拮抗葡萄灰霉病貝萊斯芽孢桿菌的生長特征及對相關防御酶活性的影響
2023-08-15 01:09:58鄒強牛新湘楊紅梅包慧芳詹發強張雨萌王靜怡佐長賡史應武
食品工業科技 2023年15期
鄒強,牛新湘,劉 萍,楊紅梅,楚 敏,王 寧,林 青,包慧芳,詹發強,張雨萌,王靜怡,佐長賡,婁 愷,史應武,4,5,
(1.新疆大學生命科學與技術學院,新疆烏魯木齊 830052;2.新疆農業科學院微生物應用研究所,新疆烏魯木齊 830091;3.新疆農業科學院土壤肥料與農業節水研究所,新疆烏魯木齊 830091;4.農業農村部西北綠洲農業環境重點實驗室,新疆烏魯木齊 830091;5.新疆特殊環境微生物實驗室,新疆烏魯木齊 830091)
葡萄(Vitis viniferaL.)是葡萄科葡萄屬木質藤本植物,在全世界范圍內分布廣泛,其味道鮮美,營養價值較高。葡萄含水量較高,采后生理代謝旺盛,在運輸和貯藏過程中極易遭受病菌的侵染而引起果實腐爛變質[1]。由灰葡萄孢(Botrytis cinerea)引起的葡萄灰霉病(Gray mold)是危害葡萄果實的主要病害之一,嚴重影響葡萄的品質,造成了巨大的經濟損失[2]。在采后葡萄灰霉病的防治中,通常采用的是SO2、硫化物、吡咯類等化學藥劑,但是長期使用化學藥劑存在很多弊端,如藥劑殘留對人體以及環境的危害、大量使用化學藥劑也增加了病原菌的耐藥性,導致防病效果明顯下降[3?5]。近年來,隨著食品安全問題的日益突出,綠色、安全、高效的生物防治逐漸成為當今果蔬防腐保鮮的新方向[6?7]。
生防微生物在宿主體內的增殖效率是決定生防效果的關鍵因素,是生防菌篩選與評價的重要指標[8]。與病原菌競爭營養物質以及生存空間是生防菌防病機制之一,Bacon 等[9]對玉米內生枯草芽孢桿菌(Bacillus subtilis)研究發現,其與玉米病原菌串珠鐮孢菌(Fusarium moniliforme)有相同的生態位點,枯草芽孢桿菌能快速地在玉米體內定殖,占據病原菌的生長位點,降低玉米的發病率。生防菌的次生代謝拮抗物質是其發揮防病作用的重要機制,高振峰等[10]研究發現貝萊斯芽孢桿菌ZSY-1 產生的脂肽類物質在濃度為200~400 μg/mL 能有效降低番茄采后早疫病的發生、延緩果實軟化。此外,防御酶活性的提高是衡量植物體防御反應的重要指標,也是生防的主要作用機理,有大量研究表明生防菌能夠誘導植物體內相關防御酶活性變化,以此來提高植物的抗病能力[11?13]。陳劉軍等[14]研究證實蠟質芽孢桿菌(Bacillus cereus)AR156 處理水稻后,誘導植株內抗病基因上調表達,增加水稻防御酶活性,從而抑制水稻紋枯病的發生。
