antifungal activity against Fusarium wilt of banana[J].Journal of Industrial Microbiology & Biotechnology, 2012,39(10):1495-1505.
[24] 李俊州,文才藝.植物病害生防細菌研究進展[J].河南農業科學,2015,44(10):1-7.
Li J Z,Wen C Y.Research progress on bacteria for biological control of plant disease[J].Journal of Henan Agricultural Sciences,2015,44(10):1-7.(in Chinese)
[25] 覃仁柳,龐師嬋,唐小付,等.番茄嫁接植株根系內生細菌和真菌群落的組成特征[J].西南農業學報,2021,34: 1062-1072.
Qin R L,Pang S C,Tang X F,et al.Characteristics of endophytic microbial diversity and community structure in roots of grafted tomatoes[J].Southwest China Journal of Agricultural Sciences,2021,34:1062-1072.(in Chinese)
[26] 杭唯.產酶溶桿菌第二信使C-dI-GMP合成相關基因lys385的克隆及功能分析[D].南京:南京農業大學, 2015.
Hang W.Identification and functional characterization of LYS385,a second messenger C-dI-GMP metabolising protein in Lysobacter enzymogenes[D].Nanjing:Nanjing Agricultural University,2015.(in Chinese)
[27] Becraft E D,Woyke T,Jarett J,et al.Rokubacteria:genomic giants among the uncultured bacterial phyla[J].Frontiers in Microbiology,2017(8):2264.
[28] 王文團.耐鹽堿細菌篩選與生物菌肥對鹽堿地作物的影響[D].泰安:山東農業大學,2020.
Wang W T.Screening of saline-tolerant bacteria and effects of biological fertilizer on saline-alkali crops[D].Tai’an Shandong Agricultural University,2020.(in Chinese)
[29] 曹佳.蚯蚓和菌根真菌促進土霉素降解及其修復污染土壤的協同機制[D].北京:中國農業大學,2018.
Cao J.Cooperation between earthworm and AM fungi on the degradation and remediation of oxytetracycline in soil[D].Beijing:China Agricultural University,2018.(in Chinese)
[30] 于竺岑.分離自蘆葦根系一株芽生球菌的新菌鑒定及全基因組分析[D].呼和浩特:內蒙古大學,2021.
Yu Z Z.Identification and genome analysis of a new strain ofisolated from the root of phragmites australis[D].Hohhot:Inner Mongolia University,2021.(in Chinese)
[31] 金桃,馮強,萬景旺,等.五種根際促生菌在改善植物的農藝性狀方面的應用[P].廣東省:CN106472568B,2020-05- 12.
Jin T,Feng Q,Wan J W,et al.Application of five inter-root promoting bacteria in improving agronomic traits of plants[P].Guangdong Province:CN106472568B,2020-05-12. (in Chinese)
[32] Farag Mf. F.First record of Melanospora chionea as a possible cause of pink root rot disease on tomato plants in Egypt[J].Journal of Plant Pathology & Microbiology, 2020 (11):1-8.
[33] 張進強,唐鑫,郭蘭萍,等.天麻連作障礙與土赤殼屬真菌的關聯分析及改善措施[J].中國中藥雜志,2022,47(9): 2296-2303.
Zhang J Q,Tang X,Guo L P,et al.Correlation analysis between continuous cropping obstacle of Gastrodia elata and Ilyonectria fungi and relieving strategy[J].China Journal of Chinese Materia Medica,2022,47(9):2296-2303. (in Chinese)
[34] 宋騰蛟,楊蔣舜,周靜,等.基于Miseq測序技術分析不同元胡產區土壤細菌功能基因組成與差異[J].浙江中醫藥大學學報,2018,42(3):178-186.
Song T J,Yang J S,Zhou J,et al.Analysis of composition and difference of soil bacteria functional genes of in different corydalis Yanhusuo producing areas based on Miseq sequencing technology[J].Journal of Zhejiang Chinese Medical University,2018,42(3):178-186.(in Chinese)
[35] 楊盼,翟亞萍,趙祥,等.叢枝菌根真菌和根瘤菌互作對苜蓿根際土壤細菌群落結構的影響及PICRUSt功能預測分析[J].微生物學通報,2020,47(11):3868-3879.
