醬香型白酒高溫大曲儲存過程中微生物群落演替與理化因子相關性研究
2023-08-15 01:10:02謝丹畢遠林胡建鋒馮小兵程平言
食品工業科技 2023年15期
謝丹,吳 成,畢遠林,胡建鋒,黃 魏,胡 峰,馮小兵,程平言
(貴州習酒股份有限公司,貴州習水 564622)
“酒乃曲之骨”,醬香型白酒發酵過程中的微生物主要源于高溫大曲[1],大曲是一種富含多酶多菌的微生物發酵劑,具有“糖化、發酵、生香”等功能,是白酒發酵過程的重要物質保障,其品質的好壞決定了醬香型白酒的產量和質量[2?5]。
醬香型高溫大曲采用生料制曲、添加母曲自然接種、經40 d 發酵逐漸形成了以細菌、霉菌和酵母菌為主的群落結構[6]。醬香型高溫大曲制作完成后,需進行拆曲入庫儲存,使大曲進入“后熟”階段。合理的儲存期對醬香型白酒釀造有至關重要的作用。在大曲儲存的理化特性研究方面,張麗等[7]研究結果表明高溫大曲儲存5~6 個月時,液化力、酯化力等理化特性趨于穩定;劉建文等[8]發現儲存3~4 個月的大曲,其功能酶活力均達到最好狀態。在微生物多樣性研究方面,粱麗文等[9]研究結果表明高溫大曲存儲過程中主要優勢細菌屬為Bacillus,真菌屬為Pichia;梁晨等[10]研究發現大曲貯存4~6 個月,更有利于白酒的釀造,且Thermoactinomyces、Bacillus等是大曲儲存過程中的優勢細菌屬,未對真菌多樣性進行相關研究。陳曉茹等[11]研究表明強化曲在貯存過程中的優勢菌屬是Bacillus、常規曲的優勢菌屬為Weissella。目前,對醬香型白酒釀造過程中的微生態結構分析方法主要是傳統可培養方法,但該方法耗時長,準確性較低,且具有一定的局限性,遠遠不能滿足對微生物群落結構和多樣性的認識,但是傳統的可培養方法在微生物檢測上的作用是無法替代的。然而隨著分子生物學技術的快速發展,能快速精確地揭示微生物種類的多樣性,鑒定樣本中微生物群落組成的能力,可檢測到未發現的微生物,更加全面地了解系統中的微生物,客觀地認識微生物的群落結構及其演替變化。對于高溫大曲“后熟”質量的評價主要依靠感官和理化指標,且在儲存過程其影響因素有待于進一步闡明。在實際生產中,大曲是否成熟主要依靠生產經驗判斷,存在著盲目性和不確定性。
因此,本文以“后熟”過程的醬香型白酒高溫大曲為研究對象,同時采用傳統可培養方法和高通量測序技術解析細菌和真菌群落組成,并結合高溫大曲中各理化因子動態變化規律,分析微生物群落和理化指標間的相互作用關系,以期為闡明醬香型白酒高溫大曲“后熟”過程各因素變化機理提供理論參考。
1 材料與方法
1.1 材料與儀器
