脂肪酸通過內質網應激促進脂肪細胞miR-4431表達及釋放的作用及機制研究
2023-06-12 09:23:30李夢環馬丁凌唐意涵袁成鋼王翠喆張君
石河子大學學報(自然科學版) 2023年2期
李夢環,馬丁凌,唐意涵,袁成鋼,王翠喆,張君
(石河子大學醫學院/新疆地方病與民族高發病教育部重點實驗室,新疆 石河子 832000)
糖尿病患病率在世界范圍內呈持續增長狀態,并已成為世界第5位死亡主因[1]。根據《IDF全球糖尿病地圖(第 10 版)》數據顯示,預計到2045年,全球糖尿病患病人數將達7.83億人,其中2型糖尿病(Type2 Diabetes Mellitus, T2DM)約占90%[2]。肥胖是胰島素抵抗(Insulin Resistance, IR)和T2DM發生發展的重要誘因,80%的T2DM患者伴有肥胖[3-4]。盡管多項研究顯示肥胖是產生IR的根源,也是T2DM發生的重要原因,但肥胖誘發T2DM的具體分子機制尚不十分明確。
最近的研究顯示,脂肪組織是循環外泌體微小核糖核酸(microRNAs, miRNAs)的一個重要來源,作為新式脂肪因子miRNAs可調節鄰近或遠處組織中受體細胞的功能[5]。已有研究表明:肥胖個體血清和組織中miRNAs的含量發生變化,可通過影響相關基因的表達誘發IR,在T2DM、心血管疾病和代謝綜合征等發病過程中發揮重要作用[6-9]。作為新型脂肪因子,miRNAs對于預防和治療肥胖相關代謝性疾病具有廣闊的應用前景。課題組前期研究發現:肥胖受試個體及飲食誘導的肥胖小鼠血清中miR-4431含量顯著增加,并且在體外細胞實驗中證實:miR-4431可通過抑制糖代謝關鍵基因TRIP10 和PRKD1的表達,誘發肝細胞IR和糖耐量受損[10]。然而,有關肥胖導致miR-4431含量增加的原因,尚未見文獻報道。
肥胖后,脂肪組織過度蓄積,導致脂解作用增強,進而引起血漿中游離脂肪酸(Free fatty acids,FFAs)含量增加。過量的血清FFAs水平會減少外周葡萄糖攝取,增加肝臟葡萄糖生成,可誘發脂肪組織炎癥和IR[11]。課題組已有研究結果表明:無論在正常體重還是肥胖個體血漿中,長鏈脂肪酸棕櫚酸(Palmitic acid,PA)的含量在34種FFAs中豐度最高,并且肥胖個體血漿中PA含量高于正常體重個體,提示PA可能在肥胖導致相關代謝性疾病的發病過程中發揮重要作用[12]。內質網是絕大部分細胞中的重要細胞器,在維持細胞代謝穩態與生物大分子合成中起著關鍵作用。已有文獻表明,肥胖后脂肪酸和葡萄糖等營養物質過量可破壞內質網功能穩態,導致內質網應激(Endoplasmic Reticulum Stress,ERS)的發生,而這種反應與IR和T2DM的發展過程密切相關[13-17]。肥胖后PA含量增加是否通過內質網應激途徑導致miR-4431的含量增加,尚未見文獻報道。
本研究在收集正常體重及肥胖受試個體腹部脂肪組織,以及構建飲食誘導肥胖小鼠模型的基礎上,比較受試個體血清中miR-4431的含量差異以及腹部脂肪組織中內質網應激標志物PERK、IRE1α、ATF6及miR-4431的表達差異;在體外培養3T3-L1脂肪細胞的基礎上,明確PA處理對內質網應激標志物及miR-4431表達及釋放水平的影響,嘗試闡明肥胖后miR-4431表達及釋放增加的可能原因,為進一步探討肥胖后脂肪組織源性microRNAs表達及釋放增加的分子機制提供理論依據。
