云南獨龍雞和紅原雞卵巢組織轉錄組水平的比較分析
2023-06-30 12:00:49丁玥竹任宇扈孟雪燕文荃郭愛偉周杰瓏吳培福陳粉粉劉莉莉
江蘇農業科學 2023年5期
丁玥竹 任宇 扈孟雪 燕文荃 郭愛偉 周杰瓏 吳培福 陳粉粉 劉莉莉
摘要:為明確獨龍雞的基因功能和種質特性,試驗采集3羽健康狀況良好的獨龍雞卵巢組織進行轉錄組測序,并與飼養條件基本相似并且同周齡的紅原雞卵巢組織進行比較。結果表明,與紅原雞相比,獨龍雞中差異表達基因共有7 404個。KEGG和GO富集分析表明,差異表達基因主要富集在GTP酶激活劑活性、膜結構、泛素蛋白連接酶活性和ATP結合等過程,并參與Ras信號通路、硒化合物的代謝通路、SNARE因子在囊泡運輸中的相互作用等通路。對顯著性通路分析發現,獨龍雞在蛋品質和免疫抗性上具有一定優勢,但在產蛋性能上與紅原雞還存在一定差距。說明NCBP1、NCBP2、TXNRD1、PTEN、ENOX1基因主要富集在抗性、產蛋性能和蛋品質相關通路中,可進一步深入研究。
關鍵詞:獨龍雞;紅原雞;卵巢組織;轉錄組測序;差異表達基因
中圖分類號:S831.2文獻標志碼:A
文章編號:1002-1302(2023)05-0038-06
獨龍雞是我國怒江傈僳族自治州貢山縣獨龍族的一種特有的地方家禽,僅在高山河谷地區出現,于2010年入選全國畜禽遺傳資源名錄[1]。獨龍蛋品質好,蛋質鮮美,且有較強的捕食力,會飛翔,抗病力極強,且肉質鮮美,特別適合中高海拔、高溫潮濕等不良環境,是云南地區特有的優良家禽品種,適宜在山地飼養[2]。目前,獨龍雞主要是由山區人民自行繁殖飼養的,由貢山縣畜牧獸醫站承擔疾病防治工作,但是由于地理位置特殊、交通不便、疾病防治設備陳舊、藥品短缺、繁育水平較低、養殖效益較差,導致獨龍雞在種質資源開發和利用等方面發展緩慢[3],因此深入研究獨龍雞的種質資源特點迫在眉睫。
卵巢是雞蛋形成的主要部位之一,是雌性家禽動物調控生殖的主要器官,卵巢所產生的雌二醇、孕酮等類固醇激素對卵泡發育成熟和排卵具有促進作用。目前,轉錄組測序(RNA sequencing,RNA-seq)在雞重要經濟性狀的候選基因鑒定方面已逐步成熟了[4],但是對獨龍雞的生產性狀研究較少。因此,本研究擬通過RNA-seq技術比較獨龍雞和紅原雞卵巢組織的基因表達水平,以分析獨龍雞和紅原雞的種質差異,為以后獨龍雞重要生產性狀候選功能基因篩選、品種改良、種質特性以及基因功能驗證等研究奠定基礎,為保護云南獨龍雞品種和開展分子育種改善獨龍雞雞蛋品質提供理論基礎。
1 材料與方法
1.1 樣本采集
試驗所需的雌性獨龍雞種蛋,由云南省貢山縣神山農產品發展公司生產供應,在西南林業大學基地培育,在與養殖要求相似的環境下,自然采食和飲水。隨機挑選43周齡健康的獨龍雞3羽,解剖收集卵巢組織,編號為LC1、LC2、LC3,并立即超低溫保存備用。
1.2 總RNA提取和RNA質量檢測
應用 TRIzol RNA技術,從卵巢中分離出全部 RNA。利用Nanodrop 2000和Agilent 2100等先進儀器設備,對RNA的含量、純化程度和完整性等進行測定,以確定轉錄組檢測工作是采用了符合要求的樣品完成的。得到的RNA樣品濃度≥500 ng/μL,RIN值≥8且28S ∶18S≥1。
