杧果MiOFP1 基因的表達與功能分析
2023-07-27 11:09:44張藝粒朱嘉偉勞安李雨澤夏黎明莫嘯胡萬里何新華羅聰
果樹學報 2023年6期
關鍵詞:功能分析
張藝粒 朱嘉偉 勞安 李雨澤 夏黎明 莫嘯 胡萬里 何新華 羅聰
摘要:【目的】卵形家族蛋白(ovate family proteins,OFPs)在植物生長發育及逆境響應過程中扮演重要角色。前期通過杧果成花基因酵母文庫篩選,獲得了一個MiOFP1 基因,為明確其功能,對MiOFP1 的表達模式和轉基因功能開展了研究。【方法】在本研究中分析了杧果MiOFP1 的啟動子序列;通過實時熒光定量PCR技術分析MiOFP1 在杧果不同組織器官和不同生長發育期葉片中的表達模式;轉化構建好的超量表達載體并侵染擬南芥研究MiOFP1 的功能。【結果】四季蜜杧MiOFP1 啟動子包含激素響應元件:ABA響應元件、GA響應元件、SA響應元件和乙烯響應元件,逆境響應元件:鹽響應元件、脫水響應元件、MYC轉錄因子和MYB轉錄因子結合位點。組織特異性表達分析顯示,MiOFP1在各組織器官中均有表達,且在童期實生樹和成年期嫁接樹的莖中表達量最高,在成熟果實中表達量最低;嫁接樹不同成花發育時期表達分析結果顯示,MiOFP1 在營養生長期的葉中表達量最高,在成花誘導期和花發育期表達水平較低。轉基因功能研究顯示,超量表達MiOFP1 的擬南芥出現晚花表型,抽薹期葉片中成花抑制基因FLOWERINGLOUS C(FLC)的表達水平顯著上調,而成花促進基因FLOWERING LOCUS T(FT)的表達水平顯著下調。逆境脅迫處理顯示,ABA處理顯著抑制擬南芥種子的萌發與根的伸長,但通過轉基因顯著提高了擬南芥種子的萌發率,降低了擬南芥根長對ABA的敏感性。進一步分析顯示,MiOFP1 顯著提高了擬南芥在ABA處理后的脯氨酸含量和過氧化物酶活性,上調了ABA代謝相關基因的表達水平。【結論】明確了杧果MiOFP1 抑制成花,且降低了轉基因植株對ABA的敏感性,為進一步探索杧果MiOFP1 參與杧果成花和逆境脅迫應答的分子機制奠定基礎。
關鍵詞:杧果;MiOFP1;表達模式;功能分析;ABA處理
中圖分類號:S667.7 文獻標志碼:A 文章編號:1009-9980(2023)06-1099-10
卵形家族蛋白(ovate family proteins,OFPs)是具有保守OVATE 結構域的轉錄因子,為植物所特有,因番茄果實形態由卵形變為梨形而發現[1]。研究發現,OFPs 參與植物次生細胞壁形成,水稻維管束發育,擬南芥胚珠發育,辣椒和番茄果實的形狀、香蕉果實的成熟及品質形成等過程[2];在擬南芥中過表達AtOFP1 發現花粉活力受到影響,植物發育遲緩[3];此外,OFPs 參與植物逆境抵御和赤霉素(gibberellin,GA)、脫落酸(abscisic acid,ABA)、乙烯(ethylene)及油菜素內酯(brassinolide,BR)等激素的信號傳遞[4];OsOFP22 通過抑制GA和BR信號傳導,調節水稻形態發育[5];外源施加GA3 抑制SlOFP20 的表達,低溫、干旱、鹽脅迫和外源施加吲哚-3-乙酸(indole-3-acetic acid,IAA)和ABA能促進SlOFP20 的表達[6]。ABA是一種在植物生長發育和各種非生物脅迫耐受過程中起重要調節作用的激素,參與調節種子成熟、種子發芽、幼苗生長和氣孔運動等[7],外源施加ABA可以有效緩解植物受到的冷害、鹽脅迫和干旱脅迫等[8-10]。
