外來入侵雜草田菁的構件生物量分配與異速生長分析
2023-07-10 23:10:24鄧美菁張忠華莫秋麗胡聰徐超昊胡剛
雜草學報 2023年1期
鄧美菁 張忠華 莫秋麗 胡聰 徐超昊 胡剛

摘要:田菁(Sesbania cannabina)是我國華南、華中和華東等地區危害嚴重的外來入侵雜草,通過測定其構件(根、莖、葉、花、果)生物量和植株高度等數據,分析其構件生物量分配特征及其與異速生長的關系,以期掌握該入侵雜草的生長發育規律。結果表明:田菁的構件生物量分配關系表現為莖>葉>果>根>花,而其變異系數從大到小為果>莖>葉>根>花,其地上與地下以及生殖與營養構件間的生物量均表現出極顯著的差異性。此外,田菁各構件之間以及株高與構件間均為異速生長關系,株高與各構件生物量間呈極顯著正相關性。田菁種群各構件間的異速生長關系以及較高的地上構件生物量與其較強的入侵性有密切聯系,該結果對評估田菁的入侵機制具有重要價值。
關鍵詞:田菁;外來入侵雜草;構件生物量分配;異速生長
中圖分類號:Q944;S451文獻標志碼:A文章編號:1003-935X(2023)01-0035-08
Biomass Partitioning in Modules and Allometric Growth of Alien Invasive Weed Sesbania cannabina
DENG Mei-jing, ZHANG Zhong-hua, MO Qiu-li, HU Cong, XU Chao-hao, HU Gang
(School of Environmental and Life Sciences,Nanning Normal University,Nanning 530100,China)
Abstract:Sesbania cannabina is a serious alien invasive weed in South China,Central China and East China. This study measured the data such as biomass partitioning in modules (root,stem,leaf,flower and fruit) and plant height,analyzed the characteristics of biomass partitioning in modules and its relationship with allometric growth so as to understand the regulation of growth and development of this alien invasion weed. Results showed that biomass partitioning in modules presented with stem>leaf>fruit>root>flower,and their coefficients of variation were fruit>stem>leaf>root>flower. Meanwhile,there were very significant difference between biomass partitioning in reproductive and nutritional modules or aboveground and underground modules (P<0.01). In addition,all the biomass