近現代中國水稻育種歷程
2023-07-10 05:56:14李勝男劉偉胡興明趙良俠
江蘇農業科學 2023年11期
關鍵詞:水稻
李勝男 劉偉 胡興明 趙良俠

摘要:栽培水稻起源于我國,我國以稻米為主食有著超過3 000年的歷史,水稻研究的歷史也十分悠久。回顧我國一百多年來水稻育種歷程,育種手段從簡單的品種比較發展為以科學理論為支撐的純系育種、雜交育種和生物育種等,期間取得了矮化育種、三系法育種、兩系法育種和超級稻育種等重大突破。文章根據各個時期的發展特點,將該歷程總結為3個發展時期,分別為科學育種初期、獨立發展時期和逐步領先時期,對各個時期的主要育種手段及所取得的成就進行了總結,列舉了我國近現代水稻育種歷程的一些關鍵基因,并對水稻育種新技術及今后我國水稻育種發展方向進行了討論和展望。
關鍵詞:水稻;育種歷程;矮化育種;超級稻;生物育種;基因編輯
中圖分類號:S511.03文獻標志碼:A文章編號:1002-1302(2023)11-0020-07
“國以民為本,民以食為天”。目前超5成的世界人口以稻米為主要食物來源,我國作為全球第一水稻生產大國,在水稻育種方面有著突出貢獻。在過去的一百多年里,我國從落后到效仿到獨立創新并逐漸領先全球的水稻育種進程,這是一代代育種家辛勤耕耘的結果。回顧我國一百多年來水稻育種的發展歷程以及對世界水稻事業的貢獻,可將我國近現代水稻育種歷程分為3個階段:科學育種初期、獨立發展時期和逐步領先時期(表1)。
1科學育種初期(清末民初至1925年)
1898年戊戌變法后,清政府提出了一系列農業改良措施,包括成立農政機構、農業試驗場和農業學校,同時翻譯西方書籍,傳播科學新思想,推進農業改革,這為我國當前水稻科學育種事業的開展奠定了基礎。20世紀初,我國大部分省份如四川、廣東、云南等都建立了農事試驗場和農事學校,并已經開始搜集國內外水稻品種,開展品種比較試驗。1914年,北京政府便要求各省將轄區內稻種送到農商部,并按照稻種品質優劣將稻種分為4個等級,再將優質稻種分發各省,用作來年試種,這是我國政府第1次組織的全國性水稻品種鑒定[1]。隨著西方遺傳學理論在我國的傳播及世界農業育種技術等各類科學技術的發展,我國開展了具有開創性意義的育種工作。1919年,有我國近代運用遺傳學原理開展水稻育種的具有初創者之稱的原頌周先生,在南京高等師范學校的試驗農場,對當地栽培的水稻品種以及從各省征集的數十個水稻良種進行品種比較試驗和選種改良,并于1924年育成了改良江寧洋秈和改良東莞白2個優良純系品種,在長江中下游地區推廣效果良好[2]。在此之后,我國南方地區的許多農事試驗場和農事學校都陸續開展了水稻育種工作。
2獨立發展時期(1925年至20世紀50年代)
20世紀20年代,純系育種和雜交育種等作物育種手段興起,全國各地育種規模迅速擴大,丁穎、趙連芳、周拾祿先生等水稻育種家也陸續學成回國。1930年中央農業實驗所和全國稻麥改進所成立,我國水稻育種事業正式進入利用現代科技方法的重要階段。國際上最先進的育種技術已在我國大多數育種機構應用。與此同時,我國各研究所開始進行獨立探索和研究,率先開展的矮化育種更是為解決人類的溫飽問題作出了重大貢獻。
2.1純系育種