高溫大曲樣品采集自GZXJ 制曲車間不同儲存時間成品曲。孟加拉紅培養基、營養瓊脂 北京奧博星生物技術有限責任公司;WL 培養基 上海博微生物科技有限公司;DNA 提取試劑盒 美國OMEGA BioTek 公司;聚合酶鏈式反應引物 上海翌圣生物科技股份有限公司。
SW-CJ-2F 無菌操作臺 蘇州凈化設備有限公司;ZQPW-70 全溫振蕩培養箱 天津市萊玻特瑞儀器設備有限公司;KG-AP32L 自動蒸汽滅菌器 日本ALP 公司;GL-88B 漩渦混合器 海門市其林貝爾儀器制造有限公司;TND03-H-H 混勻型干式恒溫器 深圳托能達科技有限公司;ETC811 PCR 儀 北京東勝創新生物科技有限公司;DYCZ-21 電泳儀北京市六一儀器廠;FR-1000 凝膠成像系統 上海復日科技有限公司。
1.2 實驗方法
1.2.1 樣品采集 選取GZXJ 公司制曲車間某一儲存曲庫,在儲存過程中進行定點跟蹤取樣,每隔1 月取1 次,共計6 個樣品。樣品磨碎后置于無菌密封袋中,于?20 ℃下低溫保藏,按取樣時間依次標記為GX-1(0 d)、GX-2(30 d)、GX-3(60 d)、GX-4(90 d)、GX-5(120 d)、GX-6(150 d)。
1.2.2 理化指標的測定 溫度、水分、酸度、糖化力、液化力的測定具體參照QB/T 4257-2011《釀酒大曲通用分析方法》[12]。
1.2.3 可培養微生物的分離保藏、計數及鑒定 a.分離及保藏:稱取10 g 樣品于90 mL 無菌水中,于25 ℃、180 r/min 條件下振蕩30 min,吸取1 mL 懸浮液稀釋備用,分別采用營養瓊脂、WL 培養基和孟加拉紅培養基分離篩選細菌、酵母菌和霉菌,觀察菌落在培養基上的形態。細菌和酵母菌采用?80 ℃甘油保藏,絲狀真菌采用4 ℃斜面保藏,每株菌株平行保藏3 管。
b.計數:采用稀釋涂布的方法,分別采用營養瓊脂培養基于37 ℃條件下培養細菌;WL 培養基和孟加拉紅培養基于28 ℃條件下培養酵母菌和絲狀真菌培養結束后,用計數器對肉眼觀察的單菌落計數,記錄稀釋倍數和相應的菌落數量。
c.鑒定:根據菌落形態特征進行初步分類后,每種類別挑選代表性菌株1~3 株[13]。分別采用Ezup柱式基因組DNA 試劑盒對菌株DNA 進行提取,具體操作流程見試劑盒說明書。參照吳成等[14]方法,細菌采用通用引物27F(AGTTTGATCMTGGCTC AG)和 1492R(GGTTACCTTGTTACGACTT)對16S rRNA 基因擴增測序;酵母菌采用通用引物NL1(GCATATCAATAAGCGGAGGAAAAG)和 NL4(GGTCCGTGTTTCAAGACGG)對26S rRNA 基因D1/D2 片段擴增測序;絲狀真菌采用通用引物ITS1(TCCGTAGGTGAACCTGCGG)和ITS4(TCCTCCG CTTATTGATATGC)對內轉錄間隔區1 和2 片段擴增測序。