partitioning in modules were the allometric growth relationships,so were they and plant heightwith highly significant positive correlation (P<0.01). Allometric growth and higher biomass in underground modules of S. cannabina were closely related to its strong invasiveness. The results was of great value to evaluate the invasion mechanism of S. cannabina.
Key words:Sesbania cannabina;alien invasive weed;biomass partitioning in modules;allometric growth
收稿日期:2022-07-31
基金項目:廣西自然科學基金(編號:2021GXNSFFA196005、2021GXNSFAA196024);國家自然科學基金(編號:31960275、31760128)。
作者簡介:鄧美菁(2002—),女,湖南衡陽人,主要從事外來入侵植物方面的研究。E-mail:3320427468@qq.com。
通信作者:胡 剛,博士,副教授,主要從事植被生態學研究。E-mail:ahhugang@126.com。
外來入侵物種會對入侵地的生態系統構成一定的威脅,可通過物種替代的方式改變入侵地生態關鍵過程,這已成為全球生物喪失多樣性的重要因素之一[1]。外來物種入侵已對全球生態、經濟和社會造成諸多負面影響,已成為政府、民眾和學者高度關注的生態環境問題,因而監測和研究入侵物種、防控其傳播擴散顯得尤為重要[2-3]。就外來入侵植物而言,其多為繁殖速度快、適應能力強的物種,侵占入侵地土著植物的生存空間并排擠其土著植物,對入侵地土著植物多樣性造成一定的危害[4]。伴隨我國經濟的快速發展,外來植物入侵問題越來越嚴重,已造成了不可估量的經濟損失[5]。最新研究表明,我國共有歸化植物1 099種,隸屬于112科578屬[6],其中外來入侵植物達72科285屬515種,這些外來入侵植物尤以菊科、豆科和禾本科為主,生活型以草本為主[7]。一些適應能力強的外來入侵植物如紫莖澤蘭(Ageratina adenophora)、加拿大一枝黃花(Solidago canadensis)、喜旱蓮子草(Alternanthera philoxeroides)、互花米草(Spartina alterniflora)等已對我國陸地或水域生態系統造成了嚴重的影響。
田菁(Sesbania cannabina)是豆科(Fabaceae)田菁屬(Sesbania)的一年生亞灌木狀草本植物,株高可達3 m,原產于澳大利亞至西南太平洋島嶼,歸化于亞洲、非洲部分地區以及印度洋島嶼,早期作為綠肥植物被有意引入我國,現已入侵至我國的廣東、廣西、湖南、湖北、上海、浙江等18個省(市),屬于嚴重入侵類的外來植物[7]。田菁適應性強,耐鹽性、耐澇性、耐旱性、抗逆性和抗病蟲害能力也較強,可快速逸生為習見的高大雜草,常在荒地、棄耕地以及人為擾動較大的區域形成優勢種群,排擠土著植物[8]。迄今,針對田菁的葉綠體全基因組測序[9]、種子萌發的生物學特征[8]、營養器官的解剖結構特征[10]、抗逆性[11]等方面都已有了一定的研究。然而,與紫莖澤蘭、加拿大一枝黃花、喜旱蓮子草等外來入侵植物相比,同樣作為一種對我國陸域生態系統造成嚴重入侵的外來物種,除有學者初步總結了其在我國的分布區范圍外[6],幾乎無其入侵機制方面的研究報道,不利于其防控措施的制定。