1.3 cDNA文庫的構建及測序
mRNA利用Oligo (dT)的磁性微粒加以富集,而后添加分離緩沖劑(fragmentation buffer),將mRNA隨機中斷。首先用mRNA作為模版,通過六堿基隨機引物(random hexamers)加以制備,之后再添加緩沖液、dNTPs、RNase H、DNA Polymerase Ⅰ,最后再用AMPure XP beads對cDNA加以提純。對純化后的cDNA經過末端修飾,加A尾,并與測序接頭進行相連。最后,再使用AMPure XP beads,對文庫中插入的片段加以篩選,然后用 PCR技術對其進行擴增。利用q-PCR方法對新構建的文庫實現更準確的定量分析,因而保證了在文庫中的有效含量>2 nm。庫檢通過后,可以利用Illumina的高通量測序平臺,完成排序解析。
1.4 測序數據及其質量控制
應用 Illumina HiSeq TM 2000軟件,通過對序列進行高通量測序,以及采用將序列中較低質量的序列切除、去連接等方式,得到了高質量的 Clean reads序列。
1.5 差異表達分析
采用飼養條件基本相似并且周齡較相近的紅原雞卵巢組織測序,結果設為對照組,本研究中的獨龍雞卵巢組織樣本設為試驗組,進行差異表達分析。并采用FPKM(fragments per kilobase of transcript per million fragments mapped)值表示對應Unigene 的表達豐度。篩選標準為差異倍數(fold change,FC)≥2且錯誤發現率(false discover date,FDR)<0.01,并使用DESeq2軟件分析樣品間的差異表達,獲得總差異表達基因。試驗組中差異表達基因的表達高于對照組中差異表達基因的表達,表現為上調基因(up-regulated gene),反之為下調基因(down-regulated gene)。
1.6 差異表達基因富集與互作分析
應用DAVID(http://david.ncifcrf.gov/)網絡軟件,對數據分析得到的差異表達基因采用KEGG(kyoto encyclopedia of genes and genomes)通路分析方法及GO(gene ontology)富集的研究。P<0.05的GO條目和KEGG通路的顯著富集。應用STRING(https://www.string-bd.org/)網絡軟件對差異表達基因開展了互作分析,提取的蛋白質-蛋白質的相互作用對的互作得分>0.9。
2 結果與分析
2.1 測序數據統計
以43周齡獨龍雞、紅原雞為例,對其基因序列進行檢測,經測序獲得39.29 Gb高質量數據,各樣本高質量數據均達到9.48 Gb,每個樣品Q30的堿基百分數都不低于82.06%,測得的數據總量和質量都均滿足后續試驗分析(表1)。
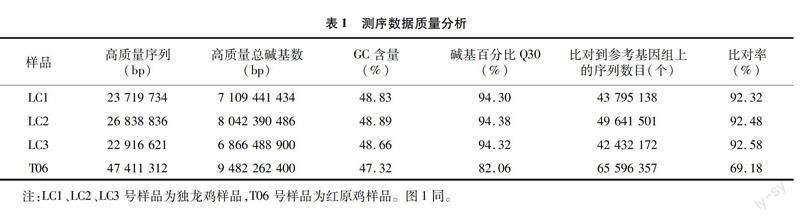
2.2 樣品間的相關性評估