PCR 反應體系為:模板DNA 0.5 μL,10×Buffer(with Mg2+)2.5 μL,dNTP2.5 μL,Taq 酶0.2 μL,正向及反向引物各0.5 μL,使用ddH2O 補齊體系至25 μL。PCR 擴增條件為:94 ℃預變性4 min;94 ℃變性45 s;55 ℃退火45 s;72 ℃延伸1 min;30 次循環;72 ℃終延伸10 min 降至4 ℃。擴增完成后,回收PCR 產物并純化。
1.2.4 大曲總DNA 提取及擴增 總DNA 提取:按照DNA 試劑盒E.Z.N.A? Mag-Bind Soil DNA Kit 提取試劑盒方法提取DNA。細菌16S V3~V4 區域片段,選擇341F(5'-CCTACGGGNGGCWGCAG-3')和805R(5'-GACTACHVGGGTATCTAATCC-3')為擴增引物;真菌采用ITS3(GCATCGATGAAGAAC GCAGC)和ITS4(TCCTCCGCTTATTGATATGC)為擴增引物。
PCR 擴增條件為:95 ℃預變性3 min;94 ℃變性20 s;55 ℃退火20 s;72 ℃延伸30 s;25 次循環;72 ℃終延伸5 min 降至4 ℃。PCR 反應體系為:2×Hieff? Robust PCR Master Mix 15 μL,正向及反向引物各1 μL,DNA 模板20~30 ng,使用ddH2O 補齊體系至30 μL。
1.2.5 高通量測序 采用Illumina Miseq 測序平臺,分別對細菌16S rRNA 基因 V3~V4 區、真菌ITS3~ITS4 區進行高通量測序分析(由上海生物工程有限公司完成)。
1.3 數據處理
使用UNITE 為參考數據庫進行物種注釋,基于Silva 庫對97%相似水平的OTU 代表序列進行分類學注釋分析,細菌和真菌分別采用RDP 和UNITE數據庫進行比對。采用Microsoft Office Excel 2016進行數據統計和處理;理化因子動態圖、可培養微生物動態圖和物種相對豐度柱狀圖采用Origin 2017軟件進行處理繪制;優勢微生物和理化因子之間相關性熱圖先利用SPSS 22.0 軟件計算Spearman 相關性系數,再使用OmicShare Tools 平臺繪制組間動態相關性熱圖;動態豐度Venn 圖使用micShare Tools平臺繪制;冗余分析(Redundancy analysis,RDA)采用Canoco5.0 軟件。
2 結果與分析
2.1 儲存過程中理化指標的動態變化