植物生物量的分配策略是植物對環境適應的結果,與植物的生長發育息息相關,反映出植物在各構件生物量分配中的權衡關系[12-13]。有學者對外來入侵植物進行了構件生物量分配格局與異速生長分析,例如,張橋英等探討了增溫對入侵植物馬纓丹(Lantana camara)生物量分配和異速生長的影響,發現溫度升高有利于其同化作用的增強,從而提高其競爭力[14];閆小紅等分析了不同生育期入侵植物大狼耙草(Bidens frondosa)的生物量分配格局及異速生長,發現其生長特性、生物量分配格局及異速生長關系在不同生育期間存在較大差異,體現了其靈活的生長和資源分配策略[15];Rayamajhi等構建了佛羅里達州外來入侵藤本植物黃獨(Dioscorea bulbifera)的異速生長模型,該模型為估計該入侵植物生物量組成和營養繁殖體密度提供了非破壞性的方法[16]。另有研究表明,異速生長和干擾之間的相互作用可能是喜旱蓮子草成功入侵的部分原因[17]。此外,對銀膠菊(Parthenium hysterophorus)[18-19]、飛機草(Chromolaena odorata)[20]、牛膝菊(Galinsoga parviflora)[21]等入侵植物均進行了構件生物量分配格局或構件異速生長方面的研究。上述學者從構件生物量分配和異速生長視角分析不同外來入侵植物的生長發育規律,有助于深入探討這些物種的入侵機制。然而,對于已在我國中東和華南地區造成嚴重入侵后果的外來雜草田菁仍未開展過相關研究,尚不清楚其構件生物量的分配規律及其與入侵能力間的關系。
廣西南寧的荒地、棄耕地和公共綠地等開闊生境往往分布有較高優勢度的田菁種群,在較大程度上抑制了土著植物的生長,并造成較為明顯的土著生物多樣性喪失。為分析田菁的生長發育規律,本研究通過分析田菁根、莖、葉、花、果的生物量分配情況并比較各構件(包括地上與地下構件、營養與生殖構件)生物量間及其與株高間的異速生長特征,解析田菁種群各構件生物量之間的分配特征和異速生長關系,以期為探究其入侵機制和防控種群擴散提供科學依據。
1 材料與方法
1.1 研究區概況
南寧市(22°13′~23°32′N,107°45′~108°51′E)位于廣西壯族自治區的南部,北回歸線南側,屬濕潤的亞熱帶季風氣候,陽光充足,雨量充沛,霜少無雪,氣候溫和,夏長冬短。年均氣溫約21.6 ℃,年均降水量達1 304.2 mm,平均相對濕度為79%。南寧地區的地貌主要有平地、低山、石山、丘陵和臺地5種類型,地帶性土壤類型為紅壤,地帶性植被為亞熱帶常綠闊葉林。良好的地理和氣候條件孕育了豐富的植物資源,已知有維管束植物3 000余種,隸屬于209科764屬[22]。
1.2 測定方法
2020年8月,選取南寧城市荒地中正處于花果期的田菁種群并對其進行取樣。隨機選取40株大小不等的田菁成熟植株,用鋼卷尺測量其株高,然后對每株田菁地上部分齊地面進行剪割,剪取后用枝剪分離花、果實、葉以及莖稈,將其分別裝入信封,稱取鮮重,再帶回實驗室備測干重;采用挖掘法連同土壤一起挖出所有根系,編號后帶回實驗室,洗凈根上的泥土并稱取鮮重。將田菁各部分構件(根、莖、葉、花、果)放入105 ℃烘箱中殺青15 min,然后75 ℃烘至恒重,稱取干重并記錄各構件的生物量數據。
1.3 數據分析