1 對象與方法
1.1 研究對象
本研究所涉及的受試個體及實驗動物均獲得石河子大學醫學院第一附屬醫院醫學倫理委員會批準(編號: 2017-049-01;編號: A2017-115-01)。2021年5月~12月,在石河子大學醫學院第一附屬醫院收集12例受試個體腹部脂肪組織樣本,包括6例正常體重及6例肥胖受試個體,所有受試個體均簽署知情同意書;于湖南斯萊克景達實驗動物有限公司購買C57BL/6雄性小鼠12只,分為普通飲食喂養組(n=6)和高脂飲食喂養組(n=6)。
1.2 細胞系及體外培養試劑
本研究所用3T3-L1細胞購自中國科學院典型培養物保藏委員會細胞庫。當25T培養瓶中的細胞生長至80%~100%時即可進行傳代處理。將細胞接種在六孔板中2~3 d后,細胞密度達到100%時,使用細胞分化誘導液Ⅰ(DMEM高糖培養基、10%胎牛血清、10 μg·mL-1胰島素、1 μmol·L-1·L-1地塞米松及 0.5 mmol·L-13-異丁基-1-甲基黃嘌呤)刺激細胞48 h后;使用誘導液Ⅱ(含DMEM高糖培養基、10%胎牛血清、10 μg·mL-1胰島素及1 μmol·L-1·L-1地塞米松)刺激細胞48 h;半換含 DMEM高糖培養基和10%胎牛血清的混合培養基;24 h后,全部更換培養基為DMEM高糖培養基和10%胎牛血清的混合培養基。待細胞成功誘導出大脂滴后,分別使用200 μmol·L-1和500 μmol·L-1PA進行處理。
1.3 棕櫚酸溶液(palmitic acid, PA)配制
使用分析天平稱取棕櫚酸粉末0.025 64 g,溶于1 mL無水乙醇中。將充分溶解的棕櫚酸加至20%無脂肪酸BSA溶液,充分混勻后,使用0.22 μm的濾器進行過濾,分裝備用,-80 ℃保存。
1.4 組織與細胞RNA提取
提前預冷無酶 EP 管、三氯甲烷、異丙醇、75%酒精及低溫超速離心機至 4 ℃。PA處理細胞24 h后,收集細胞培養上清液后,使用1 ml 1×PBS 清洗細胞,Trizol法提取組織及細胞總RNA。
1.5 miRNAs反轉錄及實時定量PCR
實驗所用miRNAs反轉錄及實時定量PCR試劑盒購買于北京天根公司,根據說明書進行操作,引物序列見表1。

表1 PCR引物序列
1.6 肥胖小鼠模型構建
使用高脂飼料(含5%PA的60%fat Kcal%,購自江蘇美迪森生物醫藥有限公司)喂養雄性C57BL/6小鼠11周,構建肥胖小鼠模型;普通飼料(含10%fat Kcal%,購自江蘇美迪森生物醫藥有限公司)喂養雄性C57BL/6小鼠11周,作為普通飲食對照組。
通過比較小鼠體重及血糖水平判定肥胖模型是否構建成功。高脂飲食喂養組小鼠體重大于普通飲食組20%,血糖顯著高于普通飲食組,認為肥胖小鼠模型構建成功。
飼養小鼠11周后,評價小鼠胰島素敏感性及葡萄糖耐受能力;內眥取血,4 000 r·min-1,離心5 min,收集血清;脫頸處死小鼠后,收集小鼠肝臟及各脂肪組織,稱取重量,所有組織-80 ℃保存。
1.7 小鼠葡萄糖耐量(Intraperitoneal Glucose Tolerance Trial, IPGTT)和胰島素敏感性(Insulin Tolerance Test, ITT)實驗