大曲中的水分、酸度、液化力、糖化力等是評價大曲品質好壞常用的理化指標。由圖1 可知,隨著儲存時間的變化,儲存倉溫度先上升后下降,曲坯的水分、酸度逐漸下降,糖化力呈現先上升后下降的趨勢,液化力在(0.02±0.01)~(0.04±0.02)g/g·h 之間波動。微生物可以利用大曲中淀粉、脂肪以及蛋白質降解成小分子進而代謝產酸,隨著儲存過程中曲坯水分逐漸揮發,微生物的生長繁殖受到影響,導致酸度下降。有研究表明,大曲的酸度主要來源于生酸微生物降解蛋白質、脂肪、淀粉等物質和有機酸代謝產酸,是微生物綜合作用的結果[15~17]。大曲的糖化力受工藝控制參數的影響,并且與產品風格密切相關[18]。由圖1 可知,隨著儲存時間的變化,糖化力先升高后下降,在儲存120 d(GX-5)時達最高,為248.97±0.02 mg/g·h。大曲的液化力反映大曲中液化型淀粉酶將淀粉水解為糊精的能力,主要是由霉菌和枯草芽孢桿菌產生[19]。
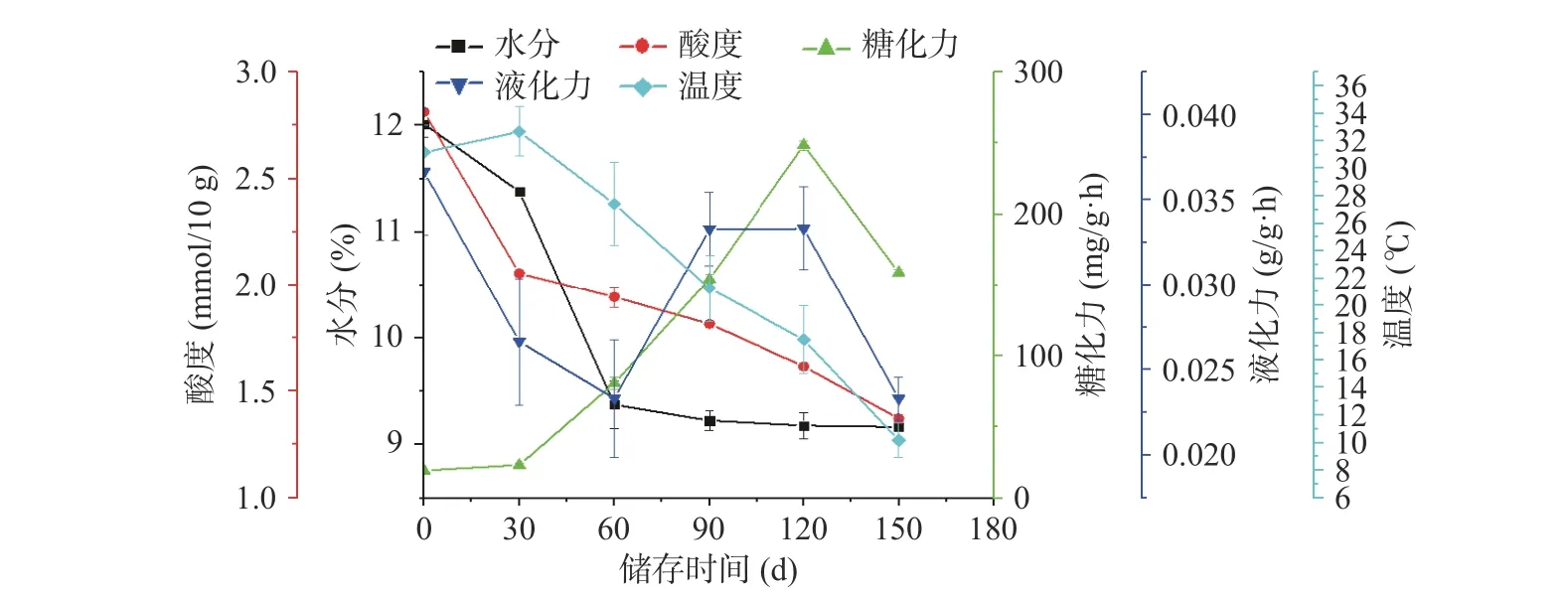
圖1 理化指標的變化規律Fig.1 Variation law of physical and chemical indexes
2.2 可培養微生物的動態變化
采用稀釋涂布平板法對儲存0~150 d 的可培養微生物進行計數、分離,其結果分別表1、圖2 所示。
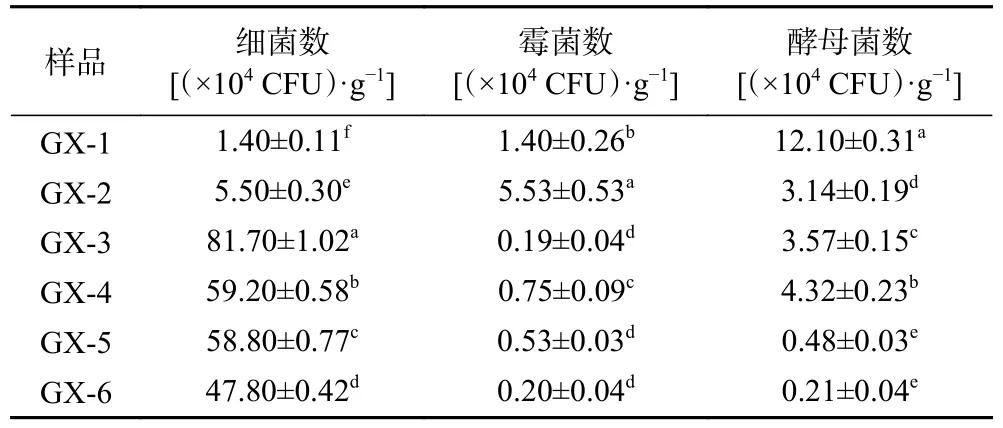
表1 可培養微生物菌落總數變化Table 1 Changes in the total number of culturable microorganisms
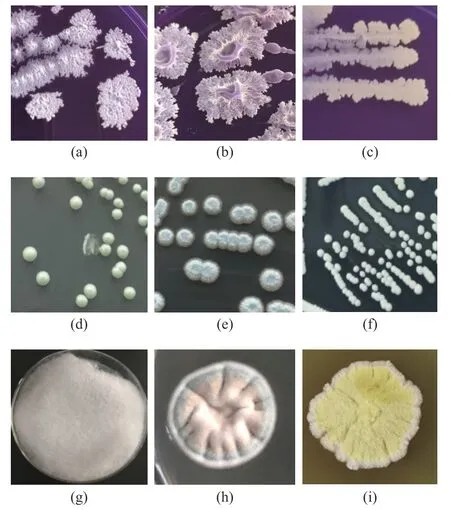
圖2 可培養方法分離篩選微生物形態特征Fig.2 Microbial characteristics revealed by culture-dependent approaches
由表1 可知,大曲初入儲存倉時,細菌和霉菌均在104CFU/g 數量級,酵母菌在105CFU/g 數量級;大曲儲存到60 d(GX-3)時,可培養微生物數量最高,90 d(GX-4)時可培養微生物的數量緩慢下降。分析可能原因是在大曲初入曲房時,儲存倉溫度較低,隨著儲存時間增加,曲房溫度升高,此階段大多數的芽孢細菌在此時形成芽孢,有利于微生物生長繁殖。后期隨著儲存時間的變化,曲坯的水分降低,儲存倉溫度下降,微生物數量緩慢下降。大曲在儲存過程中受成品曲倉、曲坯內部溫度、水分、酸度等參數的影響,同時也受曲坯內部微生物生態結構變化的調控,儲存過程中存在著一個雙向調控機制[20?21]。