杧果(Mangifera indica L.)是漆樹科杧果屬的熱帶水果,是世界五大熱帶水果之一,適時開花和對非生物脅迫的抵御能力影響杧果的產量和品質。OFPs 參與植物的成花調控且與逆境脅迫應答有關[11],目前杧果尚無OFP 基因的研究報道,在前期研究中,筆者從杧果成花酵母文庫中篩選獲得了一個OFP 基因,命名為MiOFP1。筆者在本研究中對杧果MiOFP1 基因的表達模式和轉基因功能進行研究,為揭示MiOFP1 參與杧果成花和杧果逆境應答的分子機制提供參考。
1 材料和方法
1.1 植物材料及取樣
供試杧果品種為四季蜜杧,栽培于廣西大學農學院果樹標本園,擬南芥(Arabidopsis thaliana)為哥倫比亞野生型,種子由廣西大學農學院杧果課題組保存。組織表達特性樣品采集:2022-02-15 采集6株長勢一致的童期實生杧果樹(3 年生)冬梢的成熟葉、芽和莖(韌皮部組織),6 株長勢一致的成年期嫁接杧果樹(12 年生)冬梢的成熟葉、莖(韌皮部組織)、花,以及盛花后20 d 和100 d 的植物果實胚和果肉。時間表達特性樣品為6 株長勢一致的嫁接杧果樹的成熟葉,樣品采集時間為2021 年9 月至2022 年5 月。采樣時間統一定為17:00—18:00,樣品采集后立即處理,放入-80 ℃冰箱冷凍保存備用。
1.2 試驗方法
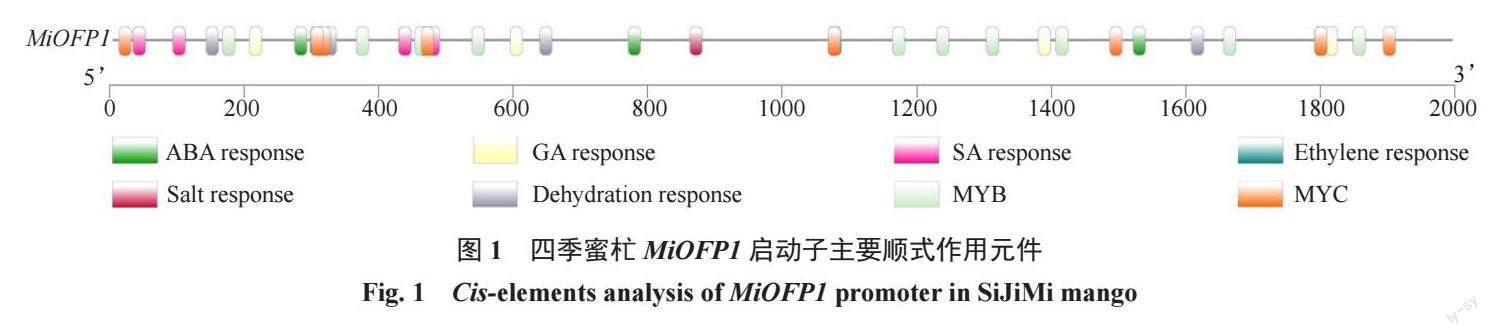
1.2.1 MiOFP1 啟動子序列分析利用NEWPLACE(https://www.dna.affrc.go.jp/PLACE/?action=newplace)在線軟件預測四季蜜杧MiOFP1 啟動子區域順式作用元件,利用TBtools 繪制啟動子順式作用元件位置信息圖。
1.2.2 MiOFP1 組織特性和時間表達特性分析提取杧果組織表達特性樣品和時間表達特性樣品的RNA,逆轉錄為cDNA待用,根據四季蜜杧MiOFP1基因序列設計熒光定量PCR引物qOFP1 F(5CTGTACCTGCCGTGCTACAA3)、qOFP1 R(5CTGTACCTGCCGTGCTACAA3)。以杧果MiActin1 為內參基因,引物為qActin F(5CCGAGACATGAAGGAGAAGC3)、qActin R(5GTGGTCTCATGGATACGAGCA3)。實時熒光定量PCR 儀器為ABI7500。擴增反應體系和程序參照試劑盒SYBR Premix Dimer Eraser (TaKaRa)說明書進行[12]。
1.2.3 擬南芥轉化與成花表型分析實驗室前期已構建好四季蜜杧MiOFP1 的超量表達載體,命名為pBI121-MiOFP1 載體,轉化EHA105 感受態。通過花序侵染法轉化哥倫比亞野生型擬南芥[13]。利用抗生素篩選和PCR技術鑒定陽性植株。以T3 代純合植株為試驗材料進行相關試驗。成花表型觀察:利用半定量技術鑒定杧果MiOFP1 在轉基因擬南芥中的表達水平,并觀察轉基因擬南芥的抽薹時間和第一朵花開放的時間。通過熒光定量技術,采集抽薹期擬南芥葉片,檢測內源成花基因的表達水平,以擬南芥AtActin2 為內參基因,引物為AtActin2 F(5-GCAGAGCGGGAAATTGTAAG- 3)、AtActin2 R(5-GGATATCAGGAAGGATCTGTAC-3),AtFLC F(5-ATCATCATGTGGGAGCAGAAG-3)、AtFLCR(5- TTCAACCGCCGATTTAAGG- 3),AtFT F(5-CTTGGCAGGCAAACAGTGTATGCAC- 3)、AtFT R(5- GCCACTCTCCCTCTGACAATTGTAGA-3),檢測方法參考課題組研究報道[14]。
