尼羅羅非魚Rab11-FIP5的原核表達及多克隆抗體制備
2023-08-28 04:17:32宋漫玲潘傳燕王則奮馮鵬霏張永德羅洪林
大眾科技 2023年7期
關鍵詞:研究
宋漫玲 潘傳燕 王則奮 馮鵬霏 張永德 羅洪林
尼羅羅非魚Rab11-FIP5的原核表達及多克隆抗體制備
宋漫玲1潘傳燕2王則奮1馮鵬霏2張永德2羅洪林2
(1.廣西產研院生物制造技術研究所有限公司,廣西 南寧 530201;2.廣西水產科學研究院/廣西水產遺傳育種與健康養殖重點實驗室,廣西 南寧 530021)
為研究尼羅羅非魚Rab11效應因子FIP5(Rab11-FIP5)的基因功能,試驗設計了Rab11-FIP5基因序列用于原核表達載體,構建原核表達載體pET-B2m-Rab11-FIP5,隨后誘導表達融合蛋白Rab11-FIP5,表達產物純化后免疫日本大耳兔制備Rab11-FIP5多克隆抗體,采用ELISA和Western Blot方法分析鑒定目的蛋白。結果:Western Blot檢測結果顯示有清晰目的條帶,且條帶單一,分子量約為62 KD,與預期分子量一致;ELISA檢測抗體效價為1:2 048 000,說明效價符合抗體標準,性能良好。結論:成功制備了Rab11-FIP5多克隆抗體,為開展Nt-Rab11-FIP5的定位和功能研究提供了基礎工具。
尼羅羅非魚;原核表達;多克隆抗體
引言
羅非魚是我國優勢水產養殖品種,具有蛋白含量高、生長快、環境適應性強等特點,羅非魚相關產業在沿海地區的經濟發展和帶動農民增收、脫貧致富方面做出了重要貢獻。近年羅非魚養殖頻發的病害制約著相關產業健康發展,造成的經濟損失無可計數。Rab11家族相互作用蛋白(Rab11-FIPs,也被稱為FIPs)是一個進化保守的蛋白家族,研究確認FIPs在調控真核細胞內吞循環中扮演著關鍵角色[1,2]。至今為止,魚類Rab11基因相關功能研究鮮有報道。Rab11基因被發現在斑馬魚神經系統發育過程中起著重要但截然不同的作用,該研究結果為Rab11基因在斑馬魚胚胎發育中起功能作用提供了新的證據[3]。張宇等[4]發現大黃魚全身組織均有Rab11蛋白表達,溶藻弧菌刺激后大黃魚的肝臟、腎臟和脾臟組織中Rab11蛋白水平均明顯上升,表明了Rab11基因在大黃魚的抗病免疫反應中存在重要作用。夏正龍[5]在發掘羅氏沼蝦(Macrobrachium rosenbergii)免疫相關基因時,發現了Mr Rab11基因在肝胰腺中的表達量最高,其次是肌肉和腸道;陰溝腸桿菌感染12 h后,羅氏沼蝦肝胰腺中Mr Rab11的表達量相比于對照組顯著提高,推測Mr Rab11參與羅氏沼蝦在肝胰腺中的應激免疫過程。上述文獻報道均有表明Rabll基因參與了機體對細菌和病毒免疫,且Rabll基因對細菌和病毒的免疫機制可能存在差異。
Rab11-FIP5是FIPs家族的一種效應蛋白分子,也被稱為Rip11或者Gaf-1/Gaf1b和pp75,其蛋白質的N端包含與膜磷酸肌醇結合的C2結構域,大部分位于極化的上皮細胞中,并與Rab11 GTPases發生相互作用,在細胞骨架重排、鐵攝取、胞吐等多種細胞生命活動中發揮作用。最初Rab11-FIP5是在系統性紅斑狼瘡患者的血清中發現[6],隨后發現其可調節內吞蛋白的運輸和靶向[1]。研究發現Rab11-FIP5在脂肪細胞對胰島素治療的反應中負責易位,把含有GLUT4轉運體的囊泡易位至細胞表面,同時也調節其他膜轉運調節蛋白的募集[7]。敲除siRNA的蛋白表明了Rab11-FIP5存在于外周核內體,調節內在受體的分類到緩慢的循環途徑,還發現了一種與Rip11/fip5結合的蛋白激肽II,這證明了Rab11-FIP5是引導內吞胞內蛋白進入相同循環通路所必需分子[8]。