本研究采用稀釋涂布平板法分離篩選到15 株細菌、6 株酵母菌和12 株霉菌,共保藏菌株99 株。選擇代表性菌株進行分子鑒定為B.licheniformis、B.amyloliquifaciens、S.gallinarum三種細菌(a~c);T.ciferrii、M.farinosa、S.cerevisiae三種酵母菌(d~f);L.ramosa、A.sydowii、A.chevalieri(g~i)三種霉菌,其形態特征如圖2 所示。相關研究表明,高溫大曲中分離出的優勢菌種也具有一定的相似性,王曉丹等[22]研究表明茅臺地區醬香型高溫大曲中的主要優勢細菌屬為Bacillus;張立強等[23]從瀘州老窖大曲中分離出高產纖溶酶的優勢菌B.licheniformis:李子鍵等[24]從中高溫大曲中分離出耐高溫的微小根毛霉、布氏橫梗霉,并且分別具有產糖化酶、蛋白酶活性功能。
2.3 大曲在不同儲存時間免培養微生物的變化情況
2.3.1 Alpha 多樣性指數分析 樣品測序完成后,得到的原始序列經去除嵌合體、過濾等處理后共得到498207 條有效序列,其中細菌202228 條、真菌295979 條。細菌和真菌測序覆蓋率都為0.99 以上,表明測序深度足夠,樣品中幾乎所有樣本都被檢出,測序結果可真實反映出醬香型高溫大曲在儲存過程中細菌和真菌群落多樣性組成情況。對于細菌而言,Shannon 指數呈先升后降,Simpson 指數先降低后趨于平穩,表明大曲群落多樣性在儲存60 d(GX-3)、90 d(GX-4)、120 d(GX-5)時最高;對于真菌而言,Shannon 指數呈緩慢下降趨勢,Simpson 指數呈逐漸升高趨勢,表明隨著大曲儲存時間延長,真菌群落多樣性逐漸降低(表2)。

表2 不同儲存時間真菌和細菌Alpha 多樣性Table 2 Alpha diversity of fungi and bacteria at different storage times
2.3.2 OTU 分布的Venn 圖 Venn 圖可以用來統計樣本中共有和獨有的OTU 的數目,直觀地展現樣品OTU 數目組成的相似性及重疊情況[25]。如圖3(a)所示,樣品GX-1(0 d)到GX-6(150 d)獨有的OTU數目依次為7、8、6、23、8 和14,其中GX-4(90 d)樣本中獨有的物種數所占比例高,那么樣本之間的相似性差,該樣本的獨特性高;6 個樣品共有的 OTU為11 個,其所代表的細菌物種長期存在于儲藏期內。圖3(b)所示,樣品GX-1(0 d)到GX-6(150 d)獨有的OTU 數目依次為3、1、2、11、5 和47,共有的OTU 為7 個,其中GX-6 樣本獨有的物種數所占比例高,獨特性高,GX-1~GX-5(0~120 d)與共有樣本的數量接近,說明共有物種的占比高,因此樣本的相似性就高。
