刺山柑種子休眠及萌發特性研究
2023-06-19 01:15:02王云興吾買爾夏提塔漢
種子 2023年4期
王云興, 吾買爾夏提·塔漢
(新疆農業大學生命科學學院, 烏魯木齊 830000)
刺山柑(CapparisspinosaL.)為山柑科山柑屬多年生藤本蔓生落葉小半灌木,又稱野西瓜、抗旱草等[1]。廣泛分布于西班牙、意大利、土耳其和伊朗等國,我國新疆、西藏和甘肅也有分布,常見于石質低山、礫石荒漠、鹽堿地和戈壁灘等[1-2]。刺山柑葉、果、根皮均可入藥,有祛風除濕、止痛、消腫等功效,可治療關節炎、風濕病等;其花蕾作為一種調味品深受歐洲人喜愛[3-4]。刺山柑是一種多用途植物,具有耐旱、耐高溫、耐貧瘠和耐鹽堿等多種優良特性,是荒漠生態恢復與植被重建的優選物種[2]。近年來,刺山柑作為固沙植物具有極高的觀賞價值,廣泛栽培于旱生植物園[2,5]。
刺山柑可通過種子繁殖或莖段扦插繁殖,然而,刺山柑在自然條件下存在萌發率低與生根困難等問題,使其在許多地區繁殖栽培困難,限制了該作物的推廣[3,6-15]。過度放牧和過度收割導致刺山柑資源極度匱乏,甚至還產生了極其嚴重的荒漠生態危機[16-17]。刺山柑種子萌發率低,可能存在休眠,但不同的學者持有不同的觀點[18]。研究表明,可能導致刺山柑種子低萌發的休眠類型分別為物理休眠、生理休眠和組合休眠[3,5,11-12,14,16-17,19]。本研究根據Baskin[20]提出的休眠分類系統,在一定的光照和溫度條件下通過低溫層積、GA浸泡、干藏和埋藏等方法,探討刺山柑種子的休眠類型和種子萌發特性。
1 材料與方法
1.1 材 料
供試材料為刺山柑的成熟種子,2021 年 9 月分兩批采集于新疆哈密二堡鎮(93°01′47″E,42°58′19″N)戈壁灘的自然群落,種子在清水中浸泡從果肉中分離出來,室溫陰干后保存于干燥通風處備用。
1.2 方 法
1.2.1種子的形態指標和千粒重測定
在體式顯微鏡下觀測種子的顏色、形狀及大小,重復20次。隨機選取成熟、發育正常的果實,用游標卡尺分別測量20粒種子的長度。采用四分法取一定量種子(大于4 000粒),將種子分為4份,每份隨機數1 000粒稱重,4次結果加權平均,即得種子千粒重。