純系育種法是近代我國水稻育種的重要手段。1925年國立廣東大學(中山大學前身)率先開展了水稻純系育種,我國最初采用洛夫的穗行純系育種法,后經實踐趙連芳、丁穎等育種家結合我國實際情況加以改進,開辟了適合我國本土的純系育種法,中大帽子頭、南特號、矮腳南特、勝利秈等優良品種均是用此法選育而來[3]。我國于1929年育成近代第1個大規模推廣的優良品種——中大帽子頭,全國推廣面積超1萬hm2。1934年江西省農業試驗場從農家品種鄱陽早選得變異單穗,育成適應性廣、產量高、高抗普矮病和黃矮病的秈型常規水稻,后經提純復壯適應性增強,1958—1962年全國推廣面積均在 400萬hm2左右。
2.2雜交育種
我國的水稻雜交育種起步整體上略晚于純系育種,后隨著雜種優勢的不斷發掘,水稻雜交育種逐漸成為主流。國內以廣東省率先開始水稻的雜交育種,丁穎院士開辟了我國水稻雜交育種技術及理論,1926年丁穎發現了竹占與野生稻自然雜交的雜交種子,并經過幾代選育,培育出中山1號,開創了雜交稻與野生稻選育品種的先河。20世紀30年代,各地農事機構才陸續開展栽培稻與野生稻及栽培稻之間的雜交工作,有目的地選配不同親本進行雜交、自交、回交等,結合雙親的優良性狀培育出大批優良品種。1951年,沈陽農學院的楊守仁等已經開始了秈粳稻雜交工作,并采用系譜法選育優良品種。雜交育種事業的重大轉折在1963年,在此之前采用雜交育種法培育的品種基本都是常規稻,直到20世紀60年代雜交水稻思想和技術被提出,1964年袁隆平在我國開創了雜交水稻研究的先河,1973年實現三系配套成功,在國際上取得矚目成就。
2.3矮化育種
矮化育種是水稻的第1次綠色革命,我國于19世紀50年代中期開始水稻矮稈育種工作,矮腳南特、矮仔占2個矮稈品種是我國矮化育種的重要矮源。1941年,廣西壯族自治區玉林市容縣華僑甘利南,從南洋帶回稻種,見其長得特別矮,稱其為矮仔占。1953年,一場風雨,眾多品種皆倒伏,唯矮仔占屹立不倒,且產量竟高達444 kg/667 m2,引起廣泛關注,各地前來索要稻種,一場矮化育種革命就此展開。“中國半矮稈水稻之父”黃耀祥先生開辟了我國水稻矮化育種事業。1956年,廣東省農業科學院黃耀祥先生用矮仔占4號與廣場13號雜交,于1959年育成廣場矮品種,隨即大面積推廣,后相繼育成推廣珍珠矮、廣解9號、廣陸矮4號及窄葉青8號等優良高產品種[4]。1956年,廣東省汕頭市潮陽縣農民育種家洪群英、洪春利2人在南特16號田內,經純系育種選育,育成了全國第1個水稻高產矮稈品種矮腳南特,成為當時的主栽品種,并成為矮化育種革命的重要矮源,是具有劃時代意義的品種。我國臺灣省臺中區農業改良場洪秋增利用低腳烏尖與菜園種雜交,于1956年育成臺中在來1號,并在1960年投入推廣。矮稈品種廣場矮等的育成,是我國水稻矮化育種研究的重大突破,比菲律賓國際水稻研究所1966年育出的矮稈品種IR8早7年[5]。我國的矮化育種事業,實現了水稻產量的第1次大飛躍,也是我國運用現代遺傳學理論獨立開展水稻研究的重大突破。
3逐步領先時期(20世紀60年代至今)
三系的配套成功,標志著我國水稻育種事業率先突破瓶頸,該技術還作為我國第1項農業高新技術出口美國。后來我國在兩系法、超級稻育種、生物育種等方面也取得傲人成就。該時期,我國水稻育種由學習效仿轉變為自主創新,卓有成就并在各國推廣,逐步領先全球。此外,我國起步于20世紀60年代的誘變育種、70年代的細胞工程育種、80年代的無融合生殖(一系法)、90年代的分子育種等水稻育種技術現均取得了不凡成就。這些豐富的水稻育種手段,為我國的水稻育種事業帶來了十足的活力,下文筆者選取了我國艱辛水稻育種歷程中最具里程碑意義的成就展開詳述。