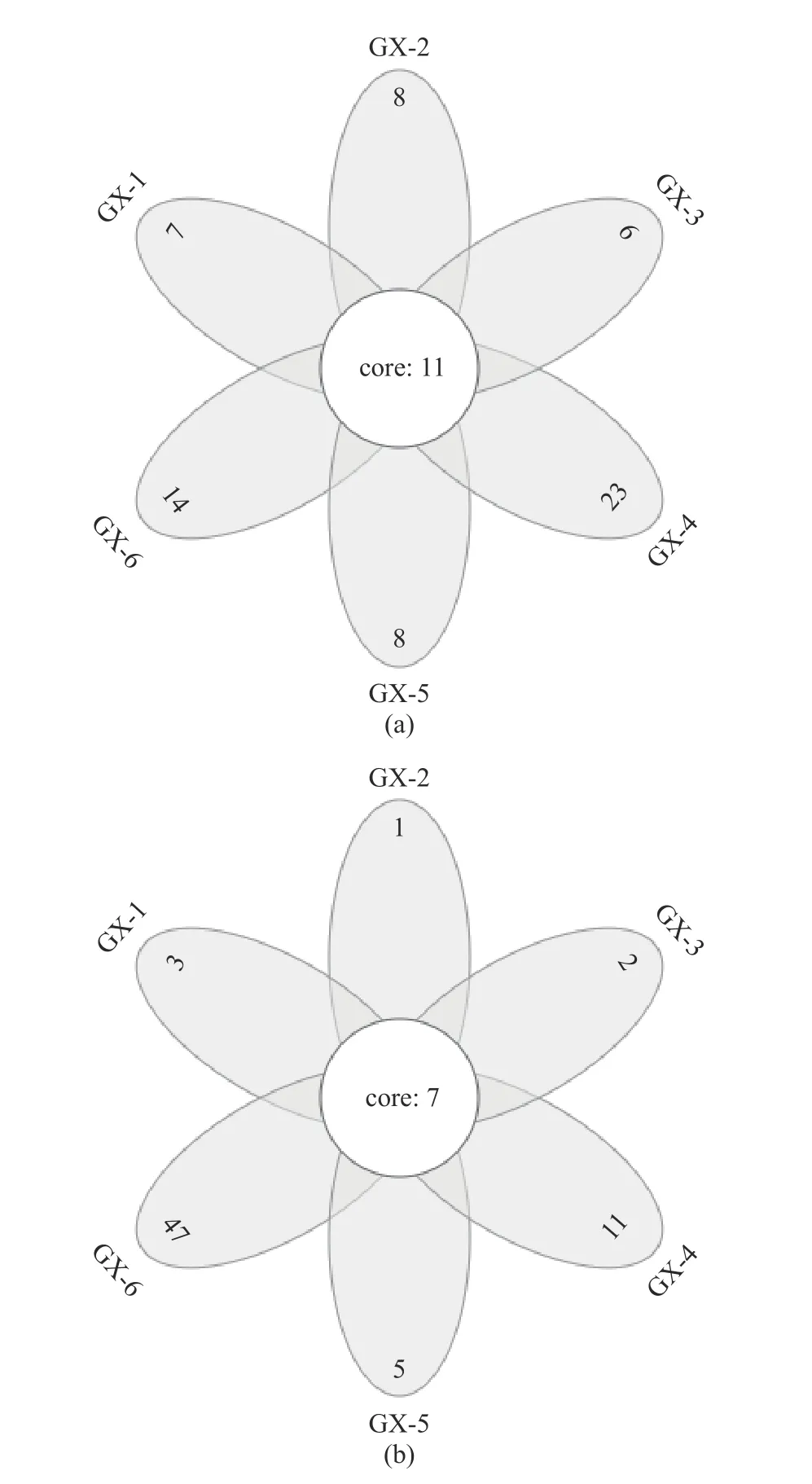
圖3 不同樣品的細菌(a)、真菌(b)OTU Venn 圖Fig.3 Venn profile of OTU of bacteria (a) and fungi (b) in different samples
2.3.3 不同儲存時間細菌、真菌群落動態變化 樣品經高通量測序后,細菌共檢出12 個門、21 個綱、43 個目、80 個科、119 個屬和179 個種,其中Firmicutes 為絕對優勢細菌門,儲存過程中平均占比71.69%;Kroppenstedtia(平均占比30.45%)、Lactobacillus(平均占比11.53%)、Staphylococcus(平均占比10.31%)為主要優勢菌屬。真菌共檢5 個門、17 個綱、37 個目、89 個科、142 個屬和203 個種,其中Ascomycota 為絕對優勢真菌門,平均占比98.11%,Thermomyces(43.87%)、Trichomonascus(29.77%)為主要優勢真菌屬。圖4、圖5 分別為平均相對豐度至少在一個大曲樣品中大于1%的菌群結構。
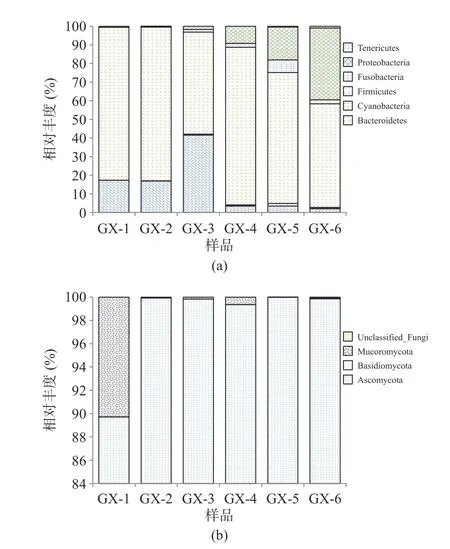
圖4 大曲在門水平上細菌(a)、真菌(b)組成情況Fig.4 Composition of bacteria (a) and fungi (b) in hightemperature Daqu at the phylum
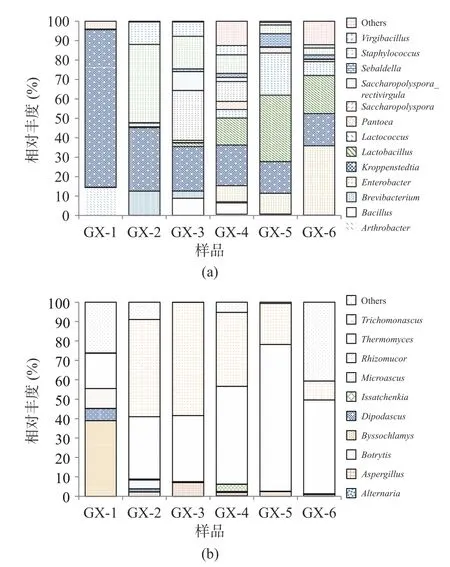
圖5 大曲在屬水平上細菌(a)、真菌(b)組成情況Fig.5 Composition of bacteria (a) and fungi (b) in hightemperature Daqu at the genus
由圖5 可知,Lactobacillus、Lactococcus、Saccharopolyspora、Virgibacillus細菌屬和Thermomyces、Trichomonascus、Aspergillus真菌屬物種多樣性呈現先增后降,在儲存到120 d(GX-5)以后,優勢菌群的相對豐度含量逐漸下降。分析造成這種現象可能的原因是儲存時間過長的曲塊,曲蟲增多,使大曲中的淀粉等物質流失,破壞大曲的微生物群落結構,因而導致微生物多樣性降低[26]。