1.2.2吸水試驗
為了確定刺山柑種子的吸水情況,2022年3月31日,在實驗室將刺山柑種子分別放入5個墊有雙層濕濾紙的培養皿中,每皿50粒。先用Sartorius BS 124 S型(0.000 1 g)電子天平稱量刺山柑種子的原始重量,從未吸水(干種子)起,每隔30 min取出(隨種子的吸水速率而適當調整稱重時間),用濾紙吸干種子表面的水分后,在電子天平上稱重,直到種子質量不再增加為止,繪制吸水曲線,分析種子的吸水動態變化。
吸水率/%=[(濕重-干重)/干重]×100%。
1.2.3萌發特性研究
將種子置于1%的次氯酸鈉溶液中消毒10 min,然后用自來水沖洗3遍,蒸餾水沖洗1遍,每處理均為25粒種子,4次重復,共100粒。種子均勻擺放于墊有2層濾紙,直徑為90 mm的培養皿中,持續培養28 d。實驗的光照條件設置為每天12 h光照[100 μmol/(m2·s),400~700 nm,白熾燈光]和全黑暗,黑暗處理是將培養皿包裹于錫紙中。
胚根突破種皮視為萌發。在光照條件下培養的種子每24 h統計一次萌發數,并將萌發的種子取出,每天加適量蒸餾水以保持濕度一致;全黑暗處理的種子要保持完全避光,萌發率在綠光燈下每7 d檢測一次。實驗結束后未萌發的種子用2,3,5-三苯基氯化四唑(TTC)檢測種子活力。
1) 室內干藏(后熟)
將剛收獲的種子于室溫下干藏,分別檢測貯藏0(剛采收),3,6,9,12個月種子的萌發率。培養皿置于15 ℃/2 ℃、20 ℃/10 ℃、25 ℃/15 ℃及30 ℃/15 ℃等溫變周期(12 h光照/12 h黑暗或全黑暗)條件下培養28 d。不同的溫度反映野外生境中一年四季的溫度變化,即5 ℃/2 ℃(11月),15 ℃/2 ℃(3—4月),20 ℃/10 ℃(5月、9—10月),30 ℃/15 ℃(6—8月);冬季生境中的溫度低于0 ℃,種子在該溫度下不會萌發。
2) 野外埋藏