3.1三系雜交稻
1961年袁隆平院士注意到天然雜交水稻的穗大粒多,經多次試驗確定水稻較于異花授粉植物雜種優勢同樣明顯,并于1964年開始研究培育雜交水稻,同時期美國、日本和國際水稻研究所也開展了水稻雜種優勢利用相關研究,但因無法解決雜交稻制種問題,無奈終止。當時,袁隆平院士選用的技術路線是三系法,1964和1965年通過數以萬計的逐穗檢查,在大田中找到6株雄性不育水稻,但幾千個雜交組合均不理想,后調整策略選擇野生不育稻。功夫不負苦心人,在袁隆平思想路線指導下,李必湖及馮克珊于1970年在海南三亞一片野生稻群體中找到了野生稻天然雄性不育株(野敗),成為了三系配套成功的突破口[6]。此后,全國18個單位共同攻艱,利用野敗進行廣泛測交、輪交和篩選,1972年顏龍安等育成二九矮1號及同型保持系,1973年廣西農學院張先程找到合適的恢復系,秈型三系終得配套成功,袁隆平研制出第1個雜交水稻南優2號,顏龍安培育出強優勢組合汕優2號,均得到大面積推廣。李錚友等于1969年育成滇型紅帽纓粳稻不育系,1975年雜交粳稻三系配套成功[7]。1980年,由謝華安研發的汕優63是我國推廣面積最大的雜交水稻,截至2020年,我國推廣面積最大的雜交稻還有汕優64、威優64、岡優22、汕優2號、汕優6號、威優6號、Ⅱ優838、D優63和汕優10號(國家水稻數據中心:http://www.ricedata.cn/variety/)。我國雜交水稻三系配套的成功,不僅讓水稻產量上升到一個新平臺,也為解決我國糧食短缺問題作出了重大貢獻。
3.2兩系雜交稻
20世紀80年代中期,三系雜交稻的產量及優勢水平一直處于徘徊局面,同時不育系繁殖和制種成本相對也很高,光溫敏雄性不育為雜交稻育種帶來轉機。1973年石明松在晚粳品種農墾58中,發現天然不育株,經多年播種鑒定,發現其長日高溫不育,短日低溫可育,可作一系兩用。1981年,石明松發表《晚粳自然兩用系選育及應用初報》一文,并稱之為兩用系(現稱為光溫敏不育系),采用此系不需要保持系,制種較三系法簡單,省力省田[8]。1980年起湖北省農業科學院開始研究該材料不育的原因,1985年,綜合各方研究,普遍認同調控其育性的主要因素是光照長度,溫度起輔助作用,并命名該材料為農墾58S[9-10]。自農墾58S發現后,全國掀起兩系法研究熱潮,兩系法雜交稻被列為國家重點攻關的課題。1987年,鄧華鳳在秈稻三系育種材料中找到并育成光敏核不育水稻安農S1,1988年[11],福建農學院楊仁崔在IR54中發現5460S不育系,這些不育系的發現為兩系雜交水稻的研究提供了材料[12]。為了應對異常氣溫的影響,1992年,袁隆平提出以不育起點溫度作為光敏不育的關鍵指標,并提出選育水稻光溫敏不育系的技術策略。此后我國兩系雜交稻育種方法逐漸形成體系,直至今日,我國兩系法雜交稻技術依然領先全球。2001年后,兩系雜交稻進入迅速發展時期,過審品種和推廣面積大幅提升,推廣面積占雜交稻的總面積由1996年的不到1%上升到2013年的33.59%[13]。截至2021年,省級以上審定的兩系雜交稻品種有 2 575 個(國家水稻數據中心,https://www.ricedata.cn/variety/)。兩系法雜交稻的提出在提升了雜交稻制種效率的同時,增強了雜種優勢,由品種間的雜種優勢利用上升到亞種間的優勢利用,是我國繼三系法水稻雜交育種后的又一次大突破,占據了雜交稻的半壁江山。