有研究表明許多微生物具有防治灰霉病的作用,如枯草芽孢桿菌、多粘類芽孢桿菌(Paenibacillus polymyxa)、解淀粉芽孢桿菌(Bacillus amyloliquefaciens)、酵母菌(Saccharomyces)、熒光假單胞菌(Pseudomonas fluorescens)等[2,15?17]。貝萊斯芽孢桿菌(Bacillus velezensis)是芽孢桿菌屬的一個新種,最早于1999 年被Ruiz 等[18]分離得到,最初認為貝萊斯芽孢桿菌是枯草芽孢桿菌的一個亞種。隨著測序技術的進步,Dunlap 等[19]通過比較基因組學的方法對貝萊斯芽孢桿菌及其相近種進行了分類,認為甲基營養型芽孢桿菌和解淀粉芽孢桿菌植物亞種是貝萊斯芽孢桿菌的異名。研究表明貝萊斯芽孢桿菌抑菌譜廣,具有防治多種植物病害的潛力[20],是一種應用前景廣泛的生防資源。張倩等[21]研究發現貝萊斯芽孢桿菌KT 可通過分泌抗菌物質以及誘導甜櫻桃果實防御酶活性的提高從而抑制甜櫻桃軟腐病的發生。還有研究發現貝萊斯芽孢桿菌SB023 發酵液可顯著抑制杧果炭疽病病原菌的生長,具有良好的生防效果[22]。然而目前關于貝萊斯芽孢桿菌防治葡萄灰霉病的研究鮮有報道,因此,研究貝萊斯芽孢桿菌在葡萄中的定殖能力以及生防機制,對該菌株的進一步開發利用具有重要意義。
本文以前期篩選的貝萊斯芽孢桿菌TP-1 為研究對象,采用抗生素標記法篩選出具有抗利福平標記的抗性菌株,研究其在葡萄中的定殖能力、對葡萄果實腐爛率的影響以及對葡萄中苯丙氨酸解氨酶(PAL)、過氧化物酶(PPO)、抗壞血酸過氧化物酶(APX)等抗病相關酶活性的影響,初步探討貝萊斯芽孢桿菌誘導葡萄對灰霉病產生抗性的機理,為葡萄灰霉病的防治提供一定的理論參考。
1 材料與方法
1.1 材料與儀器
“玫瑰香”葡萄 購自烏魯木齊市沙依巴克區北園春水果市場,選取果實飽滿、無損傷、大小相似、成熟度基本一致的葡萄,立即運回實驗室。貝萊斯芽孢桿菌TP-1 采集新疆五家渠市葡萄園根際土壤,通過稀釋涂布法分離菌株,以灰葡萄孢PH-23 為指示菌,通過平板對峙法篩選得到[23];灰葡萄孢PH-23由本實驗室從葡萄果實病健交界處分離,再回接到葡萄果實上進行驗證,得到此菌株;馬鈴薯葡萄糖瓊脂培養基(PDA)、營養瓊脂(NA)培養基、營養肉湯(NB)培養基、馬鈴薯葡萄糖水培養基(PDB)青島高科技工業園海博生物技術有限公司;苯丙氨酸解氨酶(PAL)試劑盒、多酚氧化酶(PPO)試劑盒、抗壞血酸過氧化物酶(APX)試劑盒 蘇州科銘生物技術有限公司;利福平(Rifampicin)上海生工生物工程有限公司。
ZWY-2102C 型智城恒溫培養振蕩器 上海智城分析儀器制造有限公司;UVZ2450 型紫外分光光度計 日本島津自動化設備有限公司;GMSX–280型高壓蒸汽滅菌鍋 北京市永光明醫療儀器廠;CENTRIFUGE 型高速冷凍離心機 EPPENDORF;RXZ 型智能人工氣候箱 寧波江南儀器廠制造;DH–9162 型電熱恒溫培養箱 上海恒一科技有限公司。