首先對田菁的各構件生物量進行描述性統計分析,然后對各構件生物量數據進行顯著性檢驗(t檢驗和方差分析)和相關性分析,再對各構件的生物量與株高數據進行擬合分析,最后對不同擬合模型的相關系數作顯著性檢驗。圖表中數據為“平均值±標準差”,采用SPSS 22.0統計分析軟件進行數據分析。
為分析不同構件間的異速生長關系,不同構件生物量用方程y=bxa來描述,線性轉換成ln y=ln b+a(ln x),式中:x和y分別為不同生物量數據;b為構件生物量關系的截距;a為兩者關系的斜率,也就是相對生長的指數,a=1時為等速生長,a≠1時為異速生長。分析方法采用標準化主軸估計(standardized major axis estimation,SMA)法,各參數的確定均在軟件SMATR[23]中完成。
2 結果與分析
2.1 田菁構件間的生物量分配
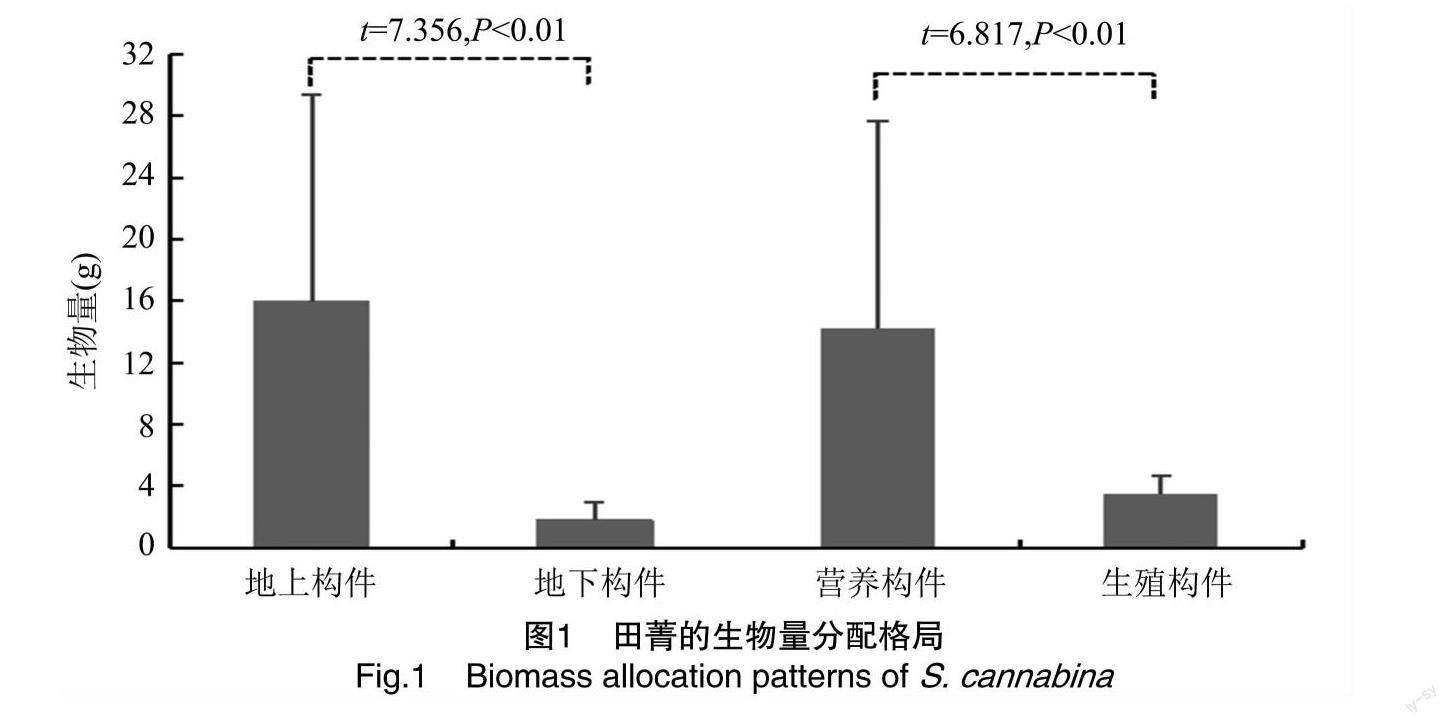
由描述性統計分析結果(表1)可知,田菁種群各構件生物量大小關系為莖>葉>果>根>花,變異系數(coefficient of variation,CV)為51.28%~114.86%,其中根與莖、葉、花之間生物量差異顯著(P<0.05),花與莖、葉、果和根之間差異顯著(P<0.05)。田菁株高為43.30~180.40 cm,CV為39.75%。田箐總生物量為3.54~61.85 g,平均值為17.71 g,CV為81.82%。由圖1可知,田菁種群的生物量分配關系表現為地上構件>地下構件、營養構件>生殖構件,且地上與地下構件、營養與生殖構件間均存在極顯著差異(P<0.01)。
2.2 田菁構件生物量間的異速生長關系
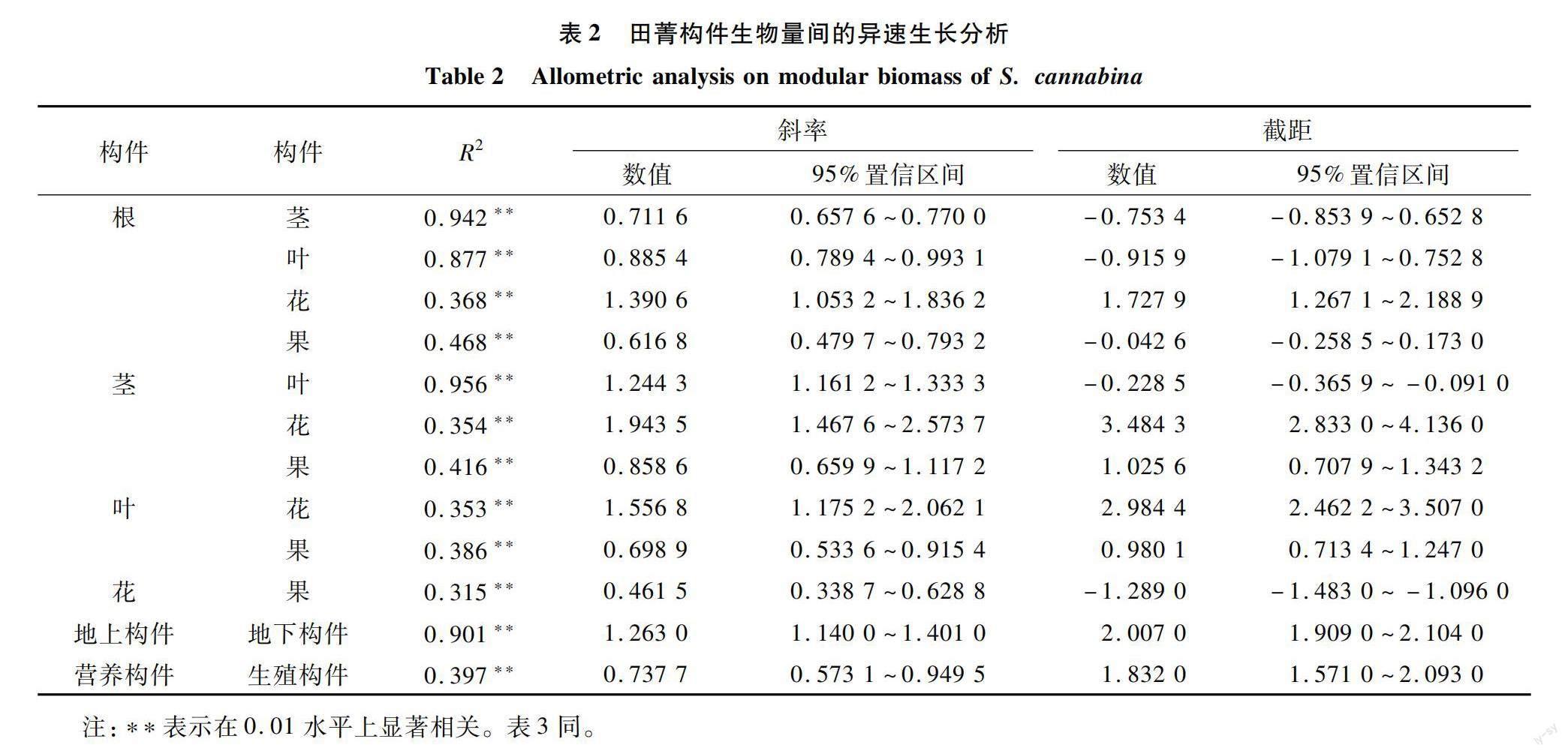
如表2所示,田菁各構件生物量間均為異速生長關系(a≠1)。地上構件與地下構件生物量差異極顯著(P<0. 01),斜率為1.263[95%置信區間(CI)=1.140~1.401],表明地上構件與地下構件之間不成比例地生長。隨著田菁地下構件生物量的增加,地上構件生物量的增加速度變緩。同時,營養構件與生殖構件生物量也呈極顯著關系(P<0.01),斜率為0.737 7(CI=0.573 1~0.949 5);隨著生殖構件生物量的增加,營養構件根、莖、葉的增加速度變緩。地上構件與地下構件、營養構件與生殖構件、根、莖、葉、花、果之間均表現出異速生長關系(a≠1)。
2.3 田菁株高與構件生物量間的擬合與異速生長關系
回歸分析和相關性檢驗(圖2)表明,在田菁繁殖生長期間,營養器官根、莖、葉的生物量和繁殖器官花、果實的生物量都與株高存在極顯著的正相關關系(P<0.01),根、莖、葉、花和果生物量都隨著株高的增長而提高。株高與總生物量、葉生物量和果生物量間可用冪函數較好擬合(圖2-a、圖2-d、圖2-f),株高與根和莖的生物量間可用對數函數擬合(圖2-b、圖2-c),而株高與花生物量間利用線性函數較好擬合(圖2-e)。此外,田菁株高與總生物量、各構件生物量間均表現為異速生長關系(a≠1)(表3)。
3 討論與結論
3.1 田菁的構件生物量結構及其分配特征
