冬凌草甲素抑制脂多糖誘導(dǎo)小鼠肺泡巨噬細(xì)胞炎癥反應(yīng)
2023-06-12 09:23:32郭民張瑞虎景志杰龐文彪陳朝陽
郭民,張瑞虎,景志杰,龐文彪,陳朝陽
(山西醫(yī)科大學(xué)實(shí)驗(yàn)動物中心,山西 太原 030001)
革蘭陰性菌細(xì)胞壁的主要成分脂多糖(lipopolysaccharide,LPS)是誘導(dǎo)炎癥發(fā)生的重要物質(zhì)[1]。炎癥反應(yīng)可以促進(jìn)機(jī)體克服不利刺激、修復(fù)受損組織并恢復(fù)到穩(wěn)態(tài)[2]。然而,它也可能產(chǎn)生不良后果,包括免疫失衡、進(jìn)一步的組織損傷、敗血癥、器官衰竭等[3]。因此,尋求抗炎制劑來克服炎癥的不良反應(yīng)尤為重要。冬凌草甲素是從冬凌草中提取的一種具有多種生物活性的有效成分,具有抗炎、抗腫瘤等活性[4-5]。文獻(xiàn)報(bào)道冬凌草甲素可以通過核因子κB(NF-κB)、核因子E2相關(guān)因子2(nuclear factor E2-related factor 2,Nrf2)等信號通路抑制炎癥反應(yīng)[6-7],然而,其在不同的炎癥誘導(dǎo)劑、不同的靶細(xì)胞發(fā)揮抗炎作用的機(jī)制不盡相同,對其抗炎機(jī)制仍有必要進(jìn)行深入研究。
郎旭宇等[8]研究發(fā)現(xiàn),冬凌草甲素通過NF-κB通路抑制脂多糖誘導(dǎo)的小鼠肺泡上皮細(xì)胞腫瘤壞死因子(TNF)-α、白細(xì)胞介素(IL)-6和IL-1β表達(dá)而發(fā)揮抗炎作用;米偉等[9]研究發(fā)現(xiàn)冬凌草甲素通過下調(diào)TLR4、MyD88和p-P65表達(dá)進(jìn)而抑制TNF-α、IL-6、IL-1β含量發(fā)揮抵抗高糖誘導(dǎo)的人腎小球系膜細(xì)胞炎癥反應(yīng)。然而冬凌草甲素對其他炎癥介質(zhì)如NO、COX-2和PGE2的抑制作用及可能機(jī)制研究較少。為進(jìn)一步探究冬凌草甲素抗炎作用,本研究選擇脂多糖(LPS)作為炎癥誘導(dǎo)劑,以小鼠肺泡巨噬細(xì)胞(MH-S)為靶細(xì)胞,一氧化氮(NO)、環(huán)氧合酶-2 (COX-2)和前列腺素2(PGE2)為炎癥介質(zhì),在基因和蛋白水平對冬凌草甲素的抗炎作用進(jìn)行了研究,并進(jìn)一步檢測了絲裂原活化蛋白激酶的活力變化,以進(jìn)一步揭示冬凌草甲素的抗炎作用及其作用機(jī)制。
1 材料與方法
1.1 試驗(yàn)藥物
冬凌草甲素(美國Merck公司,批號:3239848,純度≥94.2%),冬凌草甲素溶解于二甲基亞砜(DMSO)中,濃度為20 mg·mL-1,4 ℃避光保存,給藥前用生理鹽水稀釋。脂多糖(LPS,美國Sigma-Aldrich公司,批號:L2880,純度≥97%)。LPS用無菌生理鹽水配置成1 mg·mL-1濃度,分裝后-20 ℃保存,用時室溫解凍。
1.2 細(xì)胞株
小鼠肺泡巨噬細(xì)胞株MH-S(山西醫(yī)科大學(xué)實(shí)驗(yàn)動物中心保存,批號:QCE711)。
1.3 試劑