文獻研究顯示,Rab11-FIP5與兩棲類胚胎的端腦發育密切相關,Rab11-FIP5的敲除降低了端腦的細胞增殖[9]。綜上所述,Rab11-FIP5的功能研究涉及機體發育等生命活動,但魚類Rab11-FIP5的功能研究尚未見相關報道,更缺少用于后續研究的商業化抗體。為研究尼羅羅非魚Rab11-FIP5的功能特性,本研究將尼羅羅非魚Rab11-FIP5基因克隆至原核表達系統,構建原核表達載體pET-B2m-Rab11-FIP5進行表達和純化,免疫日本大耳兔獲得Rab11-FIP5多克隆抗體,為進一步研究尼羅羅非魚Rab11-FIP5的生物學功能打下基礎。
1 材料與方法
1.1 主要材料與儀器
1.1.1 樣品
尼羅羅非魚cDNA(由廣西水產遺傳育種與健康養殖重點實驗室保存)。
1.1.2 主要菌株及試劑
主要菌株:表達載體pET-B2m和表達宿主菌B21(購自武漢金開瑞生物工程有限公司)。
主要試劑:限制性內切酶、T4 DNA連接酶、蛋白marker和DNA聚合酶(Fermentas,美國),異丙基-β-D硫代半乳糖苷誘(IPTG)(MERCK,德國),弗式完全佐劑和弗氏不完全佐劑(Sigma,美國),PVDF膜(聚偏氟乙烯膜)(ThermoFisher,美國),羊抗兔-HRP抗體(Jackson,美國),親和層析柱料(GE Healthcare,美國)。
1.1.3 主要儀器
高速冷凍離心機Avanti J-26XP(購自貝克曼公司),恒溫搖床TS-211C(購自上海天呈實驗儀器制造有限公司),梯度PCR儀(購自德國Biometra公司),快速蛋白純化系統AKTA Purifier UPC 100(購自GE Healthcare)等。
1.2 試驗方法
1.2.1 蛋白結構分析和抗原預測
利用在線軟件ExPASy中的ProtScale模塊(網址:https://web.expasy.org/protscale/)對Rab11-FIP5蛋白質疏水性分析,TMHMM server v.2.0(網址:http://www.cbs.dtu.dk/ services/TMHMM-2.0/)預測蛋白序列的跨膜區間,DNAstar軟件的Protean模塊預測蛋白的親水性、抗原指數等參數。
1.2.2Rab11-FIP5基因擴增及原核表達質粒構建
根據尼羅羅非魚Rab11-FIP5基因序列(NCBI No. XP_019204508.1),使用Primer Premier 5.0設計特異性引物,在引物5' 端添加一段載體同源性序列。交由上海生工生物工程技術服務有限公司合成。引物序列為Rab11-FIP5-F:5'-TCCACTGGGTTCTCGGACTATGTTTACCCGTGATA-3';Rab11-FIP5-R:5'-TAAGGCCGCACTCGAGCACCACTTATT TACTACGAACCTGC-3'。
聚合酶鏈式反應后進行產物檢測。采用無縫克隆技術,構建pET-B2m-Rab11-FIP5原核表達系統繼續培養,之后進行產物測序鑒定。
1.2.3重組蛋白的表達和純化
將重組質粒(pET-B2m-Rab11-FIP5)轉化至大腸桿菌B21中,挑取單菌落接種于100 g·mL-1LB/Amp+培養基培養,過夜。收集過夜菌液,并超聲破碎,運用聚丙烯酰胺凝膠電泳方法分析所得上清液和沉淀。采用Ni-NTA樹脂層析柱純化重組蛋白,聚丙烯酰胺凝膠電泳方法檢測蛋白純度。
1.2.4動物免疫和重組蛋白Rab11-FIP5多抗制備及效價檢測