本研究的目的是確定在野外打破休眠的時間以及埋藏的種子是否經過休眠周期。2021年10月25日,將干藏1個月的種子裝入珍珠紗網袋中,共10袋。將網袋分別埋入盛滿土壤的塑料盆(深為15 cm,直徑為20 cm)的2 cm深處,然后將盆埋在新疆農業大學實驗地中,使種子經歷野外自然溫度和土壤水分條件的變化。
2021年11月25日—2022年6月,每月定期從試驗地取回一個花盆于最適萌發溫度(15 ℃/2 ℃)和光照(12 h光照/12 h黑暗或全黑暗)條件下對種子進行28 d的萌發試驗(2021年12月至翌年2月,積雪覆蓋除外)。
3) 冷層積
將剛收獲的種子置于直徑為120 mm裝有濕河沙的玻璃培養皿中,用錫紙包裹后置于4 ℃恒溫冰箱中層積0,4,6,8周。層積處理后,將種子置于最適萌發溫度(15 ℃/2 ℃)和光照(12 h光照/12 h黑暗或全黑暗)條件下連續培養28 d。
4) 赤霉素對種子萌發的影響
將干藏的種子定時(時間隨干藏處理而定)分別取回,置于不同濃度(0,100,300,500,700 mol/L)的赤霉素溶液中浸泡24 h后,在最適萌發溫度(15 ℃/2 ℃)和光照(12 h光照/12 h黑暗或全黑暗)條件下培養28 d。
1.2.4數據處理
采用SPSS 26軟件對所得數據在95%水平上通過二因素方差分析(Two-way ANOVA)或三因素方差分析(Three-way ANOVA)檢驗差異的顯著性,萌發結果以百分率±標準誤表示,圖表采用GraphDad Prism 9軟件繪制。
2 結果與分析
2.1 種子的形態指標和千粒重測定
刺山柑種子腎形,成熟種子為深棕色或黑褐色,長(2.4±0.2)mm,寬(1.7±0.2)mm,厚(1.4±0.2)mm,千粒重(3.931 6±0.096 2)g(圖1),種子小且種皮厚。

圖1 刺山柑果實(A)、成熟果實(B)、種子(C~D)
2.2 種子的吸水實驗
完整的刺山柑種子吸水速度較慢,在水中浸泡12 h其重量僅增加(24.48±1.44)%,種子的吸水于第4天才開始穩定,增重(37.5±1.77)%,達到飽和狀態(圖2)。表明刺山柑種子吸水較慢且吸水較少。
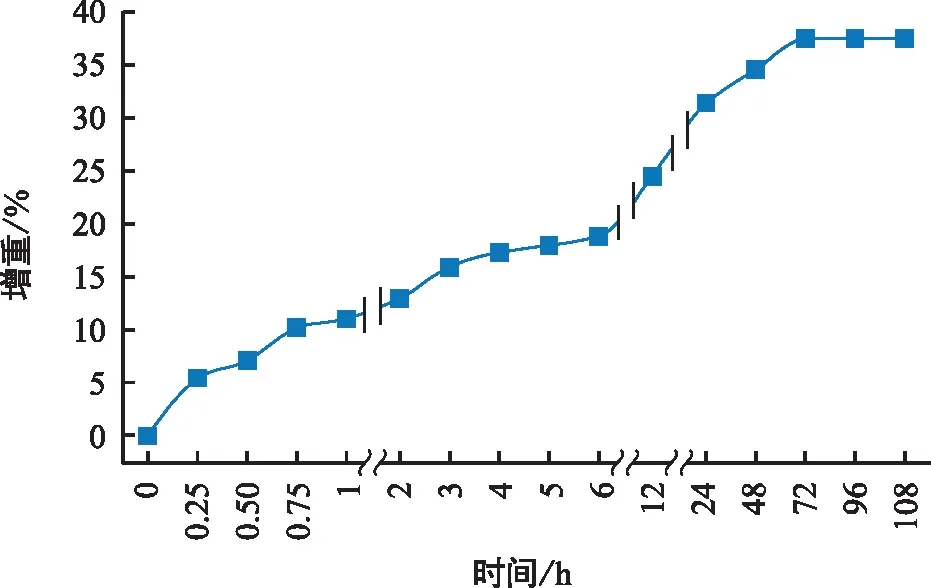