FPKM是一種用于預測轉錄組序列基因表達水平的方法,它是通過比較每個千堿基長度的基因讀出數量。皮爾遜相關因子(r)是衡量生物重復關系的一個重要指標,r2與1越接近,證明2個樣品的關系愈密切,則2個樣本間的關聯度愈高。同一條件下每對生物重復樣本的基因表達相關性分析見圖1,獨龍雞LC1、LC2、LC3號樣品間相關系數大于0.9,說明各樣品間相關性較好,與紅原雞T06號樣品的相關系數均小于0.7,說明與紅原雞樣品差異較大,表明試驗可靠性較高。
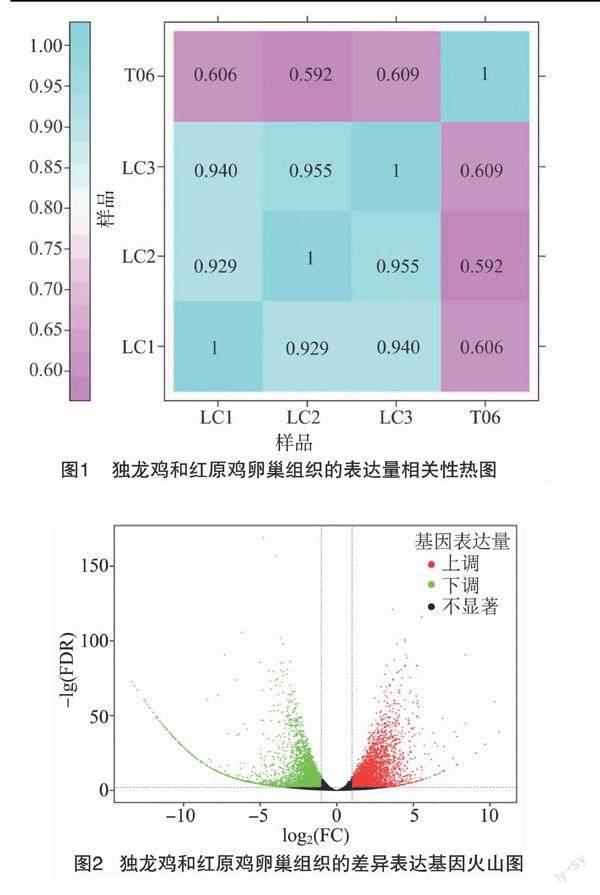
2.3 差異表達基因篩選
以紅原雞為對照組,并以|log2 Fold Change |≥1,FDR值<0.01為條件,一共篩選出差異表達基因7 404個,包括3 853個上調基因,以及3 546個下調基因。這2組樣品基因表達水平的差別,以及不同的統計學顯著性通過火山圖顯示(圖2)。
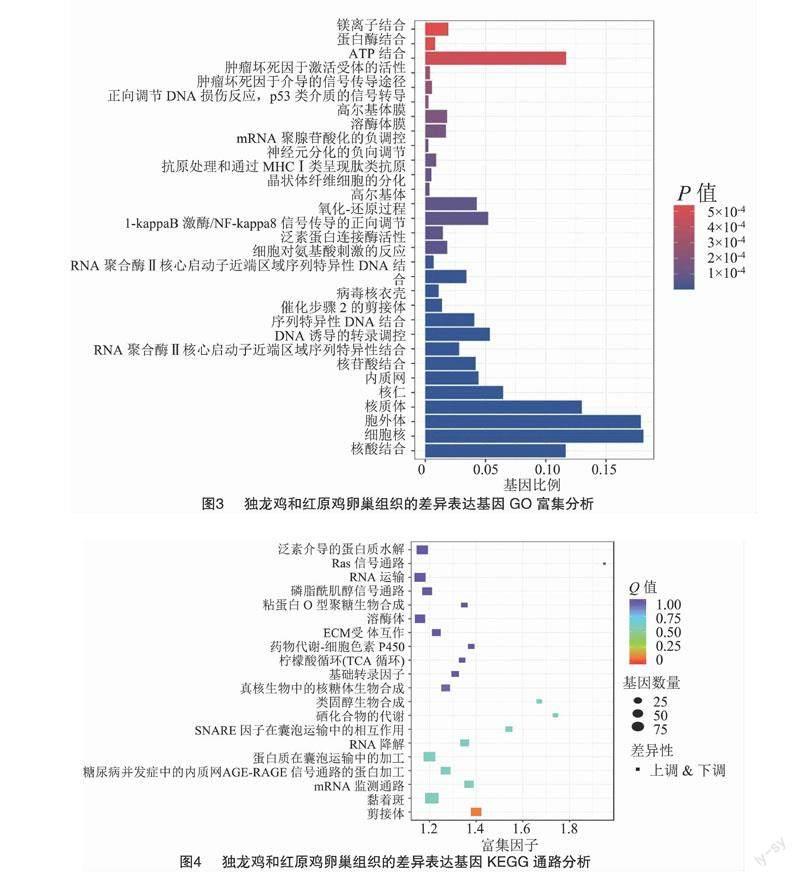
2.4 差異表達基因富集分析
用GO分析法研究紅原雞與獨龍雞的差異表達基因, 研究結果顯示, 在生物學過程(BP)中上調基因多注釋到細胞對氨基酸刺激的反應(GO:0071230)、氧化-還原過程(GO:0055114)、I-kappaB激酶/NF-kappaB信號傳導的正向調節(GO:0043123)、腫瘤壞死因子介導的信號傳導途徑(GO:0033209)、正向調節DNA損傷反應,p53類介質的信號轉導(GO:0043517),而下調基因主要與DNA誘導的轉錄調控(GO:0006355)、晶狀體纖維細胞的分化(GO:0070306)、抗原處理和通過MHC Ⅰ類呈現肽類抗原(GO:0002474)、對神經元分化的負向調節(GO:0045665)、mRNA聚腺苷酸化的負調控(GO:1900364)等有關;在細胞組分(CC)中,上調基因多與胞外區有關,如胞外體(GO:0070062)、內質網(GO:0005783)、高爾基體(GO:0005794),還與細胞膜結構有關,如溶酶體膜(GO:0005765)、高爾基體膜(GO:0000139),而下調基因多與細胞物質相關,如細胞核(GO:0005634)、核質體(GO:0005654)、核仁(GO:0005730)、催化步驟2的剪接體(GO:0071013)、還與病毒核衣殼(GO:0019013);在分子功能(MF)中,上調基因可顯著注釋到泛素蛋白連接酶活性(GO:0061630)、腫瘤壞死因子激活受體的活性(GO:0005031),還注釋到ATP結合(GO:0005524)、蛋白酶結合(GO:0002020)、鎂離子結合(GO:0000287)、核苷酸結合(GO:0000166)、核酸結合(GO:0003676),而下調基因主要與RNA聚合酶Ⅱ核心啟動子近端區域序列特異性DNA結合(GO:0000978)、RNA結合(GO:0044822)、轉錄激活劑活性(GO:0001077)、核苷酸結合(GO:0000166)、序列特異性DNA結合(GO:0043565)、RNA聚合酶II核心啟動子近端區域序列特異性結合(GO:0001077)有關,上調基因和下調基因共同注釋了核苷酸的調節,于此相關的ENOX1基因[5]與蛋品質密切相關。具體結果如圖3所示。GO分析表明,獨龍雞中上調差異基因主要參與了免疫相關的生理生化過程,下調差異基因多與基因表達調控、蛋白質合成有關。