Yang P,Zhai Y P,Zhao X,et al.Effect of interaction between arbuscular mycorrhizal fungi and Rhizobium on Medicago sativa rhizosphere soil bacterial community structure and PICRUSt functional prediction[J].Microbiology China,2020, 47(11):3868-3879.(in Chinese)
[36] 王峰,陳玉真,吳志丹,等.種植模式和坡位對茶園土壤細菌群落結構及功能類群的影響[J].生態學報,2022(20): 1-18.
Wang F,Chen Y Z,Wu Z D,et al. Effects of planting patterns and slope positions on soil bacterial community structure and functional groups in tea gardens[J].Acta Ecologica Sinica,2022(20):1-18.(in Chinese)
[37] 呂錫斌,吳云成,陳良強,等.赤水河流域浮游細菌群落特征及其與水質的關系[J].環境科學學報,2021,41(11):4596-4605.
Lv X B,Wu Y C,Chen L Q,et al. Characteristics of the bacterioplankton community and their relationships with water quality in Chishui River basin[J].Acta Scientiae Circumstantiae,2021,41(11):4596-4605.(in Chinese)
Continuous Cropping Years Affect the Rhizosphere Soil Microbial Community Structure and Functional Taxa of Celery
LIU Su1, WU Hong-liang1, CHEN Zhuo1, FENG Hai-ping2, ZHANG Hai-yu1, KANG Jian-hong1
(1.School of Agriculture, Ningxia University, Yinchuan 750021, China;2.Institute of Horticulture, Ningxia Academy of Agricultural and Forestry Sciences, Yinchuan 750000)
Celery is a characteristic vegetable in Ningxia cold region, and its large-scale planting inevitably leads to the prominent obstacle of continuous cropping. In order to reveal the diversity characteristics of microbes in celery rhizosphere soil and the difference of community structure with planting years, author used Illumina Novaseq-PE250 sequencing platform to sequence 16S V3V4 region and ITS1 region of celery rhizosphere soil samples after planting for one year (CA), continuous cropping for two years (CB) and continuous cropping for three years (CC), and made bioinformatics analysis.The results showed that the soil bacterial diversity decreased gradually with the extension of continuous cropping years. The Chao 1 index decreased from 8509.41 in the first year to 6314.09 in the third year.The CA Shannon index was the highest at 11.48 and the CC was the lowest at 11.08.The abundance of beneficial bacteria such as Deltaproteobacteria and Gemmatimonadetes in soil bacteria and,andin them decreased. The diversity index and richness index of soil fungi first decreased and then increased with the increase of continuous cropping years, which showed that the Chao 1 index of CA and CB was 505.44 and 264.66, while CC was 443.92, the Shannon index of CA and CB was 5.98, 4.93, and CC was 5.61, and there were significant differences between continuous cropping years. At the same time, there are significant differences among soil fungi, such as,,and other dominant genera. The results of microbial principal coordinate component analysis showed that there was strong heterogeneity between CC and other treatments. The function prediction analysis of PICRUSt2 showed that bacteria in the rhizosphere soil of celery mainly had 7 major categories of primary functional layers, with the highest relative abundance of Amino Acid Biosynthesis, fungi had 5 major categories of primary functional layers, and the highest relative abundance of Nucleoside and Nucleotide Biosynthesis. At the same time, there were significant differences between multiple metabolic pathways in different years of continuous cultivation of celery. To sum up, celery continuous cropping will reduce the richness and diversity of rhizosphere soil flora, change the flora structure, and lead to the imbalance of rhizosphere micro-ecological environment, thus causing related continuous cropping obstacle diseases. Among them, the microbial indices changed the most when celery was continuously cropped for three years. Therefore, it is recommended that the celery industry in the southern mountainous areas of Ningxia should be continuously cropped for no more than two years as a field planting mode.
Celery; Continuous croping year; Rhizosphere soil microorganism; High-throughput sequencing; PICRUSt2 functional prediction
10.3969/j.issn.1000-6362.2023.05.003
收稿日期:2022?05?22
寧夏自然科學基金項目(2021AAC03079);寧夏農業高質量發展和生態保護科技創新示范項目(NGSB?2021?8?06);寧夏自治區重點研發計劃(2020BBF03016);寧夏青年拔尖人才項目
通訊作者:吳宏亮,副教授,主要從事耕作制度、農業生態領域的應用基礎理論和技術研究,E-mail: nxuwu@163.com
劉素,E-mail:nxuliu@163.com
劉素,吳宏亮,陳倬,等.連作年限影響芹菜根際土壤微生物群落結構及功能類群[J].中國農業氣象,2023,44(5):372-385