1.2.4 轉基因擬南芥響應植物激素ABA試驗萌發率測定:將轉基因擬南芥T3 代純合株系種子播種于含0、2、5 μmol·L-1ABA的1/2 MS固體培養基上,每個處理3 次重復,以胚根伸出為萌發依據,每12 h記錄一次萌發率,10 d 后拍照記錄。根長測定:將T3 代純合株系種子播種于1/2 MS固體培養基上培養,5 d 后移至含0、5、10 μmol·L-1ABA的1/2 MS固體培養基上繼續培養,每個處理3 次重復,移栽5 d后拍照并統計根長。生理指標測定:播種9 d 后移栽幼苗到穴盆中,培養12 d 后進行20 μmol·L-1ABA噴施處理,以清水處理為對照,處理1 d 后采集擬南芥葉片,每個處理3 次重復,采用試劑盒檢測脯氨酸含量和過氧化物酶活性(北京索萊寶科技有限公司);提取ABA 處理組及對照組擬南芥葉片的RNA,逆轉錄后通過熒光定量技術檢測參與擬南芥ABA響應的內源基因表達量,引物為AtABI1 F(5-GTTTTCCCGTCTCACATCTTCGT- 3)、AtABI1 R(5- CTTCATCCGTCA TTACATCCCAA- 3),At-MFT F(5-CGAGCCGAACATGAGAGAAT-3)、At-MFT R(5- AAGTATCTCTTTTCCTCTTGAGGG-3)、AtDREB2B F(5- CATCAGAGCCAAGACCAAAACC-3)、AtDREB2B R(5- TGTAGGACCATTGCCTCAGAAC-3)[15]。
1.3 數據分析
將所得數據采用Excel 進行數據處理及圖片制作,利用IBM SPSS 22.0 軟件進行方差分析。
2 結果與分析
2.1 MiOFP1基因啟動子序列分析
對四季蜜杧MiOFP1 的啟動子序列(2000 bp)進行順式作用元件預測,發現MiOFP1啟動子中存在的激素響應元件有:1個乙烯響應元件,5個ABA響應元件,7個SA響應元件,9個GA響應元件;逆境響應元件有:1個鹽響應元件,4個脫水響應元件,9個MYC轉錄因子和10個MYB轉錄因子結合位點(圖1)。
2.2 MiOFP1 表達模式分析
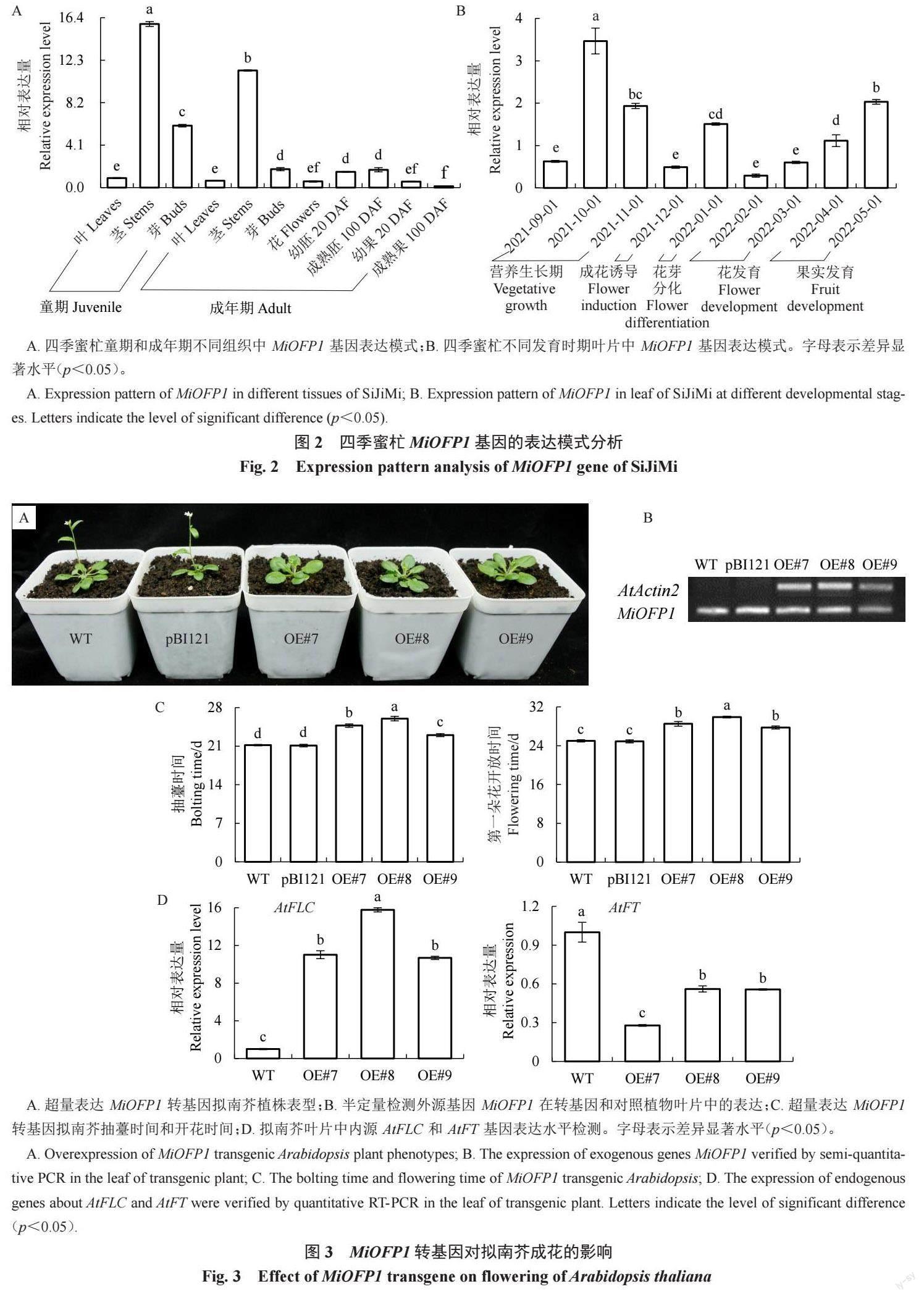
對MiOFP1 在杧果不同生長發育周期的不同組織器官中的表達模式進行實時熒光定量分析。結果如圖2- A 所示,MiOFP1 存在組織表達差異性。MiOFP1在童期樹組織中的表達水平高于成年期樹。MiOFP1在莖的韌皮部中表達水平較高,在葉、花和幼果中表達水平最低,在成熟果實中表達水平最低。不同成花發育時期葉片中的表達模式分析顯示,MiOFP1在營養生長期的葉片中表達水平較高,在成花誘導期和花發育期的葉片中表達水平較低,而后在果實發育期的葉片中表達水平逐步上升(圖2-B)。
2.3 擬南芥轉化及成花表型分析
將包含pBI121-MiOFP1 超量表達載體和空載體的農桿菌通過花序侵染法分別轉化模式植物擬南芥。半定量PCR 檢測結果顯示,轉基因株系MiOFP1#7、MiOFP1#8 和MiOFP1#9 中MiOFP1 正常表達,而在對照植株中沒有表達(圖3-B)。超量表達MiOFP1 轉基因株系表現出晚花表型,其抽薹時間和第一朵花開放時間均比WT和轉空載體植株(pBI121)晚2~4 d(圖3-A、C)。轉基因擬南芥內源成花基因表達水平檢測顯示,超量表達MiOFP1 顯著提高了擬南芥AtFLC 的表達水平,顯著下調了At-FT的表達水平(圖3-D)。
2.4 外源ABA處理對MiOFP1 轉基因擬南芥的影響
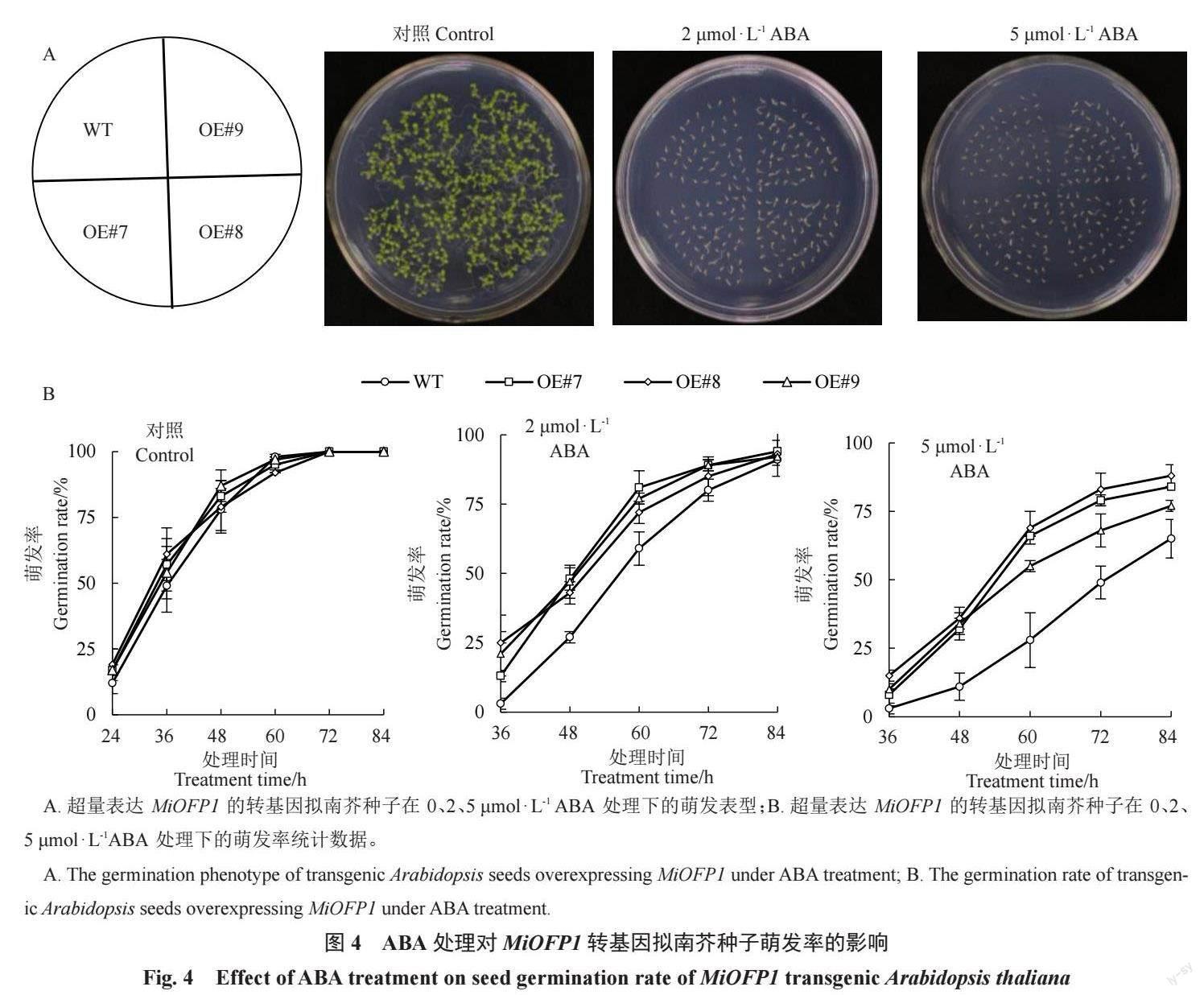