生物量是植物的基本生物學特征和功能性狀之一,其在各構件上的分配受遺傳特性和環境因素共同作用,反映了植物的物質積累狀況和對環境資源利用的能力[24]。莖和葉是植物合成、儲存和運輸營養物質的重要構件,所以其生物量投入往往最大[25]。本研究結果表明,田菁種群的莖和葉生物量占比分別為36.82%和32.85%,說明田菁將近70%的生物量分配給能獲取資源的營養構件。Tilman認為,一年生植物通常將更多的資源分配給地上部分,而多年生植物則將更多的資源分配給地下部分[26]。本研究表明,一年生植物田菁具有較高的地上構件生物量和較低的地下構件生物量,進一步印證了上述觀點。田菁較低的根冠比(0.11),表明其較小的地下構件投入便能支持較大的地上構件生長發育。繁殖是生物繁衍后代以及延續種族最基本的行為和過程,也是生物生活史過程中最為重要的一環,植物種群的更新和種群中植物個體生活史的完成均依賴于其繁殖過程[27]。植物在生殖期,其植株需要將部分資源分配給生殖構件,這勢必減少對包括根等其他構件的分配。田菁各構件生物量的變異系數為51.28%~114.86%,與入侵能力強的小飛蓬的生物量變異系數(99.69%~107.48%)[28]相比,其各構件的表型可塑性可能更強。田菁各構件生物量分配比的變異系數大小排列為果>莖>葉>根>花,表明果、莖等構件生物量可塑性可能更強。
最優分配理論(optimal partitioning theory)認為,植物傾向于將生物量分配給能夠獲取稀缺資源的器官以最大化適應外界環境的變化[29],也有研究認為生物量分配是由植物本身的生長模式可塑性主導的[30]。田菁在其入侵的群落中會形成高密度的種群,密度的增大可引起多種資源的變異,涉及復雜的種內相互作用(包括促進作用和競爭作用)。有研究表明,個體大小的增加使植物間相互作用(同時有地上構件和地下構件相互作用)增強,并逐漸開始發生競爭作用,貧瘠的土壤中資源匱乏加劇了植物地下構件間的競爭強度,導致根生物量顯著降低[31]。隨著植株的生長,貧瘠土壤中高密度田菁種群傾向于降低根生物量和根冠比,這可能是其地下構件生物量較低的原因之一。然而,田菁種群密度增加與其生境適應策略關系仍需進一步研究。
3.2 田菁的構件生物量分配與其入侵能力的關系
外來植物的入侵性與植物本身的遺傳機制和對環境的適應能力有關[32-34]。在入侵地天敵的缺乏、資源利用率高、生長競爭力強、繁殖能力強和生態幅廣等因素均決定了外來植物的入侵性。田菁是一年生草本植物,其植株高大(平均株高為93.46 cm),在群落中與低矮的土著植物相比更容易獲取光照,從而影響土著植物的生長發育。田菁葉片生物量占比達32.85%,有利于田菁把更多的光能轉變為化學能,促進田菁的快速生長,而果在總生物量中的占比為17.74%,表明田菁具有較高的種子產量。田菁在長達半年的花果期內可邊開花,邊結果,邊傳播種子,邊萌發形成新個體,這一特性有利于田菁群落搶占更多的生存資源和空間,從而具有更強的繁殖競爭能力。Wang等比較分析了入侵植物與土著植物的性狀差異,表明入侵植物的株高、根冠比和繁殖分配系數等顯著大于土著植物,這些在其成功入侵進程中可能起重要作用[33]。Cheng等研究發現,入侵植物加拿大一枝黃花生長過程中會將更多的生物量分配給繁殖器官,從而有助于其成功入侵[27]。本研究同樣表明,隨著田菁株高的增加,繁殖構件(花和果)生物量出現顯著增加的趨勢,更高的繁殖生物量分配表明其種群更新能力可能更強。
植物各構件生物量與總生物量以及不同構件生物量間的關系通常表現為異速生長關系,這種關系廣泛存在于植物中,能夠揭示植物構件生物量分配生物學特征間的內在規律[29]。異速生長被認為是表型可塑性的特征之一,是影響植物生物量分配的關鍵因素[35-36]。本研究結果表明,在田菁繁殖期間,其各構件生物量、地上構件與地下構件以及營養構件與生殖構件間均呈異速生長關系。因此,田菁在繁殖期其構件生物量的分配較為靈活,能夠更好地適應環境變化,同時又可以為其更高效的繁殖提供保障,增強了其入侵性。對植物構件異速生長關系及其變化進行研究,有助于了解植物構件對環境的適應性響應,植物構件之間多具有顯著的相關性和異速生長關系,體現了構件之間的協同生長策略[13]。不同構件間異速生長關系存在差異,這在馬纓丹[14]、大狼耙草[15]、喜旱蓮子草[17]等外來入侵植物中也得到了普遍驗證。田菁各構件生物量間及其與株高間均表現為顯著的異速生長關系,同樣表明其能較好地調節植物的生長速率,使其具有更強的適應性和入侵能力。