Trizol(批號:abs9331),反轉(zhuǎn)錄試劑盒(批號:abs601510),2×Real time PCR預(yù)混液(批號:abs601513),上海Absin公司;預(yù)染蛋白marker(批號:RTD6151),超敏ECL化學(xué)發(fā)光液(批號:EC2520),北京中科瑞泰生物科技有限公司;鼠抗ERK1/2單克隆抗體(批號:4696)、兔抗p-ERK1/2(Thr202/Tyr204)單克隆抗體(批號:4370)、兔抗JNK單克隆抗體(批號:9252)、鼠抗p-JNK(Thr183/Tyr185) 單克隆抗體(批號:9255)、兔抗 p38 單克隆抗體(批號:8690)、兔抗p-p38 (Thr180/Tyr182) 單克隆抗體(批號:4511)和鼠抗β-actin單克隆抗體(批號:3700),美國CST公司;HRP-羊抗兔IgG+HRP-羊抗小鼠IgG(武漢博士德有限公司,批號:BA1056);胎牛血清(武漢Procell有限公司,批號: 164210),RPMI-1640培養(yǎng)基(武漢博士德有限公司,批號:PYG0066);NO測定試劑盒(美國Thermo Fisher公司,批號:EMSNO),PGE2酶聯(lián)免疫吸附試劑盒(上海通蔚科研生物科技有限公司,批號:TW8620),BCA蛋白定量試劑盒(上海碧云天生物技術(shù)公司,批號:P0012S)。
1.4 儀器
熒光定量PCR儀(美國ABI公司,型號:7500),超微量核酸檢測儀(上海科器儀器設(shè)備有限公司,型號:Tnano-700),倒置顯微鏡(日本Olympus公司,型號:DSZ2000X),酶標(biāo)儀(山東博科生物產(chǎn)業(yè)有限公司,型號:BK-EL10B),凝膠圖像分析系統(tǒng)(北京天能有限公司,型號:Tanon 3500)。
1.5 方法
1.5.1 細(xì)胞培養(yǎng)
MH-S細(xì)胞培養(yǎng)于含10%胎牛血清的1640 培養(yǎng)基中,置于37℃、含5% CO2恒溫?zé)o菌培養(yǎng)箱內(nèi)培養(yǎng),細(xì)胞融合度大于80% 時,用胰蛋白酶消化傳代。
1.5.2 冬凌草甲素對細(xì)胞活性影響的臺盼藍(lán)實(shí)驗(yàn)
將細(xì)胞以每孔1×105個細(xì)胞的密度接種在24 孔板中,依文獻(xiàn)[10]加入不同濃度冬凌草甲素(0.25、0.5、1.0、2.0 μg·mL-1),同時設(shè)置不加冬凌草甲素組(加入等體積生理鹽水)作為對照組,分別作用24、48、72、96 h。室溫1 500×g離心5 min,收集細(xì)胞后加入0.4% 臺盼藍(lán)對細(xì)胞進(jìn)行染色。將染色后細(xì)胞加在血細(xì)胞計(jì)數(shù)板,光學(xué)顯微鏡下隨機(jī)選取5個視野,計(jì)數(shù)100個細(xì)胞,計(jì)數(shù)活細(xì)胞(未染色)數(shù)量即為活細(xì)胞百分比,以評估冬凌草甲素對 MH-S 細(xì)胞的細(xì)胞毒性。
1.5.3 冬凌草甲素對脂多糖誘導(dǎo)的細(xì)胞炎癥反應(yīng)的影響
當(dāng)MH-S 細(xì)胞生長至融合度約60%時,加入濃度分別為0.5、1.0、2.0 μg·mL-1的冬凌草甲素孵育2 h,同時設(shè)置不加冬凌草甲素組作為對照組,隨后加入0.1 μg·mL-1脂多糖繼續(xù)培養(yǎng)24 h[11],1 500×g離心5 min,分別收集細(xì)胞或細(xì)胞培養(yǎng)物上清液用于如下分析。