2.4.1 外源ABA 處理后MiOFP1 轉基因擬南芥種子萌發表型分析T3 代純合擬南芥種子在0、2、5 μmol · L-1 ABA的1/2 MS固體培養基上培養,記錄發芽情況。ABA 處理10 d 后拍照記錄如圖4-A所示,在未添加ABA的培養基上,擬南芥幼苗長勢基本一致;在添加ABA的培養基中,所有種子發芽均受抑制,種子在含5 μmol·L-1ABA的1/2 MS固體培養基上的長勢較在含2 μmol·L-1ABA的1/2 MS固體培養基上更弱。萌發率統計分析顯示(圖4-B),未添加ABA時,前60 h 轉基因擬南芥種子的萌發率略高于WT種子,但差異不顯著;在2 μmol·L-1 ABA 處理時,所有種子萌發均受到抑制,轉基因擬南芥種子萌發速度明顯快于WT種子;在5 μmol·L-1 ABA處理時,種子萌發受到的抑制更強,轉基因擬南芥種子的萌發率極顯著高于對照植株,最終的轉基因擬南芥種子發芽率分別為84%、88%、77%,WT種子發芽率為65%。
2.4.2 外源ABA 處理對超量表達MiOFP1 擬南芥根長、生理指標與內源基因表達的影響不同濃度的ABA處理對T3 代純合擬南芥幼苗根長的影響如圖5-A,在未施加ABA時,轉基因擬南芥與對照擬南芥的根長無顯著差異,隨著ABA濃度的增加,轉基因擬南芥與對照擬南芥幼苗的根長均表現為受抑制,但在相同處理下,對照植株根長顯著短于轉基因植株的根長(圖5-B)。
對轉基因植株與對照WT植株進行20 μmol·L-1ABA噴施處理,以清水噴施處理為對照,處理后1 d采集葉片為樣品,對處理組與對照組植株進行生理指標測定和擬南芥內源基因表達水平檢測。如圖5-C所示,清水對照處理下,轉基因植株與WT植株的脯氨酸含量和過氧化物酶活性無明顯差異,但在ABA處理后,轉基因植株及WT植株的脯氨酸含量均有增加,且轉基因植株脯氨酸含量顯著高于WT植株,是WT植株的1.4 倍;在ABA處理后,轉基因植株的過氧化物酶活性顯著高于對照WT植株。
內源基因檢測顯示,清水處理時,MiOFP1 轉基因株系中AtABI1、AtRD29B和AtNCED3 的表達水平略低于WT株系,而AtMFT 的表達水平略高于WT植株。在ABA處理后,上述4 個基因的表達水平均顯著高于WT植株(圖5-D)。
3 討論
啟動子順式作用元件關系著植物對激素、低溫、鹽和干旱等非生物脅迫作出反應,非生物脅迫與激素均可誘導相關轉錄因子結合到下游基因啟動子順式作用元件上,從而誘導其表達[16- 17],MiOFP1 前2000 bp 啟動子中包含大量激素響應元件和逆境脅迫響應元件,推測MiOFP1 可能受到激素、非生物脅迫以及上游MYB、MYC 轉錄因子的調控和誘導。組織表達分析表明,MiOFP1 具有組織表達特異性,主要在童期樹和成年期樹的莖中表達,在花、葉、果實和胚中表達量相對較低。此前的研究發現,水稻OsOFP在種子發育時期表達量較高[18],葡萄VvOFP在開花前表達量較高,開花一段時間后,表達量顯著下降[19],小麥TaOFP 在根和穗中表達量最高[20]。對比本研究結果,可以推測,OFP 參與多個植物生長發育的過程,但是不同物種OFP主要發揮的功能和表達的位置可能不同。此外,對過表達MiOFP1 擬南芥進行表型觀察,發現抽薹時間和開花時間有顯著延遲,這與擬南芥OFP1 功能類似,AtOFP1 在擬南芥中過表達出現延遲開花現象[21]。內源基因表達分析結果表明,在過量表達MiOFP1 的擬南芥株系中,成花抑制基因FLC 的表達得到促進,成花促進基因FT 的表達受到抑制,從而使植物開花時間延遲,這證明了MiOFP1 對轉基因擬南芥的開花起到抑制作用。研究發現,過量表達SlOFP20 的番茄開花時間明顯推遲,SlOFP20 轉基因植株的SlFT 表達水平顯著低于WT[22]。目前尚未見有OFP在杧果中是否同樣具有延遲開花的功能報道,但過量表達MiOFP1 的擬南芥植株與過量表達番茄SlOFP20 的植株均出現延遲開花現象,且植株內FT的表達量均顯著降低,表明二者可能通過相同或相似的途徑調控FT 的表達,從而實現對植株開花時間的調控,綜上所述,推測MiOFP1 通過影響FLC和FT的表達水平來調節杧果開花時間。