多樣性阻抗假說 (diversity resistance hypothesis) 認為,群落的生物多樣性對抵抗外來種的入侵起著關鍵性作用,越是物種多樣且結構復雜的生態系統,其空余生態位越少而對入侵種的抵抗能力越強[37]。外來植物的入侵能力在很大程度上取決于生境中可利用的非生物資源如光照、水分、養分等[38]。 田菁往往入侵荒地、棄耕地和公共綠地等人為干擾嚴重的開闊生境,筆者建議在這些生境中以鄉土樹種為主進行植物功能群配置,從而實施生態替代措施,利用鄉土植物對光照、土壤養分等資源的競爭優勢來減少空余生態位,從而阻礙陽性植物田菁的生長和種群擴散,這樣既降低了入侵植物的競爭能力,又提高了生境對入侵植物的抵抗力。
本研究對外來入侵植物田菁種群進行生物量分配和異速生長關系分析,結果表明,田菁不同構件間生物量及其分配具有顯著的差異,這些差異來源于構件間或構件與個體大小之間不同的相對生長速率。田菁各構件生物量分配表現為莖>葉>果>根>花,且地上構件生物量顯著大于地下構件,營養構件生物量顯著大于生殖構件。田菁的株高與各構件生物量呈極顯著的正相關關系,各構件之間以及株高與構件間均呈現異速生長關系,表明田菁在生物量分配時具有靈活性,可增強其對環境的適應性,從而提高其入侵能力。本研究從其生物量分配和異速生長關系角度揭示了外來雜草田菁在花果期的生長特征,有助于進一步理解其環境適應策略和入侵機制,并為其防控提供科學指導。然而,目前對田菁入侵機制的理解還較為有限,進一步的研究需要關注田菁原產地和入侵地的表型可塑性差異及其固氮能力與其入侵性的關系。
參考文獻:
[1]Doherty T S,Glen A S,Nimmo D G,et al. Invasive predators and global biodiversity loss[J]. Proceedings of the National Academy of Sciences of the United States of America,2016,113(40):11261-11265.
[2]Kopf R K,Nimmo D G,Humphries P,et al. Confronting the risks of large-scale invasive species control[J]. Nature Ecology & Evolution,2017,1(6):172.
[3]Luque G M,Bellard C,Bertelsmeier C,et al. The 100th of the world's worst invasive alien species[J]. Biological Invasions,2014,16:981-985.
[4]Paini D R,Sheppard A W,Cook D C,et al. Global threat to agriculture from invasive species[J]. Proceedings of the National Academy of Sciences of the United States of America,2016,113(27):7575-7579.