1.5.4 NO濃度的Griess測定
取1.2.3節(jié)中細(xì)胞培養(yǎng)上清液50 μL,置于96 孔板中,同時配置50 μL濃度分別為0、31.25、62.5、125、250、500、1 000 pg·mL-1的標(biāo)準(zhǔn)品。先后加入50 μL Griess A液 和G riess B 液,水平搖床輕搖混勻后室溫孵育15 min,550 nm 處測定標(biāo)準(zhǔn)品和樣品吸光度值,繪制標(biāo)準(zhǔn)曲線得出計(jì)算公式并計(jì)算樣品中NO 濃度。
1.5.5 PGE2含量的ELISA法測定
取1.2.3節(jié)中細(xì)胞培養(yǎng)上清液100 μL,加入酶標(biāo)板中,每個樣品設(shè)2 個復(fù)孔,同時加入濃度分別為0、31.25、62.5、125、250、500、1 000、2 000 pg·mL-1的標(biāo)準(zhǔn)品。37 ℃孵育1 h,棄掉上清,洗滌3次,每次3 min;加入100 μL 生物素化抗體工作液,37 ℃ 孵育1 h,棄掉上清,洗滌3次,每次3 min;加入100 μL酶結(jié)合物工作液,37 ℃ 孵育30 min,棄掉上清,洗滌3次,每次3 min;加入90 μL 底物溶液,37 ℃ 避光孵育 15 min;加入50 μL 終止液,在450 nm 波長處檢測吸光度值,繪制標(biāo)準(zhǔn)曲線得出計(jì)算公式并計(jì)算樣品中PGE2含量。
1.5.6 iNOS和COX-2 mRNA表達(dá)量的qRT-PCR法檢測
常規(guī)Trizol法提取1.2.3節(jié)收集的細(xì)胞總RNA,并用微量核酸檢測儀檢測RNA濃度及純度(OD260/OD280值在2.0左右)。取1 μg總RNA作為模板進(jìn)行反轉(zhuǎn)錄,依次加入2×反轉(zhuǎn)錄緩沖液(10 μL)、隨機(jī)引物(0.5 μg·μL-1)1 μL、反轉(zhuǎn)錄酶(1 μL),最后加入無RNA酶水定容到20 μL,42℃溫育15 min,85℃加熱5 min以滅活反轉(zhuǎn)錄酶。以反轉(zhuǎn)錄得到的cDNA為模板構(gòu)建PCR反應(yīng)體系進(jìn)行iNOS和COX-2基因的擴(kuò)增,反應(yīng)體系為2×qPCR 預(yù)混液(10 μL)、cDNA(1 μL)、上下游引物(10 μmol·L-1)分別0.5 μL,最后加水定容到20 μL,每個樣品做3個復(fù)孔。同時擴(kuò)增管家基因β-actin,記錄每個基因擴(kuò)增的Ct 值。
PCR反應(yīng)條件為94 ℃變性30 s;94 ℃變性5 s,60 ℃退火延伸30 s,40個循環(huán)。
采用2-ΔΔCt法用來計(jì)算iNOS和COX-2 mRNA的相對表達(dá)水平,ΔΔCt=(實(shí)驗(yàn)組目的基因Ct值-實(shí)驗(yàn)組管家基因Ct值)-(對照組目的基因Ct值-對照組管家基因Ct值)。所有實(shí)驗(yàn)重復(fù)3次。引物序列見表1。
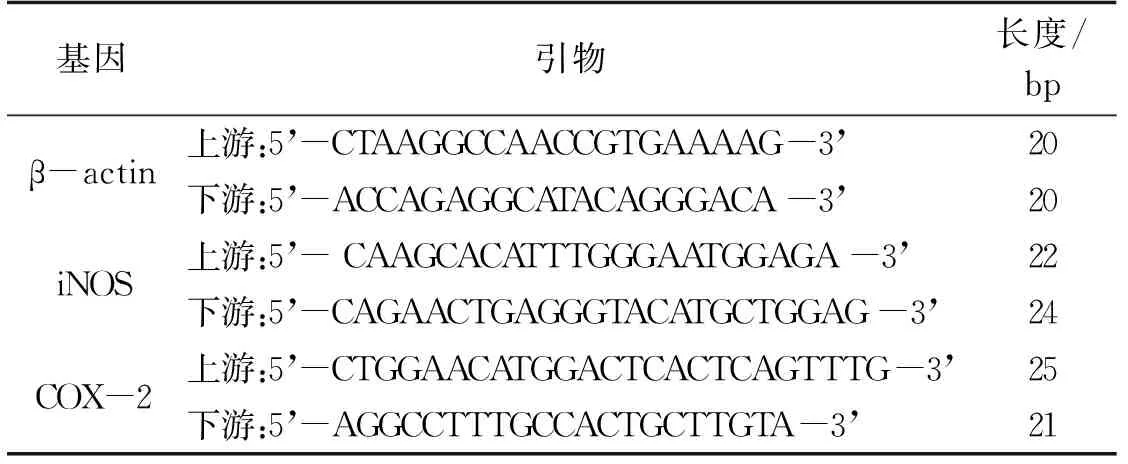
表1 β-actin、iNOS、COX-2的qRT-PCR引物序列