使用KEGG富集分析法可以確定差異表現基因的主要生物化學代謝路線和信息通道。差異表達基因KEGG的通道富集分析結果見圖4,并展示了顯著性Q值最小的前20個通道。其中7條通道最顯著富集(P<0.05),分別是剪接體(ko03040)、Ras信號(Ras蛋白是ras基因表達產物)通路(ko04014)、硒化合物的代謝(ko00450)、SNARE因子在囊泡運輸中的相互作用(ko04130)、類固醇生物合成(ko00100)、RNA降解(ko03018)、mRNA監測通路(ko03015)。
通過對差異基因的富集分析和文獻檢索,發現FoxO3、CTH、TXNRD1、ENOX1涉及4個GO條目和1個KEGG通路,且與蛋白質合成有關,因此這些基因可能與蛋白品質相關;CTH、TXNRD1、TXNRD2、TXNRD3參與硒化合物的代謝信號通路。
2.5 差異蛋白互作網絡分析
差異表達基因的相互關系采用 STRING數據庫分析,發現互作網絡共有1 879個節點和11 035個相互作用(圖5),其中,差異表達基因互作主要集中在NCBP1、NCBP2、PTEN、TXNRD1等組裝調控上。
3 討論與結論
獨龍雞來自怒江傈僳族自治州貢山縣,為當地的特色地方禽類種類,對獨龍雞習性和種質特性的相關研究相對較少。雞蛋是人類消費營養食品中優質動物蛋白源之一,具有很大的經濟價值。在優質蛋雞的選育中,通常把產蛋和蛋質性狀作為兩大指標[6]。因此,本研究采用轉錄組測序技術對紅原雞和獨龍雞卵巢組織中的差異表達基因進行篩選,并應用生物資料庫對不同表達基因參與的生物學過程和信號途徑進行研究,在RNA水平上研究獨龍雞種質特性,為以后獨龍雞生產性狀的研究,包括優良性狀基因的探索以及研究提供了理論依據。
對不同表達基因 GO功能進行分析,結果表明,這些上調基因在細胞膜結構、ATP結合、GTP酶激活以及泛素結合酶等方面具有重要的生物作用。在生物體中,ATP存在于組織細胞內,儲存能量以備機體需要[7],ATP酶能夠將大量代謝需要的物質輸入到細胞中,并將毒素、代謝廢物和其他會妨礙細胞過程的物質輸出;GTP類似于ATP,為生物蛋白的合成直接供能[8];生物膜是一種重要的細胞結構[9],它與能量、信息、物質的運輸[10]有關,尤其是與膜相關的基因ACPP、APOO、ADGRA2等顯著上調有關。基于以上結果,本研究推測相較于紅原雞,獨龍雞有較強的適應能力和抗性,更能夠適應高海拔地區的惡劣條件,這為獨龍雞的抗病育種奠定了理論基礎。