1.2 實驗方法
1.2.1 貝萊斯芽孢桿菌抗利福平標記 參照羅云艷[24]的方法,采用抗生素標記法篩選貝萊斯芽孢桿菌的抗性標記菌株,配制利福平濃度為10、20、30、50、100、150、200、250、300 μg/mL 的NA 培養基。將貝萊斯芽孢桿菌TP-1 接種于含10 μg/mL 利福平的NA 平板中,34 ℃下恒溫培養24 h,待菌落大小約1 cm 時,挑取形態與原始菌株相同的新菌落接種到含有相同濃度抗生素的NA 平板上,繼代培養24 h后,再接入利福平濃度為20 μg/mL 的NA 平板上培養,直到篩選出能在300 μg/mL 利福平下正常生長且菌落形態與原始菌株相同的標記菌株[25?26]。利用平板對峙法,測定標記菌株的對灰葡萄孢的拮抗作用,以原始菌株為對照,最終獲得穩定生長,且對灰葡萄孢的拮抗作用與原始菌株無明顯差異的標記菌株。
1.2.2 標記菌株遺傳穩定性測定 將獲得的標記菌株接種在不添加利福平的NA 培養基上,在34 ℃下繼代培養15 代后接種到利福平濃度為300 μg/mL的NA 培養基上,在相同條件下繼代培養15 代。然后檢測標記菌株在添加利福平和不添加利福平的NA 培養基中的數量,比較其有無明顯差異[27]。
1.2.3 貝萊斯芽孢桿菌生長曲線 根據郗良卿等[28]的方法進行改進,用無菌接種環分別將標記菌株與原始菌株接種于裝有100 mL NB 培養基的500 mL 三角瓶中,在30 ℃、180 r/min 條件下振蕩培養。以未接菌的NB 培養液為空白。0~10 h 每1 h 取一次樣,10 h 后每隔2 h 取一次樣,32 h 結束,用紫外分光光度計在600 nm 波長下測定OD 值,比較標記菌株與原始菌株的生長曲線是否存在差異。
1.2.4 貝萊斯芽孢桿菌在葡萄果實中的定殖能力標記菌株發酵液:將標記菌株接種至含300 μg/mL利福平的NB 培養基中,于30 ℃、180 r/min 條件下振蕩培養24 h,調節濃度至1.0×107CFU/mL。
灰葡萄孢孢子懸浮液:將灰葡萄孢接種于PDA上,27 ℃下培養7 d 后,用無菌水沖洗,然后用滅過菌的紗布進行過濾得到孢子懸浮液,調節濃度到1×106孢子/mL。
參照羅琳等[29]的葡萄處理方法,選取大小和成熟度相近的葡萄果實,用無菌水將葡萄洗凈,再用75%酒精進行表面消毒,然后用無菌水沖洗3 次,自然晾干后,用無菌打孔器在葡萄表面赤道部位打取直徑2 mm、深2 mm 的傷口。吸取10 μL 的拮抗菌液和灰葡萄孢孢子懸浮液接種于葡萄果實傷口內,以只接種拮抗菌液的葡萄為對照。于20 ℃、相對濕度為85%條件下貯藏。以接種后1 h 測定的細菌數量為起始值,而后于接種第5、10、15、20、25、30 d 取樣測定。測定方法為取傷口處果肉組織,放于無菌研缽中研磨,用無菌水稀釋至適宜濃度,取100 μL 稀釋液于含300 μg/mL 利福平的NA 平板上進行稀釋涂布,30 ℃下恒溫培養計數。每個處理3 次重復,每重復10 顆葡萄。