1.5.7 細(xì)胞ERK、JNK和p38及其磷酸化蛋白表達(dá)的Western blot法檢測
在1.2.3節(jié)收集的細(xì)胞中加入100 μl SDS裂解緩沖液(50 mmol·L-1Tris 緩沖溶液(pH8.1),1% SDS),劇烈振蕩后95℃煮沸10 min,室溫13 000×g離心10 min,取上清進(jìn)行BCA蛋白濃度定量,每樣品取25 μg總蛋白進(jìn)行SDS-PAGE,電泳結(jié)束后依次進(jìn)行轉(zhuǎn)模、5%脫脂奶粉封閉1 h、一抗4℃孵育過夜、辣根酶標(biāo)記二抗室溫孵育1 h和ECL顯影,使用Tanon 3500凝膠圖像分析系統(tǒng)分析蛋白條帶灰度。β-actin為內(nèi)參,目的蛋白條帶灰度值與內(nèi)參蛋白條帶灰度值的比值表示目的蛋白表達(dá)水平;磷酸化蛋白條帶灰度值與其對應(yīng)的總蛋白條帶灰度值的比值表示磷酸化蛋白表達(dá)水平。
1.5.8 統(tǒng)計(jì)分析
2 結(jié)果
2.1 冬凌草甲素對MH-2細(xì)胞活力的影響
臺盼藍(lán)染色法結(jié)果顯示不同濃度的冬凌草甲素(0.25、0.5、1.0、2.0 μg·mL-1)處理組細(xì)胞活力與對照組差異無統(tǒng)計(jì)意義(P>0.05),見表2,表明在該濃度范圍內(nèi)冬凌草甲素對MH-S細(xì)胞沒有明顯細(xì)胞毒性。

表2 不同濃度冬凌草甲素處理對MH-S細(xì)胞活力的影響
2.2 冬凌草甲素抑制LPS誘導(dǎo)的細(xì)胞iNOS及NO的生成
Griess 法結(jié)果顯示,對照組MH-S細(xì)胞中NO含量為(0.903±0.16)pg·mL-1,脂多糖誘導(dǎo)組NO含量為(7.10±0.96)pg·mL-1,與對照組相比,脂多糖組NO含量顯著升高,兩組比較具有統(tǒng)計(jì)學(xué)意義(P<0.05),見表3。0.5、1.0、2.0 μg·mL-1冬凌草甲素組NO含量分別為(5.19±0.94)、(3.58±0.47)和(2.79±0.35)pg·mL-1,與脂多糖誘導(dǎo)組比較均降低(P<0.05),見表3,提示冬凌草甲素劑量依賴性地抑制LPS誘導(dǎo)的NO合成。熒光定量PCR結(jié)果顯示與對照組iNOS mRNA(2-△△Ct為1.00±0.13)比較,脂多糖誘導(dǎo)組iNOS mRNA (2-△△Ct為14.34±0.09)表達(dá)顯著升高(P<0.05),見表3;0.5、1.0和2.0 μg·mL-1冬凌草甲素組iNOS mRNA(2-△△Ct分別為10.56±0.48、6.49±0.22和3.41±0.53),與脂多糖誘導(dǎo)組比較,冬凌草甲素組iNOS mRNA 表達(dá)均降低(P<0.05),見表3,提示冬凌草甲素以濃度依賴性方式抑制iNOS mRNA 的表達(dá)。
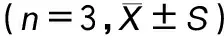
表3 不同濃度冬凌草甲素對LPS誘導(dǎo)MH-S細(xì)胞中NO的含量和iNOS mRNA表達(dá)水平影響
2.3 冬凌草甲素抑制LPS誘導(dǎo)的細(xì)胞COX2及PGE2的生成