KEGG通路富集分析顯著性最可靠的是Ras信號通路、硒化合物的代謝通路、類固醇生物合成通路、SNARE因子在囊泡運輸中的相互作用和剪接體通路。研究表明Ras信號通路與胰島素和葡萄糖轉運蛋白有關[11],葡萄糖運輸蛋白是控制葡萄糖代謝的重要載體[12]。葡萄糖可以提高代謝,對肝臟有保護功能,對雞群盡快恢復體力有很大幫助。SNARE因子是囊泡表面細胞轉運的介質,是轉運途徑中的核心蛋白[5]。SNARE因子在囊泡和靶向細胞的膜之間形成復合物,導致膜融合并且允許囊泡內容物遞送到靶細胞中。通過這種方式,SNARE因子在生物合成通路中發揮重要作用,并且在內吞循環通路和受調節的膜運輸中同樣起著重要作用,如神經分泌或胰島素的釋放[13]。雖然人們對某些系統中SNARE因子介導的膜運輸研究較少,但近年來對癌細胞背景下膜運輸的研究顯著增加[14]。SNARE 介導的侵襲性蛋白質膜的運輸有助于膜的重塑以及信號傳導成分、黏附受體和 ECM 降解酶的形成[15]。Williams等研究表明,特定的SNARE因子在運輸侵入性腺苷酸相關蛋白以促進惡性癌細胞的細胞入侵和遷移中闡明了重要的作用[16]。研究結果表明,獨龍雞相較于紅原雞,具有良好的抗性和耐高壓環境,與SNARE因子合成通路有密不可分的關系。
硒作為維持家禽生長所必需的微量元素,具有廣泛的作用。雞體內缺少硒會引起多種疾病,甚至造成死亡。其機制表現為:一是人體內抗氧化酶的形成,可保護細胞膜免遭氧化破壞,并維持其通透性;二是與硒蛋白產生螯合作用,降低有毒物質的毒性。硒被生物學家們稱為體內微量元素中的“防癌之王”。硒能提高免疫力,有抗氧化和解毒排毒等作用。硒的最重要的生理功能是構成硒谷胱甘肽過氧化物酶,它將人體細胞代謝產生的無機和有機氧化物轉化為水和羥基化合物[17],有效地清除自由基[5],因此硒在人體內參與了抗氧化作用,對機體的抗氧化起到了重要的作用[17]。并且硒可以分解組織中脂質氧化產生的過氧化物的影響,在保護細胞、細胞器和細胞膜結構的完整性和穩定性方面發揮著重要作用[18]。硒能加強身體的免疫系統,并能預防許多疾病,并且對抗衰老和抗腫瘤有比較重要的作用。最新研究表明,硒還與雞的含硫氨基酸代謝有關,硒與雞的生長、產蛋率和孵化率也都密切相關[18]。本研究發現,差異基因主要富集在硒化合物的代謝通路中,尤其是大部分富集基因顯著上調,如TXNRP1基因顯著上調,表明相較于紅原雞,獨龍雞更能夠適應高海拔和低溫的環境,有較好的免疫能力,并有較強的產蛋能力和較高的蛋品質,且其肉質和蛋中可能富含硒元素,具有較高的肉品質和蛋品質。
獨龍雞與紅原雞均屬地方肉蛋兼用型種類,覓食力極強,就巢性好強[2],但獨龍雞生活在高海拔、高寒地區,個體較小,而紅原雞生活于熱帶森林、次生竹林,海拔1 000 m以上的地方,個體較大。在顯著差異表達基因中,本研究發現PTEN和NCBP1顯著上調。卵巢是雞蛋形成的主要部位之一,蛋雞卵巢的生長發育水平與產蛋水平密切相關。PTEN參與了剪接體通路,該通路中BMP15基因已有研究表明其可調控蛋品質性狀[19]。本研究發現PTEN基因在獨龍雞卵巢中高表達,表明其可能通過剪接體通路參與了獨龍雞蛋品質的調控。