圖2 刺山柑種子的吸水曲線
2.3 干藏對刺山柑種子萌發的影響
三因素方差分析結果(表1)表明,光照、干藏時間、光照與溫度的交互作用及其光照、溫度與干藏時間的交互作用對種子萌發無顯著性影響,其他因素對種子萌發均有顯著性影響。活力檢測后發現,98%未萌發種子是有活力的。

表1 室內干藏條件下,光照、溫度、干藏時間及其交互作用對刺山柑種子萌發影響的三因素方差分析
預實驗結果表明,刺山柑種子在5 ℃/2 ℃條件下不萌發,故該溫度在干藏實驗中去除。15 ℃/2 ℃為種子的最佳萌發溫度,除干藏試驗外,其他萌發試驗均于該溫度下進行。
新收獲的種子在各溫度和光照條件下不萌發或有較低的萌發率,剛收獲的成熟種子在15 ℃/2 ℃溫度下具有較高且穩定的萌發率。種子的萌發率并沒有隨著室內干藏時間的延長而發生顯著性變化(圖3),最大萌發率出現在干藏的第3個月,在15/2 ℃的光照和全黑暗條件下分別達(52.00±11.31)%和(55.00±18.29)%。說明干藏處理不能解除刺山柑種子的休眠。
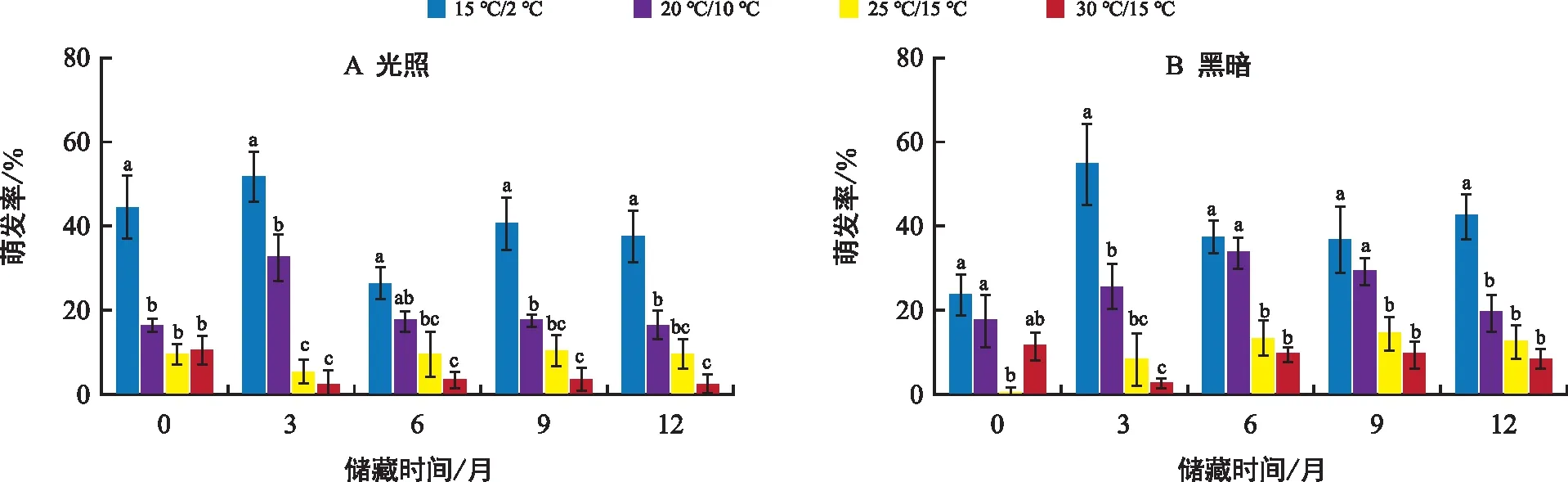
注:不同小寫字母表示相同儲藏時間在不同溫變梯度下萌發率存在差異顯著(p<0.05)。
2.4 野外埋藏對刺山柑種子萌發的影響
二因素方差分析結果(表2)表明,光照、光照與埋藏時間的交互作用對種子的萌發率沒有顯著影響,埋藏時間對種子的萌發率有顯著影響。
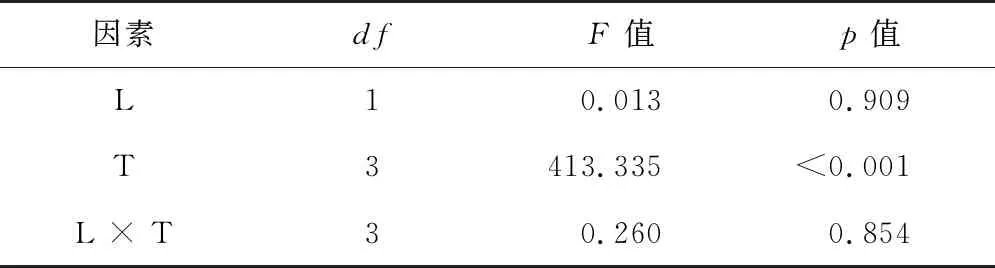
表2 光照和埋藏時間及其交互作用對刺山柑種子萌發影響的二因素方差分析
野外自然溫度下,隨著埋藏時間的延長,種子的萌發率逐漸提高(圖4)。埋藏的第6個月,種子的萌發率在各光照條件下分別達到(90.00±1.15)%和(93.00±6.83)%,說明埋藏能解除種子休眠。
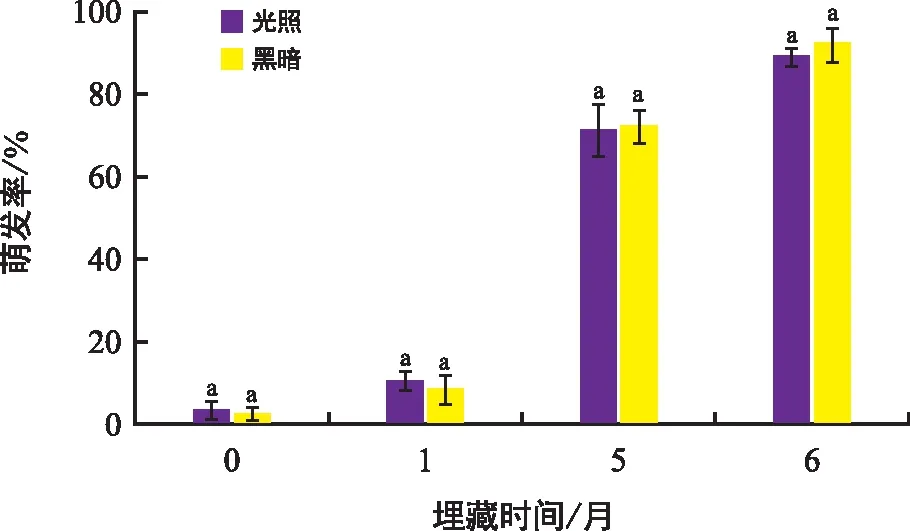
圖4 刺山柑種子野外埋藏0,1,5,6個月在光照和黑暗及15 ℃/2 ℃溫變梯度下的最終萌發率
2.5 冷層積對刺山柑種子萌發的影響