RT-qPCR結(jié)果顯示,與對照組COX-2 mRNA(2-△△Ct為1.00±0.13)比較,脂多糖誘導(dǎo)組COX-2 mRNA(2-△△Ct為25.34±2.09)表達(dá)顯著升高(P<0.05)(表4);0.5、1.0和2.0 μg·mL-1冬凌草甲素組COX-2 mRNA(2-△△Ct分別為18.56±2.48、16.49±2.22和7.41±1.03),與脂多糖誘導(dǎo)組比較,冬凌草甲素組COX-2 mRNA 表達(dá)均降低(P<0.05),見表4。提示冬凌草甲素以濃度依賴性方式抑制COX-2 mRNA 的表達(dá)。ELISA法結(jié)果顯示,對照組MH-S細(xì)胞中PGE2含量為(2.03±0.26)ng·mL-1,脂多糖誘導(dǎo)組PGE2含量為(7.58±0.41)ng·mL-1,與對照組相比,脂多糖組PGE2含量顯著升高,兩組比較具有統(tǒng)計(jì)學(xué)意義(P<0.05),見表4。0.5、1.0、2.0 μg·mL-1冬凌草甲素組PGE2含量分別為(6.37±0.34)、(5.58±0.61)和(3.84±0.75)ng·mL-1,與脂多糖誘導(dǎo)組比較均降低(P<0.05),見表4,提示冬凌草甲素劑量依賴性地抑制LPS誘導(dǎo)的PGE2合成。
表4 不同濃度冬凌草甲素對LPS誘導(dǎo)MH-S細(xì)胞中COX-2 mRNA和PGE2的表達(dá)水平影響
2.4 冬凌草甲素抑制LPS誘導(dǎo)的MAPK蛋白的磷酸化
Western blot結(jié)果如圖1和表5 所示,與對照組比較,脂多糖誘導(dǎo)組p38、ERK 和JNK 的磷酸化水平均明顯升高(P<0.05);與脂多糖誘導(dǎo)組比較,冬凌草甲素組p38、ERK 和JNK 的磷酸化水平均均明顯降低(P<0.05),而各組p38、ERK 和JNK 的總含量沒有改變,提示冬凌草甲素以濃度依賴性方式降低其磷酸化。盡管冬凌草甲素對p-p38和p-ERK抑制效果優(yōu)于對p-JNK的效果,但這些結(jié)果表明冬凌草甲素通過抑制MH-S 細(xì)胞中MAPK 活化來發(fā)揮抗炎作用。

1:對照組,2:LPS誘導(dǎo)組,3:0.5 μg·mL-1冬凌草甲素干預(yù)組,4:1.0 μg·mL-1冬凌草甲素干預(yù)組,5:2.0 μg·mL-1冬凌草甲素干預(yù)組。圖1 冬凌草甲素抑制脂多糖誘導(dǎo)的p38、ERK和JNK磷酸化的Western blot檢測
表5 冬凌草甲素對LPS誘導(dǎo)MH-S細(xì)胞p38、ERK 和JNK蛋白表達(dá)的影響
3 討論
近年來,越來越多的中藥提取物被報(bào)道具有體外和體內(nèi)抗炎作用[12]。冬凌草甲素是1種從冬凌草中分離出來的天然物質(zhì),具有抗氧化、抗炎和抗腫瘤活性[4-5],然而其抗炎機(jī)制尚未完全闡明。
誘導(dǎo)劑、傳感器、炎癥介質(zhì)和靶組織構(gòu)成炎癥通路。脂多糖作為革蘭陰性菌外膜的基本成分,已被廣泛用作炎癥誘導(dǎo)劑[13]。肺泡巨噬細(xì)胞表達(dá)Toll樣受體,后者在炎癥發(fā)生中發(fā)揮傳感器作用[14]。肺泡巨噬細(xì)胞作為靶組織細(xì)胞,在炎癥發(fā)生中可以吞噬入侵的病原體并分泌炎癥介質(zhì)(如NO、COX-2和PEG2)。