3.3超級稻
超級稻引起廣泛關注始于1994年國際水稻研究所培育的一批新株型超高產水稻品種,隨后我國農業農村部(原農業部)于1996年啟動中國超級稻育種計劃,力求我國水稻產量更上一層樓。我國在國際上所提出的理想株型和高產栽培技術結合培育超級稻的基礎上,提出理想株型與強雜種優勢結合的超級稻理念,選育高產、優質、抗逆的超級稻。超級稻計劃啟動后,各地因地制宜,根據不同氣候特征提出了適合當地栽培的超級稻理想株型。關于理想株型,我國學者楊守仁等在19世紀70年代便開展了水稻理想株型育種研究工作,并提出東北粳稻區短枝立葉,大穗直穗株型,黃耀祥等提出東南地區叢生早長型株型,周開達等提出華南稻區重穗型株型,袁隆平提出高冠層、矮穗層、中大層理想株型,還提出了通過適當增加株高,提高莖稈壁厚度,從而增加其生物量,提高收獲指數[14-18]。關于雜種優勢利用,水稻雜種優勢以秈粳亞種間雜交最優,增產潛力最大,但秈粳雜交有不親和性,結實率不甚理想,經過近10年的研究,以寧波市農業科學研究院馬榮榮先生為代表的育種家通過利用廣親和基因及具有混合親緣的中間材料使亞種間雜種優勢利用取得顯著進展,育成一大批秈粳稻雜交組合。
我國的超級稻計劃指標共分為4期:第1期(1996—2000年),產量700 kg/667 m2;第2期(2001—2005年),產量800 kg/667 m2;第3期(2006—2015年),產量900 kg/667 m2;第4期(2016—2020年),產量達1 000 kg/667 m2。4期計劃指標全部如期完成,甚至在2014年就順利突破 1 000 kg/667 m2,提前6年完成第4期指標。袁隆平接著又向農業農村部(原農業部)建議提出了第5期指標,2020年實現16 t/hm2的超高產指標[19]。2012年,超級稻就已在全國稻區無障礙推廣,據農業農村部數據,截至2018年底,全國超級稻累計推廣應用1億hm2,占水稻種植面積的30%,截至2021年,全國經農業農村部認定達到推廣要求的超級稻品種135個。我國超級稻研究為水稻單位面積產量的提升帶來了源源動力,除超級稻外,張啟發提出的綠色超級稻構想,在保障高產優質的前提下,借助分子輔助育種等手段提高品種抗性及養分利用率,做到基本不打農藥,少施肥,抗旱節水,這對解決人口對資源需求增長同環境可持續發展的矛盾有重大意義[20]。
3.4水稻生物育種
生物育種是指在經典育種手段的基礎上,利用遺傳學、分子生物學和細胞生物學等學科,并結合現代生物工程技術等方法培育新品種的過程,水稻生物育種現階段主要包括分子標記育種、轉基因育種和水稻分子模塊設計育種。隨著生物育種技術的發展,在水稻育種發展史上起重要作用的基因也被發掘出來,如矮化育種的關鍵基因SD1,三系法關鍵基因細胞質雄性不育基因WA352和恢復基因Rf3、Rf4,主導兩系法育種的光溫敏核不育基因PMS3和TMS55[21-25]。目前我國水稻功能基因發掘的主要方法有DNA標記技術、基因定位、圖位克隆等,可通過結合農藝性狀QTLs的鑒定,利用基因編輯技術和轉基因技術,創造優勢等位基因,實現水稻的定向性狀改良,水稻生物育種技術極大地提高了育種效率和育種選擇方向性。
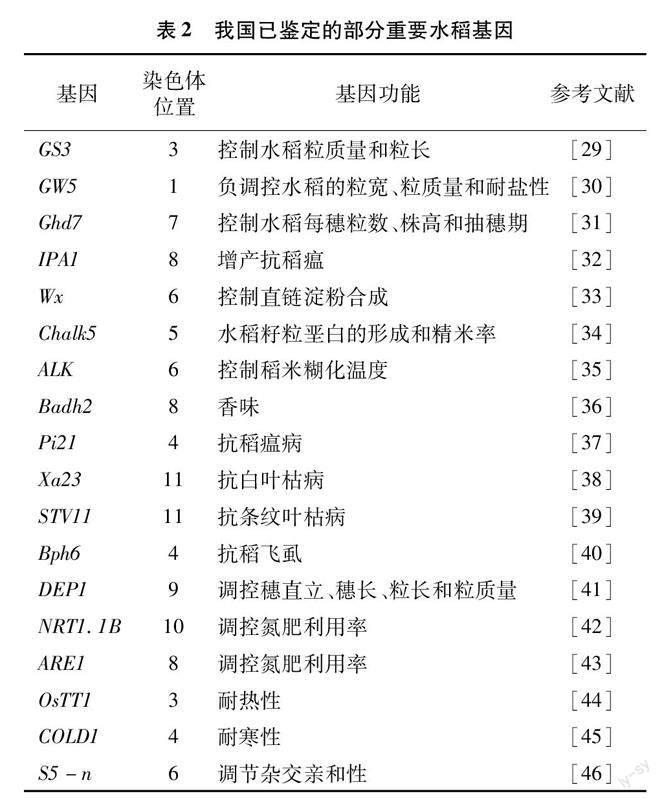