已有研究表明,除參與植物生長發育外,OVATE 家族的多數基因還參與植物激素調節及逆境脅迫響應過程。外源施加GA3可以減緩AtOFP1超表達株系的矮化現象;水稻OFP1 通過與GSK2、OsBZR1 和DLT 的相互作用來正調控BR 響應;Os-OFP6-RNAi 轉基因株系表現出正常生長條件下側根變短和IAA 處理后側根密度增加[22];過表達AtOFP8 顯著增強了擬南芥的抗旱性,其種子的發芽率和綠葉率更高,葉片中脯氨酸含量提高,丙二醇含量降低,可溶性蛋白含量高,葉片抗氧化酶的活性較強[23];對去除水稻OsOFP6 的植株進行干旱和低溫處理,明顯發現植株缺水且相對電導率升高的現象,說明OsOFP6 參與水稻對干旱和低溫脅迫的防御過程[24];蘋果OFP 家族基因在鹽、干旱和高低溫等非生物逆境下呈現出不同的響應差異,尤其是在低溫下MdOFP基因家族中多個基因的表達量均有不同程度的上升[25];同樣的,低溫條件下,油桐幼苗VfOFP3/7/10/12 等基因的表達量上升,推測VfOFP 可能在油桐低溫脅迫響應方面發揮積極的調節作用[26]。ABA 是種子萌發和大多數常見非生物脅迫反應的主要介質之一,包括對鹽、干旱和寒冷的反應[27-28]。與野生型植物相比,MiOFP1 的超量表達減輕了ABA對種子萌發的抑制作用和根長敏感性,提高了外源ABA 情況下植株的脯氨酸含量和過氧化物酶活性,促進了ABA 響應基因ABI1、MFT 和RD29B 以及ABA 合成基因NCED3 的表達。植物在受到脅迫時,細胞的滲透壓發生變化,脯氨酸含量會升高以維持細胞滲透壓;同時,細胞內產生大量的過氧化氫及酚類、胺類等物質,對細胞造成損害,過氧化物酶可催化過氧化氫及酚類、胺類等物質,達到減輕上述物質對細胞的毒害作用,通過外源噴施ABA,提高植物體內的脯氨酸含量和過氧化物酶活性,從而增強植物抵御非生物脅迫的能力[29-30];ABI1 在ABA的信號轉導過程中起負調控作用[31],MFT直接抑制ABI5 的表達,對ABA信號通路起負反饋調節作用[32],外源ABA 處理明顯誘導RD29B 表達量升高[33]。這些發現表明,MiOFP1 在杧果開花和外源ABA 響應中可能具有關鍵的作用,推測杧果MiOFP1 可能是通過介導ABA的信號通路參與種子萌發調控過程和逆境防御過程,但是MiOFP1 是如何參與ABA 信號通路的、MiOFP1 是否影響杧果開花、能否通過提高ABA 含量來增強杧果逆境抵御能力還不清楚,需要進一步研究。
4 結論
杧果MiOFP1 在童期莖的韌皮部組織中高度表達,而在成熟果實的果肉中表達水平較低;在營養生長期葉片中表達水平較高,而在成花期表達水平較低。超量表達的MiOFP1 抑制擬南芥開花,但提高了種子在ABA 處理下的萌發率,減弱了擬南芥對ABA 脅迫的敏感性。說明MiOFP1 可能與杧果的成花和ABA脅迫有關。
參考文獻References:
[1] LIU J P,VAN ECK J,CONG B,TANKSLEY S D. 番茄OVATE基因調控果實梨形發育[J]. 楊莉,張嵐嵐,徐昌杰,譯. 植物學通報,2003,38(1):127.LIU J P,VAN Eck J,CONG B,TANKSLEY S D. OVATE generegulates pear shape development of tomato fruit[J]. ,YANG Li,ZHANG Lanlan,XU Changjie,Translated. Chinese Bulletin ofBotany,2003,38(1):127.