脂多糖是公認(rèn)的巨噬細(xì)胞iNOS轉(zhuǎn)錄合成的最佳誘導(dǎo)劑,L-精氨酸在iNOS作用下生成NO,而NO可以進(jìn)一步促進(jìn)炎癥因子如白細(xì)胞介素-1、腫瘤壞死因子和干擾素-γ的表達(dá),加速炎癥反應(yīng)[15]。COX-2是1種在炎癥因子刺激下生成的誘導(dǎo)性酶,其可以催化花生四烯酸生成前列腺素產(chǎn)物(如PGE2),而前列腺素產(chǎn)物為炎性介質(zhì),參與炎癥的發(fā)生發(fā)展[16]。本研究中,觀察到 LPS 處理顯著增加了MH-S 細(xì)胞中 NO、COX-2 和 PGE2 的產(chǎn)生,而冬凌草甲素以濃度依賴性方式抑制了這些促炎介質(zhì),表明冬凌草甲素具有抗炎作用。
MAPK 信號通路包括ERK、p38 MAPK 和JNK,參與LPS、PM2.5等誘導(dǎo)的細(xì)胞炎癥反應(yīng)[17-18]。MAPK信號通路活化的標(biāo)志是ERK、p38 MAPK 和JNK磷酸化水平升高,一旦MAPK 家族被激活,細(xì)胞內(nèi)信號就會激活各種級聯(lián)信號分子,促進(jìn)炎癥介質(zhì)的表達(dá),而抑制任何或所有三個MAPK信號分子就可以阻止炎癥反應(yīng)。本研究中,LPS 誘導(dǎo)的MH-S細(xì)胞中p38、ERK 和JNK 的磷酸化水平升高,而冬凌草甲素以劑量依賴性方式減弱其磷酸化。綜上,本研究發(fā)現(xiàn)冬凌草甲素通過抑制MAPK 信號轉(zhuǎn)導(dǎo)通路進(jìn)而下調(diào)促炎介質(zhì) NO、COX-2 和 PGE2 的表達(dá),發(fā)揮抗炎作用。
冬凌草甲素抗炎作用及機(jī)制在不同的炎癥誘導(dǎo)細(xì)胞和動物模型中已有研究。宋沁嵐等研究提示冬凌草甲素通過抑制核苷酸結(jié)合寡聚化結(jié)構(gòu)域樣受體3(NLRP3)通路減緩LPS誘導(dǎo)的小鼠Raw264.7巨噬細(xì)胞TNF-α、IL-6和IL-1β等炎癥因子[19];鄶一賀等研究發(fā)現(xiàn)冬凌草甲素通過抑制TLR4-NF-κB信號通路下調(diào)Raw264.7細(xì)胞促炎因子TNF-α、IL-1β、IL-6、IL-10進(jìn)而發(fā)揮抗炎作用[20];Huahong Yang等研究顯示冬凌草甲素通過抑制Nrf2和NF-κB信號通路減輕LPS誘導(dǎo)的急性肺損傷小鼠氧化應(yīng)激和炎癥反應(yīng)[21],這些研究結(jié)果均提示冬凌草甲素可以通過多條信號通路降低LPS誘導(dǎo)的炎性細(xì)胞因子。而本研究采用LPS誘導(dǎo)小鼠肺泡巨噬細(xì)胞MH-S制備炎癥模型,結(jié)果發(fā)現(xiàn)冬凌草甲素可以下調(diào)LPS誘導(dǎo)的炎癥因子NO、COX-2 和 PGE2,機(jī)制研究顯示冬凌草甲素可以抑制LPS激活的MAPK信號通路,尤其對p38和ERK通路的下調(diào)作用比較明顯,而p38/ERK信號通路與NF-κB信號通路在炎癥反應(yīng)調(diào)控中常發(fā)生crosstalk[21],這也可以解釋不同團(tuán)隊(duì)在冬凌草甲素抗炎機(jī)制研究得出不同信號通路的結(jié)論,其實(shí)它們之間相互關(guān)聯(lián),而我們的發(fā)現(xiàn)均是對冬凌草甲素抗炎作用及機(jī)制的有益補(bǔ)充。