我國利用生物技術開展水稻育種起始于20世紀80年代,1990年中國科學院植物生理生態研究所便克隆了稻米品質基因Wx[26]。1995年華中農業大學張啟發團隊開始了轉基因抗蟲水稻的研發工作,1999年通過農業農村部(原農業部)成果鑒定,直到2009年抗蟲轉基因水稻華恢1號和Bt汕優63才獲得農業農村部頒發的全國第1張轉基因水稻安全證書,并批準在湖北省種植。2002年秈稻9311和粳稻日本晴全基因組測序完成,正式打開了水稻生物育種的大門[27-28]。隨著更多水稻品種全基因組測序完成,以及基因編輯技術的精準化和簡便化,我國各農業科研機構和院校均在生物育種工作中投入了大量精力。當前我國水稻生物育種研究如日中天,發掘并克隆了大批水稻功能基因,并在抗病蟲、抗稻瘟、產量品質改良等方面均有所成就。至今,我國已發掘利用水稻重要功能基因有產量基因(GS3、GW5、Ghd7、IPA1等)、品質基因(Wx、Chalk5、ALK、Badh2等)、抗病抗蟲基因(Pi21、Xa23、STV11、Bph6等)、營養高效利用基因(DEP1、NRT1.1B、ARE1等)、抗非生物脅迫基因(OsTT1、COLD1等)和秈粳雜種優勢利用基因S5-n[29-46]等(表2)。除此之外,我國科學家還開發了多種基因編輯工具,2015年劉耀光課題組開發了CRISPR/Cas9多靶點編輯載體系統,2020年高彩霞課題組開發了胞嘧啶堿基編輯器,并在水稻中成功應用[47-48]。
根據funRiceGenes數據顯示,截至2022年3月,已克隆的水稻基因達4 100多個,越來越多功能基因的發掘意味著水稻功能基因組學在不斷完善,為水稻分子水平的研究開拓了廣闊的前景。在現代生物科技的推動下,生物育種是必然趨勢。
4水稻育種新技術和發展內容
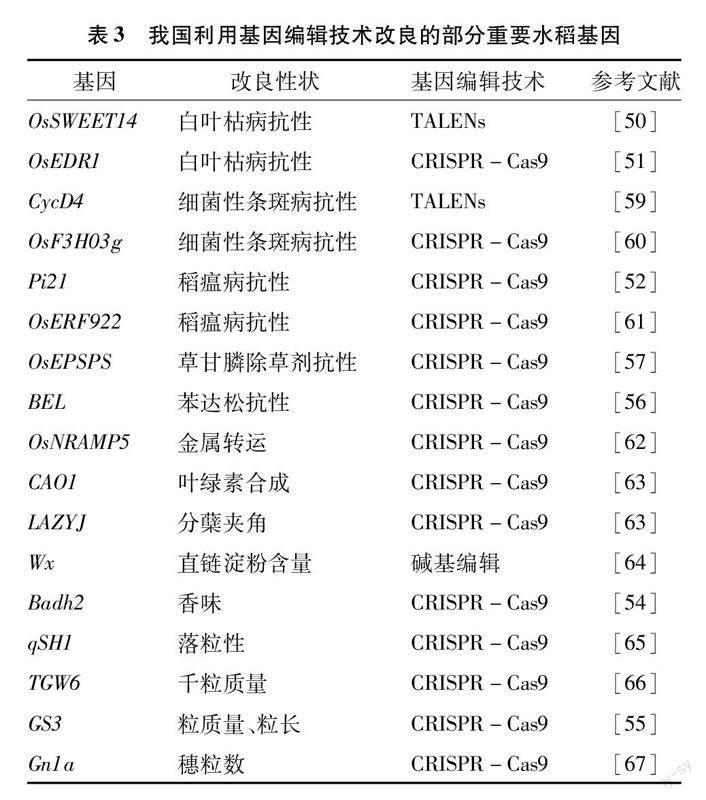
基因編輯技術主要包括鋅指核酸酶(ZFNs)、轉錄激活因子效應物核酸酶(TALENs)和成簇規律間隔短回文重復(CRISPR-Cas9)3個系統及堿基編輯技術[49]。其中鋅指核酸酶系統因特異性低、操作繁瑣在水稻性狀改良中并未發揮顯著作用,而CRISPR-Cas9系統以其簡便、高效的特點成為現階段廣泛應用的手段。目前我國科學家利用基因編輯技術定向改良品種性狀已經取得初步成果(表3)。如利用基因編輯技術對OsSWEET14、OsEDR1、Pi21等進行基因編輯提高水稻抗病性,對Wx、Badh2、GS3等進行基因編輯提高稻米品質,編輯BEL、OsEPSPS等基因增強水稻抗除草劑特性等[50-57]。2019年,Wang等利用CRISPR-Cas9系統,將與水稻減數分裂相關的PAIR1、REC8、OSD1和MTL 4個基因敲除后,先中斷減數分裂,再誘導孤雌生殖,育成克隆種子,基因編輯技術為無融合生殖帶來了曙光[58]。改良品種的關鍵在于創造有利變異,而以CRISPR-Cas9系統為代表的基因編輯技術便是現階段定向且高效創造有利變異的關鍵手段。水稻功能基因組學的完善和基因編輯技術的創新與成熟,為創造優質種質資源帶來了巨大的發展潛力。但若要實現高效率、大批量的發掘利用水稻優勢基因,僅靠基因編輯技術的進步是不夠的,還需要測序技術、基因組學分析、蛋白質組學分析等檢測手段的輔助。此外,國家基因編輯監管政策的不斷完善將推動生物育種的全面發展,為我國水稻育種事業帶來新突破!