[2] 張靜,謝碧玉,徐碧玉,劉菊華. 卵形家族蛋白對植物生長發育的調控作用[J]. 分子植物育種,2019,17(10):3283-3288.
ZHANG Jing,XIE Biyu,XU Biyu,LIU Juhua. Regulatory rolesof ovate family proteins on plant growth and development[J].Molecular Plant Breeding,2019,17(10):3283-3288.
[3] HACKBUSCH J,RICHTER K,M?LLER J,SALAMINI F,UHRIG J F. A central role of Arabidopsis thaliana ovate familyproteins in networking and subcellular localization of 3-aa loopextension homeodomain proteins[J]. Proceedings of the NationalAcademy of Sciences of the United States of America,2005,102(13):4908-4912.
[4] 李曉. 水稻OsOFP22 轉錄因子功能分析[D]. 長春:吉林大學,2017.
LI Xiao. The functional analysis of OsOFP22 transcription factor[D]. Changchun:Jilin University,2017.
[5] 陳浩源. OsOFP22 通過赤霉素和油菜素甾醇調控水稻株型和粒型的機制分析[D]. 長春:吉林大學,2021.
CHEN Haoyuan. Mechanism analysis of OsOFP22 regulatingplant morphology and grain shape by gibberellin and brassinosteroidin rice (Oryza sativa L.)[D]. Changchun:Jilin University,2021.
[6] ZHOU S G,CHENG X,LI F F,FENG P P,HU G L,CHEN GP,XIE Q L,HU Z L. Overexpression of SlOFP20 in tomato affectsplant growth,chlorophyll accumulation,and leaf senescence[J]. Frontiers in Plant Science,2019,10:1510.
[7] SHAN Y J,LI D,CAO J J,ZHANG L,HAN L Q,ZHANG MP,SHEN Z G. Over- expression of Arabidopsis ORANGE geneenhances drought stress tolerance through ABA-dependent pathwayin Arabidopsis thaliana[J]. Plant Growth Regulation,2022,96(1):91-101.
[8] 梁瀟,劉丹梅,姜兆彤,刁玉峰,劉文璐,袁珺.‘紅顏草莓栽培種FaNCED4 基因克隆與脅迫表達[J/OL]. 分子植物育種,2022:1-10[2022-07-10]. http://kns.cnki.net/kcms/detail/46.1068.S.20220613.1150.010.html.