1.2.5 貝萊斯芽孢桿菌對葡萄果實腐爛率的影響選取大小和成熟度相近的葡萄果實,以1.2.4 相同方法進行消毒清洗。用無菌打孔器在果實赤道處打取直徑2 mm、深2 mm 的孔[30]。共分6 個處理:接種10 μL 無菌水為空白對照(CK);只接種10 μL 拮抗菌發酵液(T1);接種5 μL 拮抗菌發酵液,1 h 后接種5 μL 灰葡萄孢孢子懸浮液(T2);接種10 μL 灰葡萄孢孢子懸浮液為病原菌對照組(T3);接種5 μL 灰葡萄孢孢子懸浮液,1 h 后接種5 μL 拮抗菌發酵液(T4);拮抗菌發酵液(5 μL)與灰葡萄孢孢子懸浮液(5 μL)同時接種(T5)。接種后果實置于密封的保鮮盒中,在20 ℃、相對濕度為85%條件下貯藏。每個處理3 次重復,每重復10 顆葡萄。每5 d 統計果實腐爛情況,共統計5 次,計算腐爛率。將病斑直徑大于4 mm 記為發病果實,其相較于總果數的比例記為腐爛率。
1.2.6 防御相關酶活性的測定 選取大小和成熟度相同的葡萄,用無菌打孔器在葡萄果實赤道部位打出一個直徑、深淺相同的傷口(2 mm)。實驗選取1.2.5 中CK、T1、T2、T3 4 個處理組。常溫下貯藏,相對濕度85%。每個處理3 次重復。在貯藏第5、10、15、20、25 d 取樣檢測,按照試劑盒步驟測定酶活性。
PAL 酶活單位定義:每g 組織在每mL 反應體系中每min 使290 nm 下吸光值變化0.05 為一個酶活性單位(U/g FW)。PPO 酶活單位定義:每min 每g組織在每mL 反應體系中使525 nm 處吸光值變化0.005 為一個酶活性單位(U/g FW)。APX 酶活單位定義:每g 組織每min 氧化1 nmol AsA 為1 個酶活性單位(nmol/min/g FW)。
1.3 數據處理
本研究每處理3 個平行,每平行3 次重復,數據以平均值±標準差表示,采用Excel 2016 和SPSS 20.0 軟件進行數據分析,應用Duncan 新復極差法進行顯著性檢驗(P<0.05)。使用Origin 2020 軟件繪圖。
2 結果與分析
2.1 標記菌株穩定性實驗
2.1.1 抗生素標記菌株的篩選 采用抗生素標記法獲得可以在利福平濃度為300 μg/mL 的平板上正常生長的標記菌株TP-1R,并且其菌落形態沒有發生明顯改變。圖1(a)為標記菌株TP-1R在含利福平濃度為300 μg/mL NA 平板的菌落圖。圖1(b)顯示標記菌株抑菌圈直徑為22.14±0.24 mm,原始菌株抑菌圈直徑為22.48±0.16 mm,兩者無明顯差異,表明經利福平標記的標記菌株與原始菌株對病原菌的拮抗能力無顯著差異(P>0.05)。