5討論與展望
近現代我國水稻育種歷程是艱辛的,縱觀這一百多年的水稻育種歷程,無時不提醒著我們要始終保持謙虛敬畏,要與時俱進,不能固步自封,只有站在前人已有的研究基礎上,不斷汲取全球最新知識,拓寬思維,勇于探索創新,才能不斷突破,行穩致遠。這一路我們以純系育種和雜交育種為技術基礎,降低株高實現第1次產量飛躍,三系、兩系配套成功是我國稻米產量的第2次飛躍,將優質株型與強雜種優勢結合培育超級稻又實現了第3次產量飛躍。水稻生物育種是實現我國第4次產量飛躍的關鍵,這要求我們在經典育種手段的基礎上,利用現有的基因編輯、分子標記、遺傳轉化等手段,運用計算機、物聯網、大數據、人工智能等現代高科技手段,通過分子標記輔助選擇基因聚合育種,從而育成高產、優質、多抗、廣適的水稻新品種。水稻生物育種的不斷發展將加快新品種選育效率,加快制種速度,提高制種產量并降低制種成本,豐富種質資源,為實現我國農業現代化,全面推進鄉村振興,解決種業“卡脖子”難題,提供重要的智力支撐,讓智慧育種走進現實,為國家糧食安全保駕護航。
在全球經濟一體化時代,挑戰與機遇并存。全球疫情與局部動蕩的存在,讓整個社會充滿不確定性。我們應搶抓戰略機遇期,更好地繼承和發揚前人自力更生、敢為人先的優良傳統,沿著先驅者的足跡,積極探索,作出無愧于時代的貢獻。雖然我國水稻育種已經進入發展新階段,但還存在許多亟待解決的問題,在前沿科學理論方面還欠深入,在關鍵功能基因挖掘鑒定和突破性育種技術上還存在不足,存在優異種質資源缺乏,育種成果滯后于市場需求、轉化率低等問題。水稻育種新階段機遇與挑戰并存,這要求我們深入探究多維組學,研發更高效、更精準的新型生物技術,提高科技競爭力,加大種質資源的搜集、鑒定、評價、利用和保護力度,加快育種新技術的開發、推廣和應用,大力研發順應市場需求的突破性新品種,推動育種成果快速轉化應用,加快實現民族種業的提檔升級,增強核心競爭力,全力推進現代化種業強國的建設步伐,提高我國民族種業的全球市場占有率和國際話語權,通過頂層設計和自主創新,發揮體制優勢,促進全國大協作,盡快躍居行業戰略制高點,實現我國水稻育種事業在更高水平、更深層次、更大平臺上的跨越式突破。
參考文獻:
[1]夏如冰. 北洋政府時期的農政機構與農業政策(1912—1928年)[J]. 南京農業大學學報(社會科學版),2003,3(3):90-95.
[2]曹幸穗. 從引進到本土化:民國時期的農業科技[J]. 古今農業,2004(6):45-53.
[3]夏如兵. 近代中國水稻育種科技的發展歷程[J]. 農業考古,2010(4):34-38,45.
[4]程式華. 中國水稻育種百年發展與展望[J]. 中國稻米,2021,27(4):1-6.
[5]咸金山. 我國近代稻作育種事業述評[J]. 中國農史,1988,7(1):54-63.
[6]袁隆平. 我在雜交水稻方面所做的工作[J]. 中國科技獎勵,2001(1):14-19.
[7]李振宇,吳建利. 我國三系雜交粳稻育種研究的現狀與展望[J]. 雜交水稻,1991(增刊2):13-16.
[8]石明松. 晚粳自然兩用系選育及應用初報[J]. 湖北農業科學,1981,20(7):1-3.
[9]馮云慶,王長義,李全新. 湖北長日核不育水稻的研究與利用[J]. 作物學報,1985,11(4):227-234.
[10]元生朝,張自國,許傳楨. 光照誘導湖北光敏感核不育水稻育性轉變的敏感期及其發育階段的探討[J]. 作物學報,1988,14(1):7-13.
[11]鄧華鳳,舒福北,袁定陽. 安農S-1的研究及其利用概況[J]. 雜交水稻,1999,14(3):3-5.
[12]楊仁崔,王乃元,梁康逕,等. 秈稻溫敏核不育5460S的研究[J]. 福建農學院學報,1993,22(2):135-140.
[13]牟同敏. 中國兩系法雜交水稻研究進展和展望[J]. 科學通報,2016,61(35):3761-3769.
[14]楊守仁,張龍步,沈钖英,等. 三十六年來秈粳稻雜交育種的研究及發展[J]. 沈陽農業大學學報,1987,18(3):3-9.
[15]徐正進,陳溫福,張文忠,等. 北方粳稻新株型超高產育種研究進展[J]. 中國農業科學,2004,37(10):1407-1413.
[16]黃耀祥,林青山. 水稻超高產、特優質株型模式的構想和育種實踐[J]. 廣東農業科學,1994,21(4):1-6.
[17]周開達,馬玉清,劉太清,等. 雜交水稻亞種間重穗型組合選育——雜交水稻超高產育種的理論與實踐[J]. 四川農業大學學報,1995,13(4):403-407.
[18]袁隆平. 超級雜交水稻的現狀和展望[J]. 糧食科技與經濟,2003,28(1):2-3.