二因素方差分析結果(表3)表明,光照、冷層積時間、光照與冷層積時間的交互作用對種子萌發率均有顯著影響。

表3 光照和冷層積時間及其交互作用對刺山柑種子萌發影響的二因素方差分析
在4 ℃恒溫冰箱進行冷層積處理后,種子萌發率隨層積時間的延長而顯著提高(圖5),在黑暗條件下的萌發率比光照條件下的稍高,說明低溫層積能快速解除種子休眠。
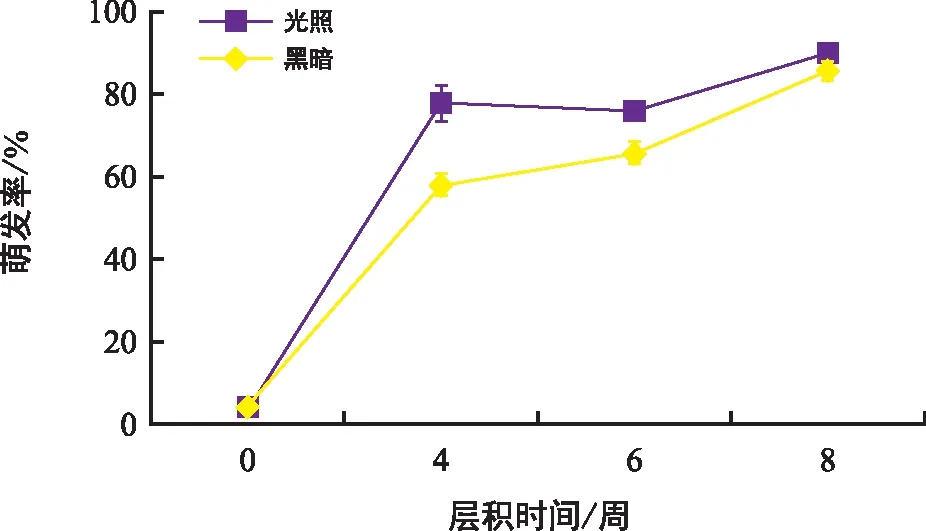
圖5 刺山柑種子冷層積0,4,6,8周在光照和黑暗及15 ℃/2 ℃溫變梯度下的最終萌發率
2.6 赤霉素對刺山柑種子萌發的影響
三因素方差分析結果(表4)表明,光照和干藏時間的交互作用、光照與干藏時間和赤霉素濃度的交互作用對種子萌發率無顯著影響。光照、干藏時間、赤霉素濃度、光照與干藏時間的交互作用、干藏時間與赤霉素濃度的交互作用對種子萌發率有顯著影響。
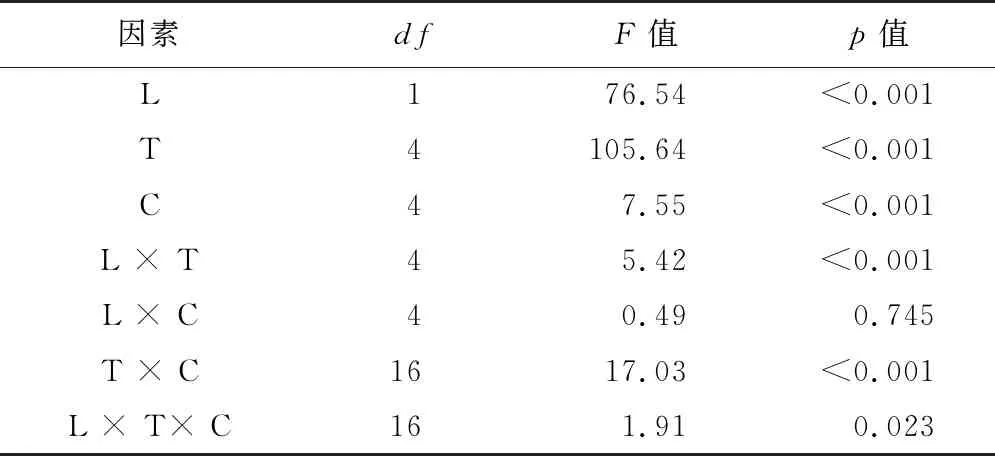
表4 室內干藏條件下,光照、干藏時間、赤霉素及其交互作用對刺山柑種子萌發影響的三因素方差分析
室內干藏的種子經不同濃度赤霉素浸泡24 h后,在黑暗環境中有較高的萌發率,萌發率隨干藏時間的增加呈先增加后減少的趨勢(圖6)。除第6個月外,其余處理的萌發率在赤霉素濃度為300 mg/L時相對較高且穩定,第9個月萌發率在光照和黑暗條件下均最大,分別為(55.00±3.416)%和(65.00±3.41)%。結果表明,赤霉素處理能明顯提高種子萌發率,300 mg/L 赤霉素+黑暗條件為該處理下的最佳萌發方案。

注:圖中不同小寫字母表示相同儲藏時間在不同赤霉素濃度梯度下萌發率存在顯著差異(p<0.05)。
3 討 論
休眠是指整個植物或植物器官以生長和發育的暫時停止為特征的一種狀態[21]。目前國際上認可的種子休眠類型有五類,分別是物理休眠(PY)、形態休眠(MD)、生理休眠(PD)、生理形態休眠(MPD)和組合休眠(PY+PD)[20]。刺山柑種子因種皮特性引起休眠的報道較多[3,5,10,12,16,22],但本研究結果表明,刺山柑的種子具有一定的吸水能力,這與Foschi等[11]的研究結果一致,表明水分不是限制刺山柑萌發的主要因素,即刺山柑種子不存在物理休眠。