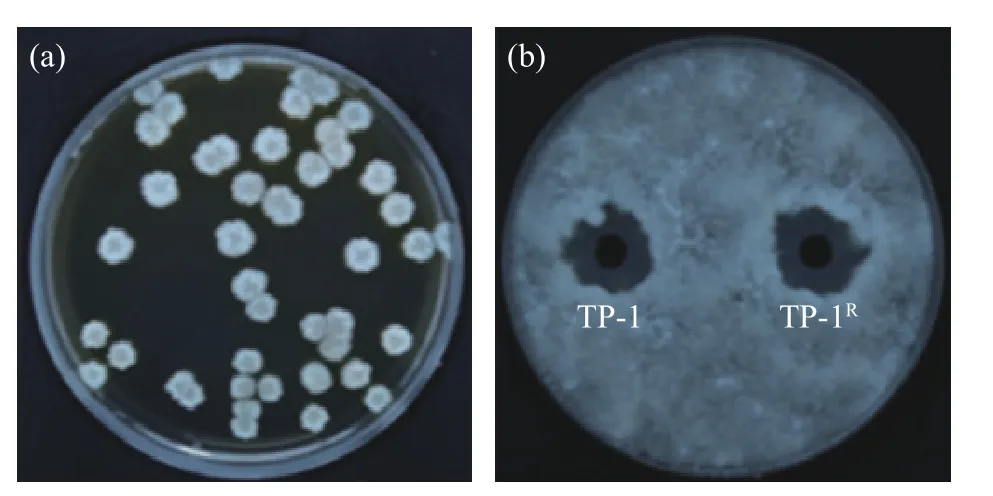
圖1 標記菌株的篩選Fig.1 Screening of labeled strains
2.1.2 標記菌株遺傳穩定性分析 標記菌株在含利福平的NA 培養基中傳代培養15 代,其第5、10、15 代菌落數分別為1.0×109、1.01×109、0.97×109CFU/mL,在不含利福平的NA 培養基中第5、10、15 代菌落數分別為1.0×109、0.99×109、0.95×109CFU/mL,兩者間無明顯差異(P>0.05),均能穩定生長(圖2)。
圖2 標記菌株TP-1R 的抗性遺傳穩定性Fig.2 Genetic stability of resistance of marker strain TP-1R
2.1.3 標記菌株與原始菌株生長曲線 從圖3 可以看出標記菌株與原始菌株的生長曲線基本一致,3 h后進入對數生長期,在培養18 h 時標記菌株OD 值達到峰值,為3.986,原始菌株OD 值為4.063,差異不顯著(P>0.05)。結果表明標記菌株的抗性穩定,能夠用于后續實驗。
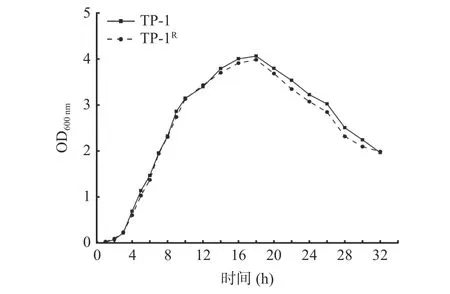
圖3 標記菌株與原始菌株的生長曲線Fig.3 Growth curve of labeled strain and original strain
2.2 貝萊斯芽孢桿菌在葡萄果實中的定殖及其防腐效果
2.2.1 貝萊斯芽孢桿菌在萄果實中的定殖 由圖4可知,兩個處理組在葡萄果實上的數量隨著貯藏時間的延長呈現先下降后上升再下降的趨勢。可能是由于拮抗菌在接種后需要適應其生態位點并與病原菌進行競爭,定殖數量有所下降,在拮抗菌適應生態位點后,定殖數量開始上升,在宿主營養消耗殆盡,拮抗菌定殖數量開始逐漸下降[31]。不同時期T 處理組回收到的標記菌株數量均多于T+P 處理組,都在第15 d時定殖數量達到最高值,T 處理組為4.32×106CFU/g,T+P 處理組為4.08×106CFU/g。分別是初始值的1.23倍和1.20 倍。隨著時間的延長,標記菌株的數量也逐漸下降,其中在第30 d 時T 處理組和T+P 處理組可以回收到的標記菌株數量分別為3.11×106CFU/g,2.93×106CFU/g,表明貝萊斯芽孢桿菌TP-1R在葡萄上具有較強的定殖能力。
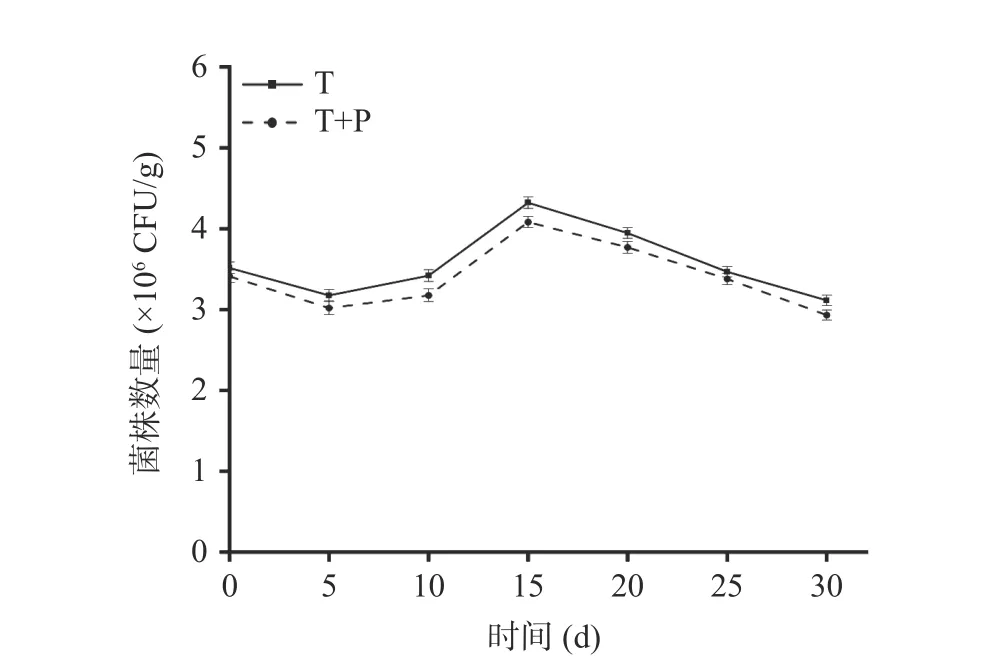
圖4 拮抗菌TP-1R 在葡萄中的定殖動態Fig.4 Colonization dynamics of antagonistic TP-1R in grape