[19]袁隆平. 中國超級雜交稻育種技術的未來展望[J]. 中國鄉村發現,2015(1):18-21.
[20]張啟發. 綠色超級稻培育的設想[J]. 分子植物育種,2005,3(5):601-602.
[21]Sasaki A,Ashikari M,Ueguchi-Tanaka M,et al. Green revolution:a mutant gibberellin-synthesis gene in rice[J]. Nature,2002,416(6882):701-702.
[22]Luo D P,Xu H,Liu Z L,et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice[J]. Nature Genetics,2013,45(5):573-577.
[23]Yao F,Xu C,Yu S B,et al. Mapping and genetic analysis of two fertility restorer loci in the wild-abortive cytoplasmic male sterility system of rice (Oryza sativa L.)[J]. Euphytica,1997,98:183-187.
[24]梅明華,陳亮,章志宏,等. 農墾58S光敏不育基因突變位點的確定及pms3區間的進一步作圖[J]. 中國科學(C輯),1999,29(3):310-315.
[25]Wang Y G,Xing Q H,Deng Q Y,et al. Fine mapping of the rice thermo-sensitive genic male-sterile gene tms5[J]. Theoretical and Applied Genetics,2003,107(5):917-921.
[26]Wang Z Y,Wu Z L,Xing Y Y,et al. Nucleotide sequence of rice waxy gene[J]. Nucleic Acids Research,1990,18(19):5898.
[27]Yu J,Hu S N,Wang J,et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica)[J]. Science,2002,296(5565):79-92.
[28]Goff S A,Ricke D,Lan T H,et al. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica)[J]. Science,2002,296(5565):92-100.
[29]Mao H L,Sun S Y,Yao J L,et al. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice[J]. Proceedings of the National Academy of Sciences of the United States of America,2010,107(45):19579-19584.
[30]Weng J F,Gu S H,Wan X Y,et al. Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight[J]. Cell Research,2008,18(12):1199-1209.
[31]Xue W Y,Xing Y Z,Weng X Y,et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice[J]. Nature Genetics,2008,40(6):761-767.
[32]Wang S S,Wu K,Qian Q,et al. Non-canonical regulation of SPL transcription factors by a human OTUB1-like deubiquitinase defines a new plant type rice associated with higher grain yield[J]. Cell Research,2017,27(9):1142-1156.
[33]Huang L C,Li Q F,Zhang C Q,et al. Creating novel Wx alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system[J]. Plant Biotechnology Journal,2020,18(11):2164-2166.
[34]Li Y B,Fan C C,Xing Y Z,et al. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice[J]. Nature Genetics,2014,46(4):398-404.
[35]Zhang G Y,Cheng Z J,Zhang X,et al. Double repression of soluble starch synthase genes SSⅡa and SSⅢa in rice (Oryza sativa L.) uncovers interactive effects on the physicochemical properties of starch[J]. Genome,2011,54(6):448-459.
[36]Lorieux M,Petrov M,Huang N,et al. Aroma in rice:genetic analysis of a quantitative trait[J]. Theoretical and Applied Genetics,1996,93(7):1145-1151.
[37]Fukuoka S,Okuno K.QTL analysis and mapping of pi21,a recessive gene for field resistance to rice blast in Japanese upland rice[J]. Theoretical and Applied Genetics,2001,103(2/3):185-190.
[38]Wang C L,Fan Y L,Zheng C K,et al. High-resolution genetic mapping of rice bacterial blight resistance gene Xa23[J]. Molecular Genetics and Genomics:MGG,2014,289(5):745-753.
[39]Wang Q,Liu Y Q,He J,et al. STV11 encodes a sulphotransferase and confers durable resistance to rice stripe virus[J]. Nature Communications,2014,5:4768.
[40]Guo J P,Xu C X,Wu D,et al. Bph6 encodes an exocyst-localized protein and confers broad resistance to planthoppers in rice[J]. Nature Genetics,2018,50(2):297-306.
[41]Huang X Z,Qian Q,Liu Z B,et al. Natural variation at the DEP1 locus enhances grain yield in rice[J]. Nature Genetics,2009,41(4):494-497.
[42]Zhang J Y,Liu Y X,Zhang N,et al. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice[J]. Nature Biotechnology,2019,37(6):676-684.
[43]Wang Q,Nian J Q,Xie X Z,et al. Genetic variations in ARE1 mediate grain yield by modulating nitrogen utilization in rice[J]. Nature Communications,2018,9(1):735.
[44]Li X M,Chao D Y,Wu Y,et al. Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice[J]. Nature Genetics,2015,47(7):827-833.
[45]Luo W,Huan Q,Xu Y Y,et al. Integrated global analysis reveals a vitamin E-vitamin K1 sub-network,downstream of COLD1,underlying rice chilling tolerance divergence[J]. Cell Reports,2021,36(3):109397.