IPGTT實驗前,對小鼠進行饑餓12 h,斷尾檢測血糖,記為0 min。對小鼠進行腹腔注射無水葡萄糖,無水葡萄糖用量:2 g·kg-1(5 g 無水葡萄糖溶于10 mL 生理水,每只小鼠葡萄糖用量=體重(g) ×4 μL);檢測小鼠15、30、45、60、90、120 min血糖水平。
ITT實驗前,對小鼠進行饑餓6 h,斷尾檢測血糖,記為 0 min。ITT 諾和靈30R用量:0.5 IU·kg-1(4 μL的 insulin 溶于 2 mL 生理鹽水,每只小鼠胰島素用量=體重(g) ×2.5 μL);測量小鼠15、30、45、60、90、120 min。
實驗過程中,倘若小鼠出現抽搐無力,立即大量注射葡萄糖(100 μL·只-1),密切關注小鼠狀態 3~5 min,可進行多次注射。
1.8 統計學分析
應用 SPSS 26.0 軟件對各組數據進行統計學分析。服從正態分布或方差齊的兩組計量數據采用t檢驗,非正態分布的兩組計量數據采用非參數秩和檢驗,P<0.05 認為差異具有統計學意義。
2 結果
2.1 肥胖個體脂肪組織中內質網應激標志物及miR-4431表達水平增加
比較受試個體一般資料和生化指標后發現:肥胖個體(n=6,男性3例,女性3例)體重和體質指數顯著高于正常體重(n=6,男性2例,女性4例)個體(P<0.05),結果見表2。
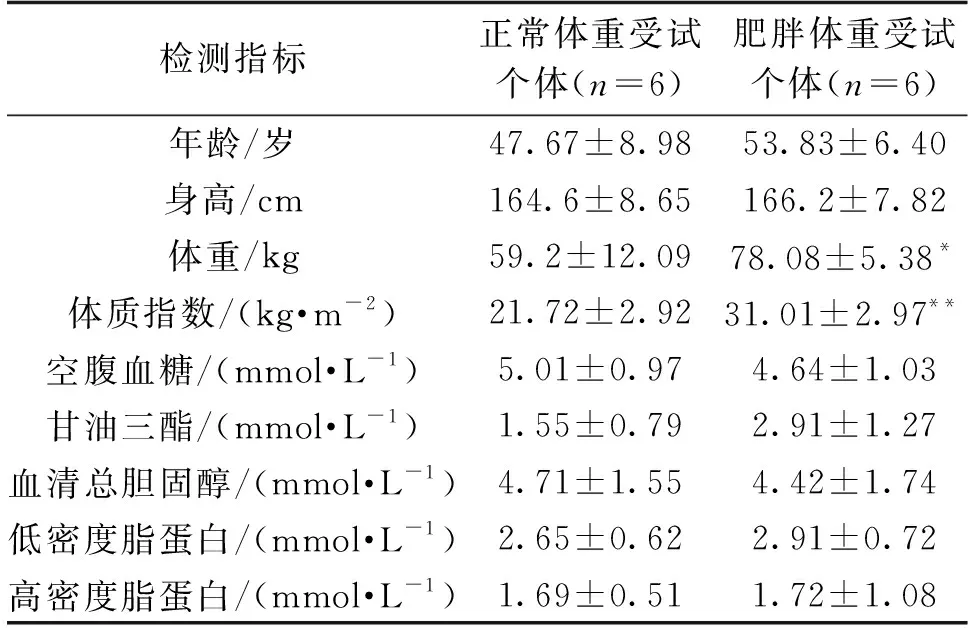
表2 受試個體一般資料
比較受試個體腹部脂肪組織中內質網應激標志物及miR-4431表達水平后發現:與正常體重受試個體相比,肥胖受試個體脂肪組織中內質網應激標志物PERK、IRE1α、ATF6及miR-4431的表達水平均顯著增加,差異有統計學意義(P<0.05,圖1A~圖1D)。相關性分析結果顯示:脂肪組織中miR-4431表達水平與受試個體血糖水平、內質網應激標志物PERK、IRE1α、ATF6正相關(圖1E~圖1H)。

A: PERK;B: IRE1α;C: ATF6;D: miR-4431;E-H:miR-4431與血糖、PERK、IRE1α、ATF6斯皮爾曼相關分析。*P <0.05,**P <0.01, 差異有統計學意義。圖1 內質網應激標志物PERK、IRE1α、ATF6及miR-4431在正常(n=6)和肥胖個體(n=6)脂肪組織中的表達水平
2.2 肥胖小鼠脂肪組織中內質網應激標志物及miR-4431表達增加,血清中miR-4431含量增加
高脂飲食喂養雄性C57BL/6小鼠11周后,小鼠體重顯著高于普通飲食喂養組(大于20%,P<0.05)(圖2A,圖2B)。且小鼠血糖水平較普通飲食組相比顯著升高,葡萄糖耐量及胰島素敏感性明顯受損(P<0.05)(圖2C~圖2E)。提示通過含5%的棕櫚酸高脂飼料喂養11周后,成功構建肥胖小鼠模型。

A:小鼠大體圖片;B:小鼠體重;C:血糖;D:小鼠葡萄糖耐量;E:胰島素敏感性;F-I:附睪脂肪組織中PERK、IRE1α、ATF6、miR-4431的表達水平;J:血清中miR-4431的釋放水平;K-N:miR-4431表達水平與血糖、PERK、IRE1α、ATF6的相關性分析。*P <0.05,**P <0.01, ***P <0.001,差異有統計學意義;斯皮爾曼相關分析,P <0.05,差異有統計學意義。圖2 正常飲食(NCD)以及高脂飲食(HFD)喂養小鼠肥胖相關指標、內質網應激標志物及miR-4431含量的比較