醬香制曲過程中的細菌主要來源于空氣、場地、原輔料及人體,在醬香型白酒釀造過程中的主要作用是產香,在發酵體系中能產生大量酶類,是形成醬香型白酒風味前體物質的主要菌群[27?28]。相關研究表明,Bacillus是醬香型白酒發酵過程中主要的功能細菌種群,具有代謝水解酶、乙偶姻等風味物質功能,對醬香型白酒的風味形成有重要貢獻[29?30];Lactobacillus屬于乳酸菌菌系,大多是兼性厭氧或厭氧菌,在發酵過程具有重要的生物學調控作用[31]。真菌方面,釀酒酵母是產生酒精的主要功能菌種,曲霉菌和根霉菌具有很強的酶活力,且絕大多數的真菌屬于嗜熱真菌屬,可以在高溫下進行生長代謝,產酶豐富,具有分解酒醅中大分子物質、產生白酒中風味化合物及其前提物質的作用[32]。
2.3.4 理化因子與微生物群落相關性分析 理化因子的變化對微生物的生長繁殖及其行為具有較強的影響,進而調控微生物群落的結構及其演替[21]。如圖6(a)所示,RDA 分析結果表明大曲在儲存0 d(GX-1)時,大曲的后熟過程與溫度、水分、液化力和酸度呈正相關關系,與糖化力呈負相關關系,表明該時期溫度和水分適宜,比較適合微生物生長;儲存30~60 d(GX-2~GX-3)時,大曲中的微生物群落結構不受水分、酸度等因素的影響,表明該時間段理化指標對微生物群落結構影響不大;儲存90~150 d(GX-4~GX-6)時,溫度、水分、酸度、液化力與儲存過程呈負相關,表明了大曲在存儲后期,曲倉溫度降低,曲坯水分含量低,不適宜微生物代謝活動,因此,微生物多樣性降低。
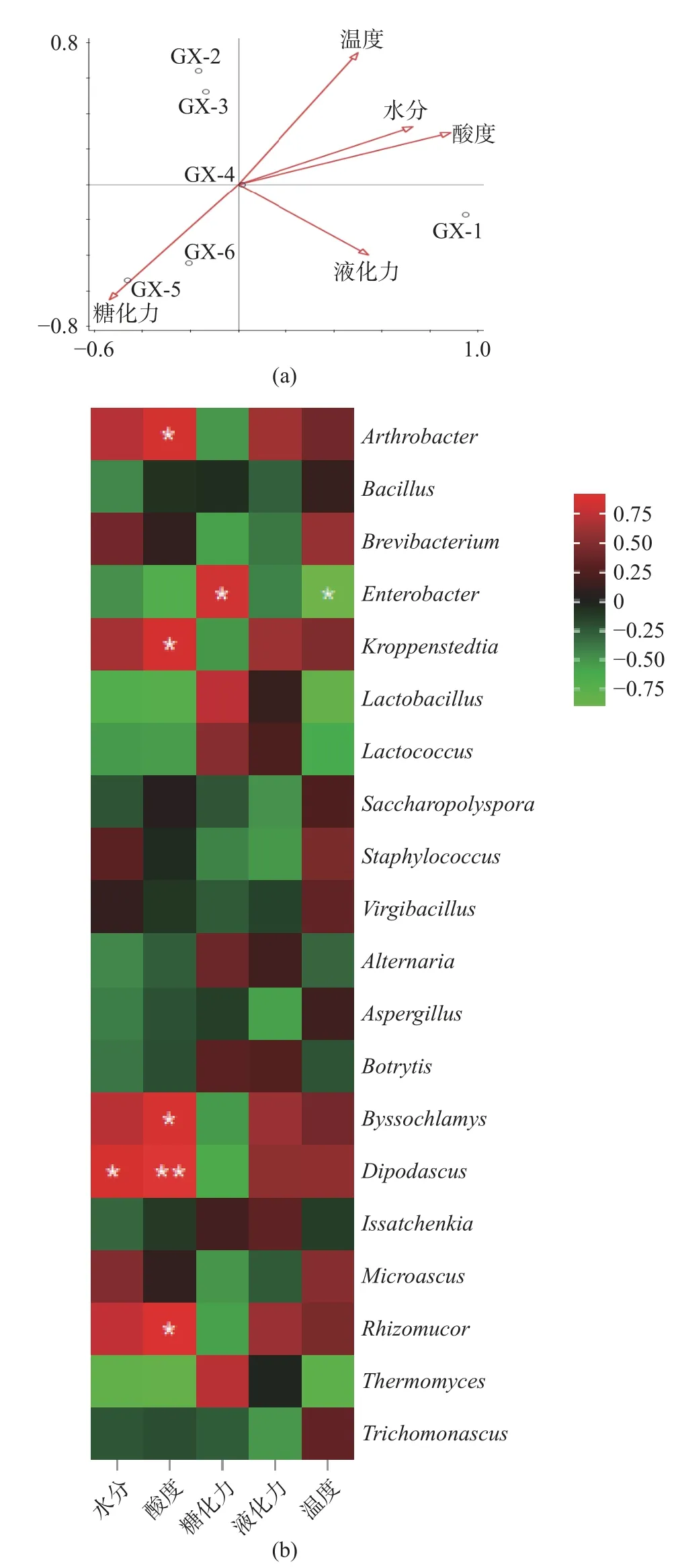
圖6 微生物群落結構與大曲理化因子相關性分析Fig.6 Correlation analysis between microble community structure and physical and chemical factors of Daqu
為揭示理化因子對優勢微生物群落的影響,選取相對豐度分別排名前10 的優勢細菌屬、優勢真菌屬(相對豐度含量至少在一個大曲樣品中大于1%的菌群結構)與儲存過程中理化因子進行相關性分析。結果如圖6(b)所示,對細菌而言,Bacillus與水分、酸度、糖化力、液化力呈負相關關系,與溫度呈正相關關系,表明了該細菌屬的生長代謝等活動受溫度的影響,在大曲儲存后期,曲倉溫度降低,其相對豐度含量降低,高酸的環境也會抑制其生長。Staphylococcus與溫度、水分呈正相關關系、與酸度、糖化力、液化力呈負相關關系;Kroppenstedtia與溫度、水分、酸度、液化力呈正相關關系,與糖化力呈負相關關系。