[46]Yang J Y,Zhao X B,Cheng K,et al. A killer-protector system regulates both hybrid sterility and segregation distortion in rice[J]. Science,2012,337(6100):1336-1340.
[47]Ma X L,Zhang Q Y,Zhu Q L,et al. A robust CRISPR/Cas9 system for convenient,high-efficiency multiplex genome editing in monocot and dicot plants[J]. Molecular Plant,2015,8(8):1274-1284.
[48]Wan X Y,Weng J F,Zhai H Q,et al. Quantitative trait loci (QTL) analysis for rice grain width and fine mapping of an identified QTL allele gw-5 in a recombination hotspot region on chromosome 5[J]. Genetics,2008,179(4):2239-2252.
[49]Komor A C,Kim Y B,Packer M S,et al. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage[J]. Nature,2016,533(7603):420-424.
[50]Li T,Liu B,Spalding M H,et al. High-efficiency TALEN-based gene editing produces disease-resistant rice[J]. Nature Biotechnology,2012,30(5):390-392.
[51]武廣珩,傅仙玉. 利用CRISPR/Cas9技術編輯水稻負調控抗病基因OsEDR1及基因功能分析[J]. 應用與環境生物學報,2019,25(6):1375-1380.
[52]王芳權,范方軍,李文奇,等. 利用CRISPR/Cas9技術敲除水稻Pi21基因的效率分析[J]. 中國水稻科學,2016,30(5):469-478.
[53]Zeng D C,Liu T L,Ma X L,et al. Quantitative regulation of Waxy expression by CRISPR/Cas9-based promoter and 5′UTR-intron editing improves grain quality in rice[J]. Plant Biotechnology Journal,2020,18(12):2385-2387.
[54]邵高能,謝黎虹,焦桂愛,等. 利用CRISPR/CAS9技術編輯水稻香味基因Badh2[J]. 中國水稻科學,2017,31(2):216-222.
[55]沈蘭,李健,付亞萍,等. 利用CRISPR/Cas9系統定向改良水稻粒長和穗粒數性狀[J]. 中國水稻科學,2017,31(3):223-231.
[56]Xu R F,Li H,Qin R Y,et al. Gene targeting using the Agrobacterium tumefaciens-mediated CRISPR-Cas system in rice[J]. Rice,2014,7(1):5.
[57]Sun Y W,Zhang X,Wu C Y,et al. Engineering herbicide-resistant rice plants through CRISPR/Cas9-mediated homologous recombination of acetolactate synthase[J]. Molecular Plant,2016,9(4):628-631.
[58]Wang C,Liu Q,Shen Y,et al. Clonal seeds from hybrid rice by simultaneous genome engineering of meiosis and fertilization genes[J]. Nature Biotechnology,2019,37(3):283-286.
[59]Cai L L,Cao Y Y,Xu Z Y,et al. A transcription activator-like effector Tal7 of Xanthomonas oryzae pv. oryzicola activates rice gene Os09g29100 to suppress rice immunity[J]. Scientific Reports,2017,7(1):5089.
[60]Wu T,Zhang H M,Yuan B,et al. Tal2b targets and activates the expression of OsF3H03g to hijack OsUGT74H4 and synergistically interfere with rice immunity[J]. The New Phytologist,2022,233(4):1864-1880.
[61]Wang F J,Wang C L,Liu P Q,et al. Enhanced rice blast resistance by CRISPR/Cas9-targeted mutagenesis of the ERF transcription factor gene OsERF922[J]. PLoS One,2016,11(4):e0154027.
[62]Tang L,Mao B G,Li Y K,et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield[J]. Scientific Reports,2017,7(1):14438.
[63]Miao J,Guo D S,Zhang J Z,et al. Targeted mutagenesis in rice using CRISPR-Cas system[J]. Cell Research,2013,23(10):1233-1236.
[64]Huang X R,Su F,Huang S,et al. Novel Wx alleles generated by base editing for improvement of rice grain quality[J]. Journal of Integrative Plant Biology,2021,63(9):1632-1638.
[65]盛夏冰,譚炎寧,孫志忠,等. 利用CRISPR/Cas9基因組編輯技術定向降低水稻落粒性[J]. 中國農業科學,2018,51(14):2631-2641.
[66]王加峰,鄭才敏,劉維,等. 基于CRISPR/Cas9技術的水稻千粒重基因tgw6突變體的創建[J]. 作物學報,2016,42(8):1160-1167.
[67]Huang L Y,Zhang R,Huang G F,et al. Developing superior alleles of yield genes in rice by artificial mutagenesis using the CRISPR/Cas9 system[J]. The Crop Journal,2018,6(5):475-481.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