[5]Horvitz N,Wang R,Wan F H,et al. Pervasive human-mediated large-scale invasion:analysis of spread patterns and their underlying mechanisms in 17 of China's worst invasive plants[J]. Journal of Ecology,2017,105(1):85-94.
[6]許光耀,李洪遠,莫訓強,等. 中國歸化植物組成特征及其時空分布格局分析[J]. 植物生態學報,2019,43(7):601-610.
[7]閆小玲,劉全儒,壽海洋,等. 中國外來入侵植物的等級劃分與地理分布格局分析[J]. 生物多樣性,2014,22(5):667-676.
[8]Iqbal N,Manalil S,Chauhan B S,et al. Germination biology of sesbania (Sesbania cannabina):an emerging weed in the Australian cotton agro-environment[J]. Weed Science,2019,67(1):68-76.
[9]Su C,Chen H F,Chang Z Y,et al. The complete chloroplast genome of Sesbania cannabina (Fabaceae) from China[J]. Mitochondrial DNA (Part B:Resources),2020,5(2):1890-1891.
[10]吳 曉,周金鑫,翁 樂,等. 田菁營養器官的解剖結構特征[J]. 亞熱帶植物科學,2018,47(2):128-133.
[11]Srivastava N,Kumar G.Influence of drought stress on cytological behavior of green manure crop Sesbania cannabina Poir.[J]. Cytologia,2014,79(3):325-329.
[12]Poorter H,Nagel O.The role of biomass allocation in the growth response of plants to different levels of light,CO2,nutrients and water:a quantitative review[J]. Functional Plant Biology,2000,27(6):595.
[13]Liu R,Yang X J,Gao R R,et al. Allometry rather than abiotic drivers explains biomass allocation among leaves,stems and roots of? Artemisia across a large environmental gradient in China[J]. Journal of Ecology,2021,109(2):1026-1040.
[14]張橋英,彭少麟. 增溫對入侵植物馬纓丹生物量分配和異速生長的影響[J]. 生態學報,2018,38(18):6670-6676.
[15]閆小紅,何春蘭,周 兵,等. 不同生育期入侵植物大狼杷草的生物量分配格局及異速生長分析[J]. 生態與農村環境學報,2017,33(2):150-158.