對真菌而言,Aspergillus、Trichomonascus等菌屬與水分、酸度、糖化力、液化力呈負相關關系,與溫度呈正相關關系;Byssochlamys、Dipodascus與溫度、水分、酸度、液化力呈正相關關系,與糖化力呈負相關關系;Thermomyces與溫度、水分、酸度呈負相關關系;Rhizomucor與水分、溫度、酸度、液化力呈正相關關系。研究表明Thermomyces屬于嗜熱真菌屬,具有促進蛋白質降解的作用,使得微生物能充分利用釀酒原料進行生長代謝[33]。Rhizomucor具有糖化發酵能力,能形成大量乳酸[34]。Aspergillus、Thermomyces、Bacillus、Issatchenkia、Lactococcus等是大曲儲存過程中的主導菌群,這些菌群的存在有利于提升大曲的發酵性能[35]。
3 結論
本文同時采用了傳統可培養方法和高通量測序技術對醬香型白酒高溫大曲在儲存(后熟)過程中微生物群落組成情況及理化因子間的相互關系進行了解析。可培養結果表明,隨著儲存時間的增加,菌落總數呈現先上升后下降趨勢;高通量測序結果表明,在儲存過程中Kroppenstedtia、Lactobacillus、Staphylococcus、Thermomyces、Trichomonascus等為主要優勢菌屬;RDA 分析結果表明大曲在儲存0 d 時,大曲的后熟過程與溫度、水分、液化力和酸度呈正相關關系,與糖化力呈負相關關系,表明該時期溫度和水分適宜,比較適合微生物生長;儲存30~60 d 時,大曲中的微生物群落結構不受水分、酸度等因素的影響,表明該時間段理化指標對微生物群落結構影響不大;儲存90~150 d 時,溫度、水分、酸度、液化力與儲存過程呈負相關,表明了大曲在存儲后期,曲倉溫度降低,曲坯水分含量低,不適宜微生物代謝活動,因此,微生物多樣性降低。目前,對于醬香型高溫大曲微生物多樣性相關研究層出不窮,在優勢微生物研究方面結論基本一致,說明了醬香型高溫大曲的微生物生態系統基本保持一致。儲存過程中微生物群落與理化因子存在復雜的作用關系,大曲在儲存過程中微生物群落的多樣性受溫度、水分和酸度等理化因子的影響,表明了儲存過程中各微生物、生化指標間相互關聯,性質上相互影響,使醬香型大曲在最佳的貯存期進行釀造,從而達到良好的產酒、產香風味。通過相關性計算可知微生物與環境因子之間的關系,目前主要是基于數學模型進行統計分析,但其內在的相互作用機制還有待進一步研究。理化因子在儲存過程中對微生物的生長繁殖及其行為具有較強的影響,進而調控微生物群落的結構及其演替,但具體的影響過程還需進一步的實驗驗證。醬香型白酒高溫大曲“后熟”時間的長短關系著大曲的品質及產酒的質量,但是目前沒有統一的標準來界定具體的儲存時間,因此,該研究為進一步闡明醬香型白酒高溫大曲后熟機理提供了理論參考。