2.2.2 貝萊斯芽孢桿菌對葡萄果實腐爛率的影響腐爛率能夠直接反映果實的感官品質,是影響果實貯藏品質的重要指標。由圖5 可以看出,在整個貯藏期間玫瑰香葡萄腐爛率呈現逐漸上升的趨勢。T1 處理組的葡萄腐爛率要顯著低于其他處理組(P<0.05),在第5 d 時T1 處理組無腐爛果實,在貯藏第25 d 時T1 處理組腐爛率為35.76%,CK、T2、T3、T4、T5 處理組腐爛率分別為81.84%、39.14%、100%、53.39%、44.28%。其中T2 處理組腐爛率要低于T5 處理組,說明提前接種貝萊斯芽孢桿菌可有效抑制病原菌生長,減緩貯藏期間葡萄果實的腐爛。
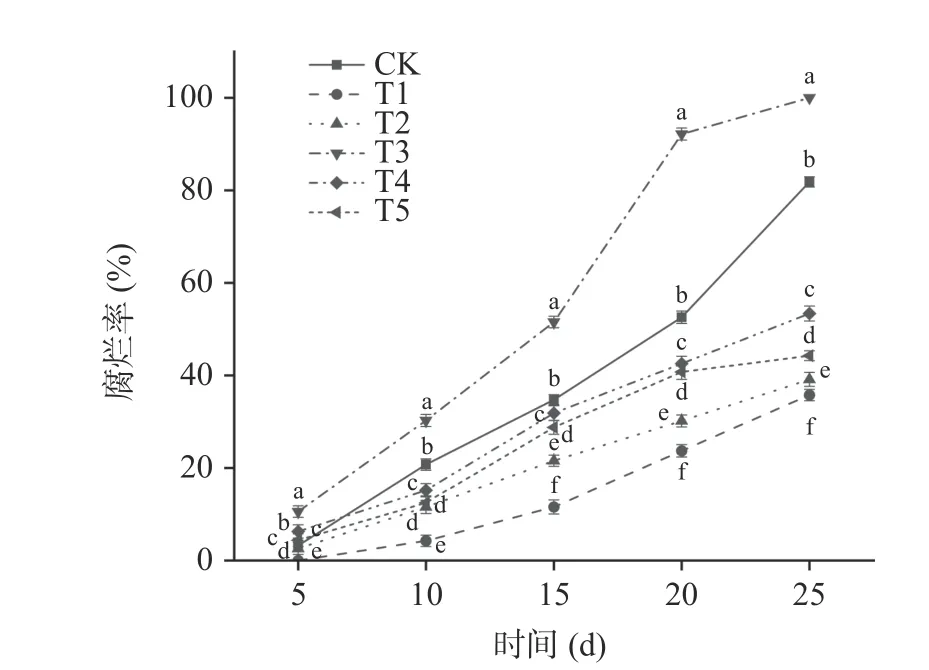
圖5 貝萊斯芽孢桿菌TP-1R 對葡萄腐爛率的影響Fig.5 Effect of Bacillus velezensis TP-1R on decay rate of grape
2.3 防御相關酶活性的變化
2.3.1 苯丙氨酸解氨酶(PAL)活性的變化 由圖6可以看出,4 種不同處理下,葡萄中PAL 的活性都隨時間延長呈現先上升后下降的趨勢。初期在受到病原菌侵染,激活宿主防御反應,PAL 活性逐漸上升,后期宿主體內代謝減緩,宿主體內酶活也逐漸下降。在貯藏第15 d 時,4 個處理組葡萄中PAL 活性達到最大值,其中,T2 處理組的葡萄PAL 活性最大,為6.80 U/g FW,是對照組(CK)的1.41 倍。T1 處理組葡萄PAL 活性為5.96 U/g FW,是對照組(CK)的1.23倍。T2 處理組PAL 活性峰值明顯大于T1 處理組PAL 活性峰值(P<0.05)。表明混合接種對葡萄PAL誘導活性要大于單獨接種貝萊斯芽孢桿菌TP-1R對葡萄PAL 誘導活性,可能是因為兩者在誘導PAL 酶活中具有協同作用。拮抗菌TP-1R處理葡萄后能有效減緩PAL 活性的降低。
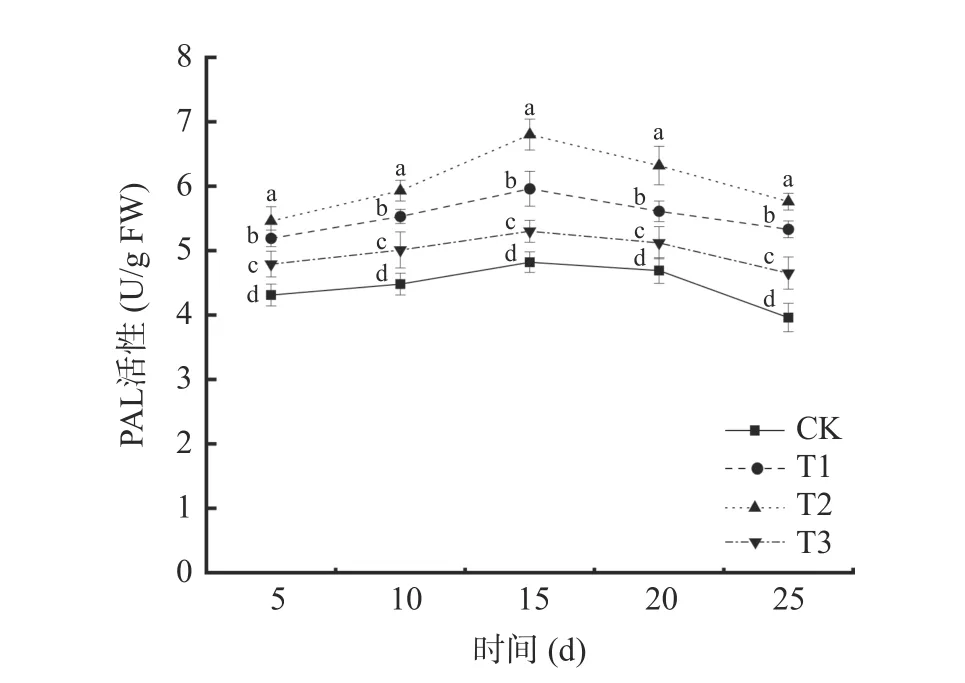
圖6 不同處理對葡萄PAL 活性的影響Fig.6 Effects of different treatments on PAL activity of grape
2.3.2 多酚氧化酶(PPO)活性的變化 由圖7 可知,在貯藏期間,不同處理下的葡萄PPO 活性呈現先上升后下降的趨勢。T1 處理與T2 處理均能提高葡萄PPO 活性,除第10 d 二者無顯著差異外(P>0.05),T1 處理對葡萄PPO 的誘導效應均大于T2 處理。四個處理的PPO 活性均于貯藏第15 d 達到峰值,其中T1 處理的PPO 活性最大,為73.15 U/g FW,是對照組(CK)的1.19 倍;T2 處理組葡萄PPO 活性為68.11 U/g FW,是對照組(CK)的1.11 倍。