刺山柑種子不易吸水有兩種原因:一是刺山柑種子的種皮木質化,只能從種臍吸水;二是種子內脂肪含量高達30%[19,23]。另外,在一年的干藏實驗中,刺山柑種子的萌發率在各溫變周期均無顯著提高,說明種子不存在后熟現象,即刺山柑種子不存在形態休眠。綜上所述,刺山柑種子存在生理休眠。生理休眠又分為深度生理、中度生理和非深度生理休眠[24],本研究中,通過赤霉素處理和低溫層積8周均可顯著提高刺山柑種子萌發率,說明刺山柑屬于非深度生理休眠。
溫度對種子有重要的生理作用。溫度不僅可以改變種子的休眠狀態,即低溫或高溫層積可解除種子的休眠狀態。輪葉碎米薺(Cardamineconcatenata)種子在5 ℃冷層積28周后萌發率達到98%;在30 ℃/15 ℃溫度條件下進行4周暖層積預處理后于5 ℃下冷層積(持續50周),萌發率可達100%[25]。本研究結果表明,低溫層積可顯著提高刺山柑種子的萌發率,僅層積8周便可解除休眠,萌發率達90%。此外,溫度還會影響無休眠種子的萌發速率,種子在萌發過程中代謝反應活躍,在一定的溫度范圍內,種子的萌發進程隨溫度的升高而加快。大多數種子在變溫條件下的萌發率較恒溫條件下高。變溫可加快酶的催化作用,促進貯藏物質的轉化,其次變溫利于種殼軟化,利于種胚突破種皮,促進萌發。Hypericumericoides在12 ℃/20 ℃變溫條件下萌發率幾乎達100%。變溫還有助于幼苗建成,如沙芥和斧翅沙芥種子在黑暗和20 ℃/30 ℃變溫條件下有利于種子萌發和幼苗建成[26]。李欣勇等[27]對風車草種子的萌發特性研究發現,恒溫和變溫條件下的萌發率有顯著差異,恒溫下的萌發率顯著低于變溫。不同物種的種子最適萌發溫度有差異,種子萌發的最適溫度與植物各自的生境有密切關系。本研究結果表明,刺山柑種子經4 ℃低溫冷層積處理后,萌發率迅速提高,且15 ℃/2 ℃的溫變周期為刺山柑種子最適萌發溫度。