[16]Rayamajhi M B,Rohrig E,Tipping P W,et al. Allometric equations for the invasive vine air potato (Dioscorea bulbifera) in its exotic range in Florida[J]. Invasive Plant Science and Management,2020,13(2):76-83.
[17]Jia X,Pan X Y,Li B,et al. Allometric growth,disturbance regime,and dilemmas of controlling invasive plants:a model analysis[J]. Biological Invasions,2009,11:743-752.
[18]蒲高忠,唐賽春,潘玉梅,等. 入侵植物銀膠菊在不同生境下表型可塑性和構件生物量[J]. 廣西植物,2010,30(5):641-646,625.
[19]Rathee S,Ahmad M,Sharma P,et al. Biomass allocation and phenotypic plasticity are key elements of successful invasion of Parthenium hysterophorus at high elevation[J]. Environmental and Experimental Botany,2021,184(1):104392.
[20]Beest M T,Stevens N,Olff H,et al. Plant-soil feedback induces shifts in biomass allocation in the invasive plant Chromolaena odorata[J]. Journal of Ecology,2009,97(6):1281-1290.
[21]齊淑艷,徐文鐸,文 言. 外來入侵植物牛膝菊種群構件生物量結構[J]. 應用生態學報,2006,17(12):2283-2286.
[22]黃良美,黃玉源,黎 樺,等. 南寧市植物群落結構特征與局地小氣候效應關系分析[J]. 廣西植物,2008,28(2):211-217.
[23]Falster D S,Warton D I,Wright I J. SMATR:standardised major axis tests and routines,Version 2.0[DB/OL]. [2022-05-15]. http://www.bio.mq.edu.au/ecology/SMATR/.
[24]陶 冶,張元明. 準噶爾荒漠6種類短命植物生物量分配與異速生長關系[J]. 草業學報,2014,23(2):38-48.
[25]Poorter H,Niklas K J,Reich P B,et al. Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control[J]. New Phytologist,2012,193(1):30-50.
[26]Tilman D. Plant strategies and the dynamics and structure of plant communities[M]. Princeton:Princeton University Press,1988.
[27]Cheng H Y,Wang S,Wei M,et al. Reproductive allocation of Solidago canadensis L. plays a key role in its invasiveness across a gradient of invasion degrees[J]. Population Ecology,2021,63(4):290-301.
[28]潘玉梅,唐賽春,蒲高忠,等. 外來入侵植物小飛蓬種群構件生物量結構特征[J]. 廣西植物,2009,29(4):476-480.
[29]Reich P B,Luo Y J,Bradford J B,et al. Temperature drives global patterns in forest biomass distribution in leaves,stems,and roots[J]. Proceedings of the National Academy of Sciences of the United States of America,2014,111(38):13721-13726.
[30]McCarthy M C,Enquist B J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation[J]. Functional Ecology,2007,21(4):713-720.
[31]Wang S,Li L,Zhou D W. Root morphological responses to population density vary with soil conditions and growth stages:the complexity of density effects[J]. Ecology and Evolution,2021,11(15):10590-10599.
[32]陳 超,滕文軍,武菊英. 細莖針茅的園林應用、生物學和生態學特性及風險評估[J]. 雜草學報,2022,40(1):1-7.
[33]Wang C Y,Zhou J W,Liu J,et al. Differences in functional traits and reproductive allocations between native and invasive plants[J]. Journal of Central South University,2018,25(3):516-525.
[34]胡沁沁,周 穎,華春霞,等. 浙江農林大學校園外來入侵植物及其入侵風險等級評估[J]. 雜草學報,2021,39(2):28-35.
[35]熊韞琦,趙彩云. 表型可塑性與外來植物的成功入侵[J]. 生態學雜志,2020,39(11):3853-3864.
[36]Weiner J. Allocation,plasticity and allometry in plants[J]. Perspectives in Plant Ecology,Evolution and Systematics,2004,6(4):207-215.
[37]Elton C S. The ecology of invasions by animals and plants[M]. Berlin:Springer Science+Business Media,1958.
[38]廖慧璇,周 婷,陳寶明,等. 外來入侵植物的生態控制[J]. 中山大學學報(自然科學版),2021,60(4):1-11.