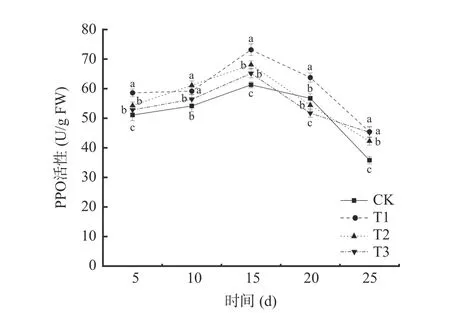
圖7 不同處理對葡萄PPO 活性的影響Fig.7 Effects of different treatments on PPO activity of grape
2.3.3 抗壞血酸過氧化物酶(APX)活性的變化從圖8 可以看出,在貯藏期間,不同處理下葡萄APX活性呈先上升后下降的趨勢,且T1 和T2 處理組葡萄APX 活性顯著高于其他處理組(P<0.05)。各處理組的APX 活性均于貯藏第15 d 達到峰值,T1 處理為816.51 nmol/min/g FW,T2 處理為703.58 nmol/min/g FW,分別是對照組(CK)的2.01 倍和1.74 倍。單獨接種貝萊斯芽孢桿菌對葡萄APX 誘導效應要顯著高于其他處理組(P<0.05)。表明貝萊斯芽孢桿菌處理能有效減緩葡萄APX 活性的降低。
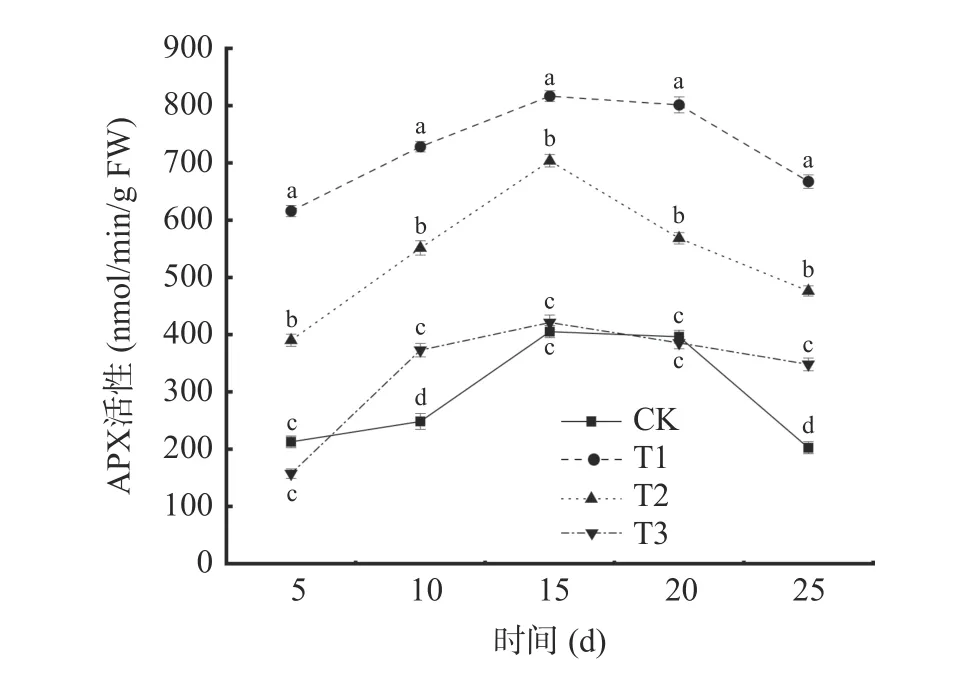
圖8 不同處理對葡萄APX 活性的影響Fig.8 Effects of different treatments on APX activity of grape
3 討論
生防菌是否可以在宿主體內穩定定殖以及定殖數量是影響生防效果的關鍵因素。目前用于研究生防菌在宿主體內定殖情況的方法主要有:抗生素標記法、同位素示蹤法、基因標記法等。其中抗生素標記法具有簡單、快速、實用、不會改變原始菌株的重要性質等特點,適用于追蹤微生物在宿主體內的定殖情況[8,12,27,32]。本研究通過用抗生素標記法,篩選出了能抗300 μg/mL 利福平的標記菌株TP-1R,其拮抗性能、外觀形態等均與原始菌株無明顯差異,且抗性標記可穩定遺傳。本研究發現貝萊斯芽孢桿菌在葡萄中的定殖規律呈現先下降后上升再下降趨勢。在貯藏后期其定殖數量還可達到3.11×106CFU/g,表明菌株TP-1R能在葡萄上穩定定殖。生防菌在宿主上的定殖是一個十分復雜的動態過程,溫度、接種量、宿主自身特性等因素都會對其產生影響[33]。其中生防菌能否很快適應宿主所在的微生態環境,從而占據有利位點,抑制病原微生物的生長是其有效發揮生防作用的重要因素。目前只是在實驗室環境下研究了貝萊斯芽孢桿菌的定殖能力,為了進一步地提高其在實際應用的效果,還需要對影響其定殖量的因素做更深入的研究。