比較小鼠附睪脂肪組織中內質網應激標志物PERK、IRE1α、ATF6、miR-4431的表達水平及血清中miR-4431含量后發現:內質網應激標志物PERK、IRE1α、ATF6及miR-4431在肥胖小鼠附睪脂肪組織中顯著高表達,同時小鼠血清中miR-4431含量顯著增加(P<0.05)(圖2F~圖2J)。脂肪組織miR-4431表達水平與小鼠血糖水平及內質網應激標志物PERK、IRE1α、ATF6的表達水平均顯著正相關(P<0.05)(圖2K~圖2N)。
2.3 PA促進脂肪細胞內質網應激及miR-4431的表達和釋放
體外培養3T3-L1小鼠胚胎成纖維細胞,誘導分化為成熟的脂肪細胞,分別使用200 μmol·L-1和500 μmol·L-1PA處理24 h后,檢測細胞脂質合成能力及內質網應激標志物PERK、IRE1α、ATF6的表達水平,以及miR-4431表達和釋放水平,結果顯示:與NC組相比,200 μmol·L-1及500 μmol·L-1PA處理均可顯著促進細胞脂質合成(圖3A)及內質網應激標志物PERK、IRE1α、ATF6以及miR-4431的表達水平,同時細胞培養上清中miR-4431的含量也顯著升高(P<0.05)(圖3B~圖3F)。


A:油紅O鑒定;B~D:檢測細胞內質網應激標志物PERK、IRE1α、ATF6的表達水平;E~F:miR-4431表達和釋放水平。*P <0.05,**P <0.01, ***P <0.001,差異有統計學意義。圖3 PA處理3T3-L1脂肪細胞對內質網應激和miR-4431表達及釋放的影響
2.4 miR-4431上游轉錄因子調控網絡的預測
運用生物信息學數據庫TransmiR預測miR-4431上游轉錄因子,結果顯示:DDX5可能為miR-4431的上游轉錄因子(圖4A)。進一步使用hTFtarget數據庫預測DDX5的上游轉錄調控網絡后發現。共有205個轉錄因子可調控DDX5,涉及的通路包括細胞增殖與凋亡、免疫調控、神經系統發育等,其中與內質網應激相關因子包括:XBP1、ETS1、ATF2、ATF3、CEBPA、CEBPB等18個轉錄因子其具體功能見表3和圖4B。

A:miR-4431上游轉錄調控因子;B:DDX5上游轉錄調控因子。圖4 生物信息學分析miR-4431及DDX5上游轉錄因子

表3 內質網應激調控的DDX5上游轉錄因子
3 討論
2型糖尿病(T2DM)是世界上患病率增長速度最快的代謝性疾病[18]。調查結果顯示[19-20],目前中國成年人T2DM患病率達11.6%,并且有50.1%處于前糖尿病狀態,已成為全世界T2DM患者最多的國家。肥胖是胰島素抵抗(Insulin Resistance, IR)和T2DM發生的重要誘因之一,但肥胖誘發T2DM的具體分子機制尚不完全清楚。在過去的幾十年里,越來越多的證據表明,脂肪組織已不再是單純的能量儲備器,還是機體最大的內分泌器官[21],可以分泌多種脂肪因子從而參與調節機體生長發育、代謝、免疫應答等生物過程[22]。肥胖狀態下,脂肪因子表達譜的改變,對了解肥胖與糖脂代謝紊亂的關系具有一定意義。新式脂肪因子miRNAs自從被發現以來就備受關注,作為一類非編碼小RNA,廣泛參與基因轉錄后的表達調控[23]。近年來,隨著miRNAs在糖脂代謝領域中的作用越來越受到重視,其作為肥胖相關代謝性疾病的診斷標志物和治療靶點的應用前景十分廣闊。