雞蛋的品質可以分為雞蛋的外在質量和雞蛋的內部質量[20]。外在質量包含蛋殼硬度、蛋質量、蛋形指數等指標;內在質量一般包含蛋黃品質(蛋黃色澤、蛋黃比重等),蛋清質量(蛋清高度、哈氏單位等)及其余技術指標(結構、味道和香氣、健康指數、血斑和肉斑、雞蛋的功用特點等)。Zhang等研究結果表明,NCBP1的缺失解除了許多基因的表達調控,降低了細胞增殖率[21]。NCBP1對于維持mRNA的表達至關重要,但其功能部分依賴于NCBP3,表明NCBP3可能在mRNA生物過程中充當RNA結合蛋白之間的橋梁,在mRNA輸出的上游起作用,RNA免疫沉淀試驗表明,NCBP2和NCBP3結合的RNA類型具有特異性[22]。mRNA和核糖體之間有多種結合位點,因此mRNA的轉錄和翻譯、蛋白質的生物合成都和核糖體密不可分。核糖體結構是蛋白質組成的最主要部分[23]。膨大部的核糖體首先組裝在mRNA上,然后沿著mRNA鏈翻譯,合成的蛋白質包裹在卵黃的外面,由于輸卵管的蠕動,包有蛋白質的卵也跟著旋轉前進,逐漸形成卵黃外的濃、稀蛋白層,濃蛋白層越高,對應的哈氏單位值越大,蛋品質就越好。在評價蛋品質高低的諸多性狀中,哈氏單位是非常關鍵的性狀之一。本研究發現NCBP1、NCBP2和NCBP3的表達顯著上調,這可能提示了獨龍雞與紅原雞相比,具有較高的蛋白比例、蛋白高度等。
本研究利用RNA測序技術分析獨龍雞和紅原雞的基因表達差異,并對獨龍雞和紅原雞卵巢組織的差異表達基因通過GO、KEGG、STRING等進行了富集分析,并篩選得到NCBP1、NCBP2、TXNRD1、PTEN、ENOX1等5個與抗性、產蛋性能和蛋品質相關的重要差異表達基因作為重要性狀候選基因。本研究為獨龍雞品種改良、后續基因功能選擇和種質特性等研究奠定了基礎。
參考文獻:
[1]李青青,李沫沫,陳粉粉,等. 獨龍雞生獨龍雞生長曲線擬合分析[J]. 中國家禽,2017,39(24):59-61.
[2]楊國強. 獨龍雞資源保護與開發利用的探索[J]. 云南畜牧獸醫,2017(4):16-18.
[3]張 勇,馮冬仙,李 貴. 獨龍江鄉獨龍雞的生產現狀及發展對策[J]. 安徽農業科學,2016,44(14):131-132.
[4]Haunshi S,Burramsetty A K,Kannaki T R,et al. Survivability,immunity,growth and production traits in indigenous and White Leghorn breeds of chicken[J]. British Poultry Science,2019,60(6):683-690.
[5]Gorshtein G,Grafinger O,Coppolino M G. Targeting SNARE-mediated vesicle transport to block invadopodium-based cancer cell invasion[J]. Frontiers in Oncology,2021,11:679955.
[6]賈保中. 硒對雞的營養作用[J]. 當代畜禽養殖業,2001(5):35-36.
[7]Bonora M,Patergnani S,Rimessi A,et al. ATP synthesis and storage[J]. Purinergic Signalling,2012,8(3):343-357.