LIANG Xiao,LIU Danmei,JIANG Zhaotong,DIAO Yufeng,LIU Wenlu,YUAN Jun. Cloning and expression of FaNCED4gene in‘Benihoppestrawberry cultivars under stress[J/OL].Molecular Plant Breeding,2022:1- 10[2022- 07- 10]. http://kns.cnki.net/kcms/detail/46.1068.S.20220613.1150.010.html.
[9] 羅忍忍,王瑞丹,曹磊,李麗麗,李翔,袁燁,晏家茹,侯娟,胡建斌. 植物生長調節劑對冷脅迫下甜瓜幼苗生理特性及相關基因表達的影響[J]. 河南農業大學學報,2022,56(3):411-419.
LUO Renren,WANG Ruidan,CAO Lei,LI Lili,LI Xiang,YUANYe,YAN Jiaru,HOU Juan,HU Jianbin. Effects of plantgrowth regulators on physiological characteristics and relatedgene expression in melon seedlings under cold stress[J]. Journalof Henan Agricultural University,2022,56(3):411-419.
[10] 李振華,劉容,張馨馨,趙心笛,劉敏婷,柴琦. 外源脫落酸增強高羊茅耐鹽性的作用[J]. 北方園藝,2022(7):66-75.
LI Zhenhua,LIU Rong,ZHANG Xinxin,ZHAO Xindi,LIUMinting,CHAI Qi. Effects of exogenous abscisic acid on enhancingsalt tolerance of Festuca arundinacea[J]. Northern Horticulture,2022(7):66-75.
[11] 盧琳. OsOFP19 過表達轉基因水稻農藝性狀與干旱響應的鑒定與分析[D]. 長春:吉林大學,2022.
LU Lin. Identification and analysis of agronomic traits anddrought response of OsOFP19-overexpressing transgenic rice[D].Changchun:Jilin University,2022.
[12] 范志毅,羅聰,余海霞,曾學梅,王金英,謝小杰,何新華. 芒果MiFY 基因克隆和表達模式分析[J]. 熱帶作物學報,2021,42(2):297-302.
FAN Zhiyi,LUO Cong,YU Haixia,ZENG Xuemei,WANGJinying,XIE Xiaojie,HE Xinhua. Cloning and expression analysisof a MiFY gene in mango[J]. Chinese Journal of TropicalCrops,2021,42(2):297-302.
[13] GUO Y H,LUO C,LIU Y,LIANG R Z,YU H X,LU X X,MOX,CHEN S Q,HE X H. Isolation and functional analysis of twoCONSTANS- like 1 genes from mango[J]. Plant Physiology andBiochemistry,2022,172:125-135.
[14] MO X,LUO C,YU H X,CHEN JW,LIU Y,__________XIE X J,FAN Z Y,HE X H. Isolation and functional characterization of two SHORTVEGETATIVE PHASE homologous genes from mango[J]. InternationalJournal of Molecular Sciences,2021,22(18):9802.
[15] ARIF M,LI Z T,LUO Q,LI L H,SHEN Y Q,MEN S Z. TheBAG2 and BAG6 genes are involved in multiple abiotic stresstolerances in Arabidopsis thaliana[J]. International Journal ofMolecular Sciences,2021,22(11):5856.
[16] 楊天宸,陳曉童,呂可,張荻. 百子蓮脫水素基因ApSK3 對逆境與激素信號的應答模式與調控機制[J]. 園藝學報,2021,48(8):1565-1578.
YANG Tianchen,CHEN Xiaotong,L? Ke,ZHANG Di. Expressionpattern and regulation mechanism of ApSK3 dehydrin (Agapanthus praecox) response to abiotic stress and hormonesignals[J]. Acta Horticulturae Sinica,2021,48(8):1565-1578.
[17] 李世琦. 巨球百合細胞壁轉化酶基因及其啟動子功能初探[D].杭州:浙江大學,2021.
LI Shiqi. A preliminary study on the function research of cellwall invertase gene and its promoter of Lilium brownii var. giganteum[D]. Hangzhou:Zhejiang University,2021.
[18] 于慧. 水稻OsOFP 轉錄因子家族基因克隆與功能分析[D]. 長春:吉林大學,2015.
YU Hui. The genes cloning and functional analysis of OsOFPtranscription factor family in rice (Oryza sativa L.)[D]. Changchun:Jilin University,2015.
[19] 袁月,張亞光,高世敏,陶建敏. 葡萄OVATE 基因家族生物信息學及表達[J]. 中國農業科學,2016,49(19):3786-3797.
YUAN Yue,ZHANG Yaguang,GAO Shimin,TAO Jianmin.Bioinformatics and expression of the OVATE gene family ingrape[J]. Scientia Agricultura Sinica,2016,49(19):3786-3797.
[20] 徐渴,王偉偉,武寧靜,張樹華,趙勇,楊學舉. 小麥OFP 基因家族鑒定及其低溫脅迫的表達分析[J]. 農業生物技術學報,2021,29(9):1665-1677.
XU Ke,WANG Weiwei,WU Ningjing,ZHANG Shuhua,ZHAO Yong,YANG Xueju. Identification of OFP gene familyand their expression analysis under low- temperature stress inwheat (Triticum aestivum) [J]. Journal of Agricultural Biotechnology,2021,29(9):1665-1677.
[21] 蔡玲. 卵形蛋白家族轉錄因子AtOFP15 對擬南芥莢果形態的調控[D]. 長春:東北師范大學,2017.