在荒漠或半荒漠地區,不同植物種子的萌發對光照有不同的要求。雀麥屬(Bromus)植物種子在萌發時并不需要光照[28],而油蒿的瘦果在光照條件下萌發,在暗條件下萌發受到抑制[29]。許多荒漠植物的種子在光照和黑暗條件下萌發都很好,部分在暗中的萌發較在光下好,如梭梭種子[30]。本研究結果表明,干藏、埋藏處理的種子萌發率在光照和黑暗條件下均無顯著性差異,層積和赤霉素處理的種子在黑暗條件下萌發較光照下好,說明刺山柑種子的萌發對光照沒有嚴格要求,在黑暗條件下較光下稍好。這與馬生軍等[31]對刺山柑發芽條件研究的結果一致。
刺山柑作為一種落葉小灌木,在自然生境中,5月始花,6—9月生產果實,果熟期為1個月,10月植株的地上部分死亡。因此,在2021年9月采集當年的種子進行實驗,采集過程中并未發現掉落的種子有出苗現象,于當年10月將刺山柑種子埋藏于試驗地中,經6個月野外埋藏的刺山柑種子全在網袋中萌發。埋藏通過調節土壤濕度和溫度調節種子萌發,冬季野外生境溫度低于0 ℃,大雪消融后取回埋藏的刺山柑種子進行萌發實驗,發現其萌發率得到顯著提高,并在4月達到90%以上。綜上,該處理等同于低溫層積處理打破種子休眠,這與Gulizebo等[8]的研究結果一致。
赤霉素(GA)是一種調控植物許多關鍵生理過程(如種子萌發、根和莖的伸長、開花、坐果和種子發育)的重要激素。GA能取代一些種子對低溫后熟的需求,通常可解除種子的淺度生理休眠。外源添加赤霉素可直接提高其種子內部赤霉素含量,促進種子萌發。休眠的石生茶藨(Ribessaxatile)種子經低溫層積30 d后使用300 mg/L的GA3溶液處理2 h,萌芽率可高達82.67%[32]。然而,GA對解除生理休眠的作用因種而異。GA處理對美國梧桐(Acerpseudoplantanus)種子的萌發率沒有顯著提高,而細胞分裂素能夠提高45%[33]。本研究結果表明,GA3處理能顯著提高刺山柑種子萌發率,300 mg/L的GA處理下,刺山柑種子萌發率較高且穩定,這與Bahrani等[7]的研究結果一致。
4 結 論
1) 刺山柑種子存在非深度生理休眠;
2) 種皮具透水性;
3) 最適萌發溫度為15 ℃/2 ℃;
4) 光照不是刺山柑種子萌發的必要條件;
5) 可通過低溫層積打破休眠,赤霉素能顯著提高其萌發率。