[8]Wallin G,Kamerlin S C L,Aqvist J. Energetics of activation of GTP hydrolysis on the ribosome[J]. Nature Communications,2013,4:1733.
[9]李雙陽. 生物膜的結構與性質關系的模擬研究[D]. 北京:北京化工大學,2010:136.
[10]Weiss S,Jakobs J,Reemtsma T. Discharge of three benzotriazole corrosion inhibitors with municipal wastewater and improvements by membrane bioreactor treatment and ozonation[J]. Environmental Science & Technology,2006,40(23):7193-7199.
[11]Kozma L,Baltensperger K,Klarlund J,et al. The ras signaling pathway mimics insulin action on glucose transporter translocation[J]. Proceedings of the National Academy of Sciences of the United States of America,1993,90(10):4460-4464.[HJ2mm]
[12]匡鼎偉,吳永貴,陸福明. 葡萄糖轉運蛋白與糖尿病腎病[J]. 國外醫學:泌尿系統分冊,2000(3):109-112.
[13]Steffen A,Le Dez G,Poincloux R,et al. MT1-MMP-dependent invasion is regulated by TI-VAMP/VAMP7[J]. Current Biology,2008,18(12):926-931.
[14]Kean M J,Williams K C,Skalski M,et al. VAMP3,syntaxin-13 and SNAP23 are involved in secretion of matrix metalloproteinases,degradation of the extracellular matrix and cell invasion[J]. Journal of Cell Science,2009,122(22):4089-4098.
[15]Williams K C,Coppolino M G. SNARE-dependent interaction of Src,EGFR and β1 integrin regulates invadopodia formation and tumor cell invasion[J]. Journal of Cell Science,2014,127(Pt 8):1712-1725.
[16]Williams K C,McNeilly R E,Coppolino M G. SNAP23,Syntaxin4,and [JP+2]vesicle-associated membrane protein 7 (VAMP7) mediatetrafficking of membrane type 1-matrix metalloproteinase (MT1-MMP) during invadopodium formation and tumor cell invasion[J]. Molecular Biology of the Cell,2014,25(13):2061-2070.
[17]Gan L P,Zhao Y Z,Mahmood T,et al. Effects of dietary vitamins supplementation level on the production performance and intestinal microbiota of aged laying hens[J]. Poultry Science,2020,99(7):3594-3605.
[18]黃 晶,林伯全. 富硒酵母的營養生理作用及其在雞生產中的應用[J]. 廣東飼料,2013,22(8):32-34.
[19]李洪林,李 維,張福平,等. 思州雞BMP15基因第1外顯子多態性及其與蛋品質的關聯分析[J]. 中國畜牧獸醫,2019,46(12):3690-3697.
[20]夏佳豪. 汶上蘆花雞蛋品質及其候選基因遺傳效應分析[D]. 泰安:山東農業大學,2018:88.
[21]Zhang H J,Wang A,Tan Y L,et al. NCBP1 promotes the development of lung adenocarcinoma through up-regulation of CUL4B[J]. Journal of Cellular and Molecular Medicine,2019,23(10):6965-6977.
[22]Miura P,Coriati A,Bélanger G,et al. The utrophin A 5′-UTR drives cap-independent translation exclusively in skeletal muscles of transgenic mice and interacts with eEF1A2[J]. Human Molecular Genetics,2010,19(7):1211-1220.
[23]Zhao H Y,Wang J S,Qu Y F,et al. Transcriptomic and proteomic analyses of a new cytoplasmic male sterile line with a wild Gossypium bickii genetic background[J]. BMC Genomics,2020,21(1):859.
收稿日期:2022-02-23
基金項目:國家自然科學基金(編號:31902152);云南省教育廳科學研究基金項目(研究生類)(編號:2021Y267);云南省教育廳科學研究基金教師項目(編號:2018JS333);西南林業大學博士科研啟動基金(編號:112119);云南省科技廳基礎研究項目(編號:2019FD06)。
作者簡介:丁玥竹(1997—),女,黑龍江雞西人,碩士研究生,主要研究方向為動物遺傳育種。E-mail:752958200 @qq.com。
通信作者:劉莉莉,博士,副教授,碩士生導師,研究方向為動物遺傳育種研究。E-mail:liulily0518@163.com。