課題組前期研究發現,miR-4431在肥胖個體血清中的含量顯著增加,并與受試個體血糖水平顯著正相關,體外細胞實驗發現miR-4431可通過抑制下游糖代謝相關基因的表達,誘發靶細胞的IR[10]。最近的研究報道,脂肪組織是循環miRNAs的一個重要來源,大量學者的工作證實:肥胖個體血清和組織中miRNAs的含量發生變化[9]。本研究在肥胖受試個體和飲食誘導的肥胖小鼠脂肪組織中,均發現miR-4431表達量增加,同時肥胖小鼠血清中miR-4431含量也出現相應的增加。以上結果初步提示:miR-4431可能來源于脂肪組織,尚需進一步的研究加以證實。肥胖導致miR-4431高表達的分子機制目前尚不清楚。已有文獻報道,miRNAs的生物發生需要Dicer、Drosha等多種剪切酶的參與,Soufi FG等人的研究證明,高血糖患者臍靜脈內皮細胞中參與miRNAs成熟的基因Dicer、Drosha、DGCR8和Ago-2的表達水平顯著增加[24]。
此外,肥胖后體內游離脂肪酸過度蓄積(以棕櫚酸PA的含量最為豐富),可誘發脂肪細胞內質網應激[13-17],而內質網應激反應也可導致miRNAs含量的增加,如Xu等[25]的研究表明內質網應激能夠特異性誘導肝細胞中miR-26a的上調,Hiramatsu N的研究同樣證實了內質網應激廣泛重塑miRNAs轉錄組,誘導miR-215、miR-483和miR-616的表達[26]。本研究結果表明:肥胖受試個體的脂肪組織中內質網應激標志物顯著高表達,相關性分析結果顯示:脂肪組織miR-4431表達水平與內質網應激標志物表達水平顯著正相關。此外,本研究體外實驗結果表明:高濃度PA可上調脂肪細胞內質網應激標志物的表達,同時促進了miR-4431的表達和釋放。進一步運用TransmiR數據庫預測后發現:在miR-4431的上游轉錄因子中有18個與內質網應激相關。以上結果提示:肥胖后棕櫚酸PA含量的增加可能通過誘發脂肪細胞內質網應激,促進miR-4431的表達和釋放,而內質網應激后具體以何種途徑發揮上述作用,值得今后更深入的研究加以探索和證實。
綜上所述,本研究通過體內外實驗發現,肥胖后脂肪組織miR-4431表達及釋放水平顯著增加,且miR-4431的高表達可能受內質網應激的調控。在本研究基礎上,深入開展內質網應激影響miR-4431表達及釋放的分子機制研究,將為肥胖及相關代謝性疾病的臨床診斷和治療提供新的理論基礎和作用靶點。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
現代臨床醫學(2021年1期)2021-01-26 00:56:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
人大建設(2019年12期)2019-05-21 02:55:32
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