誘導系統抗性是生防菌發揮防病作用的重要機制之一。參與植物體內生理代謝的APX、PPO、PAL、POD 等酶系與植物抵抗病原菌侵染有著密切關系。APX 是植物活性氧代謝中重要的抗氧化酶之一,APX 酶能有效提高植物的活性氧代謝水平,并且能促進維生素C 的代謝水平,有效清除植物體內過多的活性氧,提高APX 的活性可以減輕果實的氧化損傷,降低腐爛率[4]。其中PAL 是苯丙烷代謝途徑的關鍵酶,參與酚類、植保素和木質素等抗菌物質的合成[34],已有研究表明酚的代謝產物是潛在的抗病因子,而PPO 酶能將酚類物質氧化為活性更強的醌類物質,并能促進木質素的合成,增加植物的抗病能力[35?36]。已有大量研究報道芽孢桿菌能夠誘導植物防御酶活性的提高,從而抵抗病原菌的侵染。孫一凡等[37]發現側孢芽孢桿菌BI13 可以誘導番茄葉片防御酶活性的提高增強植物對番茄早疫病的抗性。孫建波等[38]研究發現經枯草芽孢桿菌XB16 處理后的香蕉,其POD 和PPO 活性均比接種病原菌和清水對照高。王雪等[39]采用灌根法對人參進行處理,發現解淀粉芽孢桿菌FS6 能夠提高人參CAT、PPO、POD 酶活性,增強人參對根腐病菌的抗性。本研究中發現,葡萄果實在受到灰葡萄孢侵染時,與對照相比葡萄中PAL、PPO、APX 活性略有上升,表明病原菌也可以誘導植物相關防御酶活性的提高,可能原因是葡萄在受到病菌入侵后,為了抵御病菌,體內防御酶基因被激活,相關防御酶上調表達。此外經拮抗菌處理和拮抗菌與病原菌共同處理的葡萄防御酶活性均高于對照組以及灰葡萄孢處理,防御酶活性有更大程度的增加,說明菌株TP-1R能誘導植物進一步提高防御酶活性,增強對灰葡萄孢的抗性。且拮抗菌和病原菌共同誘導的PAL 活性要高于單獨接種拮抗菌和病原菌,表明兩者共同誘導對PAL 活性有協同增效的作用。防御酶的活性是植物抗病性的重要生理特征,酶活性越高,抗病性越強。此外防御酶相關基因的增強有助于防御系統的激活,防御酶相關基因的表達水平與相應酶活性密切相關[4,40]。關于貝萊斯芽孢桿菌TP-1R誘導相關防御酶基因的表達水平以及關于其抑菌物質還有待進一步研究。
4 結論
本研究通過抗生素標記法測定貝萊斯芽孢桿菌TP-1R在葡萄上定殖能力,發現其在貯藏第30 d 菌株定殖數量仍可達到3.11×106CFU/g,表明拮抗菌TP-1R能夠在葡萄果實穩定定殖,從而有效發揮其生防效果。后續課題組將采用GFP 基因標記法,研究菌株TP-1R在葡萄生長期間的定殖動態,對本實驗進行補充驗證。在接種拮抗菌發酵液后,能有效防治葡萄灰霉病,降低葡萄果實腐爛率,在貯藏第25 d 時葡萄腐爛率為35.76%。在貯藏期間,葡萄防御酶PAL、PPO、APX 均呈先上升后下降的趨勢,接種拮抗菌TP-1R的葡萄3 種酶活性顯著高于CK(P<0.05),在第15 d 時,拮抗菌處理組PAL、PPO、APX 活性是CK 的1.23、1.19、2.01 倍。貝萊斯芽孢桿菌TP-1R處理能夠提高葡萄防御酶PAL、PPO、APX 的活性,增強葡萄對灰霉病的防御能力。本研究為利用該菌株進行生防菌劑的研制提供了理論依據,為后續研究貝萊斯芽孢桿菌對葡萄防御物質合成相關基因表達奠定了基礎。