CAI Ling. Regulation of silique morphology by ovate familyprotein AtOFP15 in Arabidopsis[D]. Changchun:Northeast NormalUniversity,2017.
[22] 周升恩. 兩個番茄轉錄因子SlOFP20 和SlGT16 在生長發育中的功能研究[D]. 重慶:重慶大學,2020.
ZHOU Shengen. Functional study of two tomato transcriptionfactors SlOFP20 and SlGT16 genes in growth and development[D]. Chongqing:Chongqing University,2020.
[23] 唐堯. 擬南芥轉錄因子AtOFP8 在干旱脅迫下的功能分析[D].哈爾濱:東北農業大學,2018.
TANG Yao. Functional analysis of Arabidopsis transcriptionalfactor AtOFP8 under drought stress[D]. Harbin:Northeast AgriculturalUniversity,2018.
[24] MA Y M,YANG C,HE Y,TIAN Z H,LI J X. Rice OVATEfamily protein 6 regulates plant development and confers resistanceto drought and cold stresses[J]. Journal of ExperimentalBotany,2017,68(17):4885-4898.
[25] 許瑞瑞,李睿,王小非,郝玉金. 蘋果OFP 基因家族的全基因組鑒定與非生物逆境表達分析[J]. 中國農業科學,2018,51(10):1948-1959.
XU Ruirui,LI Rui,WANG Xiaofei,HAO Yujin. Identificationand expression analysis under abiotic stresses of OFP gene familyin apple[J]. Scientia Agricultura Sinica,2018,51(10):1948-1959.
[26] 王靜雅,劉文娟,呂祎馨,尚海,米小琴,張琳,劉美蘭. 油桐OFP 基因家族的全基因組鑒定及低溫響應分析[J]. 植物生理學報,2020,56(5):949-960.
WANG Jingya,LIU Wenjuan,L? Yixin,SHANG Hai,MI Xiaoqin,
ZHANG Lin,LIU Meilan. Genome-wide identification andlow temperature response analysis of OFP gene family in tungtree (Vernicia fordii) [J]. Plant Physiology Journal,2020,56(5):949-960.
[27] FUJII H,VERSLUES P E,ZHU J K. Arabidopsis decuple mutantreveals the importance of SnRK2 kinases in osmotic stressresponses in vivo[J]. Proceedings of the National Academy ofSciences of the United States of America,2011,108(4):1717-1722.
[28] NISHIMURA N,YOSHIDA T,KITAHATA N,ASAMI T,SHINOZAKIK,HIRAYAMA T. ABA- Hypersensitive Germination1encodes a protein phosphatase 2C,an essential componentof abscisic acid signaling in Arabidopsis seed[J]. The Plant Journal,2007,50(6):935-949.
[29] 符楊磊,魏志園,王宇,劉瀟陽,王冰冰,喬亞科,李桂蘭,張鍇.冀東野生大豆(Glycine soja)耐鹽堿性鑒定及耐性生理指標測定[J]. 核農學報,2020,34(10):2316-2325.
FU Yanglei,WEI Zhiyuan,WANG Yu,LIU Xiaoyang,WANGBingbing,QIAO Yake,LI Guilan,ZHANG Kai. Identificationof saline- alkali stress and determination of physiological indexof wild soybean (Glycine soja) in eastern Hebei Province[J].Journal of Nuclear Agricultural Sciences,2020,34(10):2316-2325.
[30] 湯日圣,唐現洪,鐘雨,張大棟,余永柱,童紅玉. 微生物源ABA 對茄苗抗冷性和某些生理指標的影響[J]. 園藝學報,2006,33(1):149-151.
TANG Risheng,TANG Xianhong,ZHONG Yu,ZHANG Dadong,YU Yongzhu,TONG Hongyu. Effect of microorganismsourcedABA on chilling resistance and some physiological indexesin eggplant seedling[J]. Acta Horticulturae Sinica,2006,33(1):149-151.
[31] SEO Y J,PARK J B,CHO Y J,JUNG C,SEO H S,PARK S K,NAHM B H,SONG J T. Overexpression of the ethylene-responsivefactor gene BrERF4 from Brassica rapa increases toleranceto salt and drought in Arabidopsis plants[J]. Molecules andCells,2010,30(3):271-277.
[32] 韓萍萍. 藜麥PEBP 基因家族及種子休眠相關基因功能分析[D]. 濟南:山東師范大學,2021.
HAN Pingping. Function analysis of quinoa PEBP family geneand seed dormancy related genes[D]. Jinan:Shandong NormalUniversity,2021.
[33] MSANNE J,LIN J S,STONE J M,AWADA T. Characterizationof abiotic stress- responsive Arabidopsis thaliana RD29A andRD29B genes and evaluation of transgenes[J]. Planta,2011,234(1):97-107.
猜你喜歡
現代交際(2016年20期)2017-02-15 18:33:15
科技資訊(2016年19期)2016-11-15 10:39:12
電子技術與軟件工程(2016年18期)2016-11-14 00:49:04
戲劇之家(2016年20期)2016-11-09 23:08:07
老區建設(2016年4期)2016-09-28 05:09:17
電腦知識與技術(2016年16期)2016-07-22 19:10:51
科技視界(2016年13期)2016-06-13 21:00:36
戲劇之家(2016年6期)2016-04-16 11:52:45
科技視界(2016年4期)2016-02-22 20:53:20
科技資訊(2015年8期)2015-07-02 18:49:25