用純化后的融合蛋白Rab11-FIP5作為抗原免疫兩只成年日本大耳白兔,抗原注射量為500 μg/只,首次免疫將目的蛋白乳化,和弗氏完全佐劑等體積混勻后背部皮下多點注射大耳白兔進行免疫,之后每隔14天將目的蛋白與弗氏不完全佐劑等體積混勻注射大耳白兔做加強免疫,免疫次數至少4次/只,對每次免疫1周后的大耳白兔耳靜脈采血于室溫靜置2 h,或者是放置溫度為4 ℃的冰箱中過夜,收集析出血清于-20 ℃下保存;陰性對照為日本大耳白兔免疫前血清。根據說明書使用酶聯免疫吸附法(Enzyme linked immunosorbent assay,ELISA)測定分析陰性、陽性血清的效價。
1.2.5抗體特異性的Western blotting分析
將制備的蛋白樣品進行聚丙烯酰胺凝膠電泳,以免疫前血清為陰性對照,所得的抗Rab11-FIP5蛋白多克隆抗血清稀釋液(稀釋1000倍)為一抗,37 ℃孵育1小時,洗滌后再加入HRP標記的羊抗兔IgG(稀釋10 000倍)繼續孵育30分鐘,之后進行多克隆抗體特異性條帶檢測。
1.2.6抗體純化
根據張永德等[10]的方法對Nt-Rab11-FIP5多抗進行純化,用2×PBS含有0.02% NaN3以及1 m M EDTA的緩沖液收集洗脫產物,濃縮到所需體積,置于-20 ℃環境下保存,同時對樣品進行SDS-PAGE分析。
2 結果與分析
2.1 Rab11-FIP5抗原預測
對Rab11-FIP5基因進行了抗原預測(圖1)顯示,Rab11-FIP5蛋白原子數為9 261,編碼氨基酸605個,分子量為66 100.90 u,理論pI為7.25,不穩定性指數(II)為53.00,為不穩定蛋白;脂肪指數為71.85,水溶性總平均值(GRAVIY)為-0.660,該蛋白為親水性蛋白;跨膜區以及信號肽分析結果顯示,Rab11-FIP5蛋白存在跨膜區1個為第90-113氨基酸序列,該段序列有較強的疏水性,無信號肽序列;Rab11-FIP5蛋白二級結構顯示主要含有α-螺旋(Hh)、隨機線圈(Cc)和延伸鏈(Ee),三級結構顯示蛋白序列含超家族為序列17-149aa。本研究選取了150-605aa表達蛋白進行多抗免疫,該段抗原指數得分適中(圖1),可以引起較好的免疫。
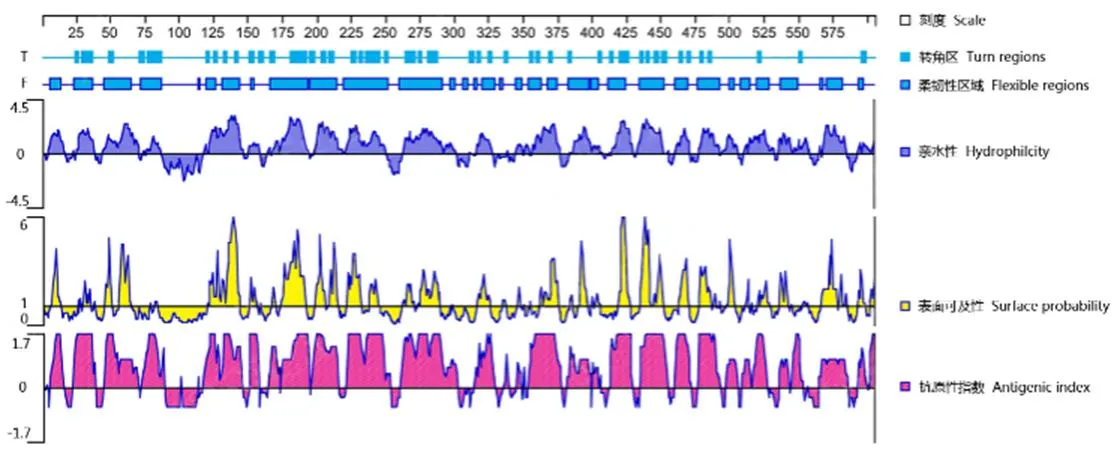
圖1 尼羅羅非魚Rab11-FIP5抗原表位分析
2.2 重組質粒Rab11-FIP5的構建與鑒定
在重組質粒pET-B2m-Rab11-FIP5目的基因片段用SalⅠ和BamHⅠ進行雙酶切和鑒定,可以清楚看到1條大小為1368 bp(圖2)的條帶,符合目的條帶預期,表明目的基因片段已成功導入表達系統Nt-Rab11-FIP5- pET-B2m構建成功。
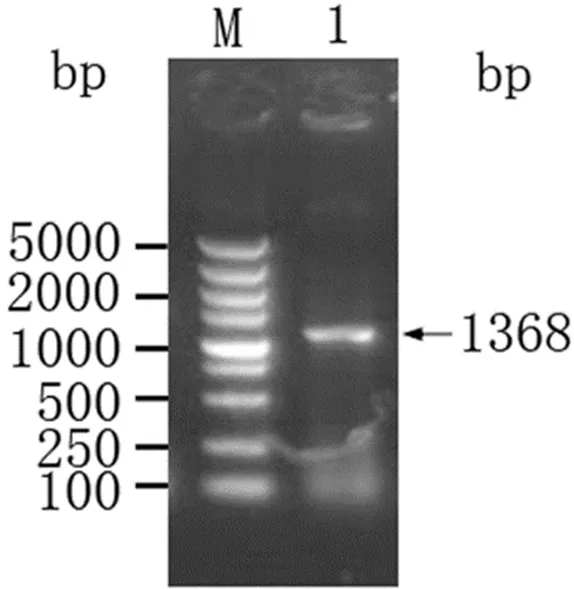
M:DL2000 DNA Marker;1:Rab11-FIP5 擴增產物
2.3 重組蛋白Rab11-FIP5的表達與純化
將已導入重組質粒pET-B2m-Rab11-FIP5的表達菌E.coli BL21置于100g·mL-1LB/Amp+培養基培養液中進行培養,加入誘導劑IPTG)誘導Rab11-FIP5蛋白表達,從圖3可以看出,上清液和沉淀泳道出現了與Rab11-FIP5蛋白分子量大小一致的蛋白,清晰可見且蛋白條帶粗細相當,表明目的蛋白在上清與沉淀均有表達且分子量表達接近,并且在0.5 mmol·L-1誘導劑、30℃下,3 h時有較高水平的表達量。同時說明pET-B2m-Rab11-FIP5重組表達載體構建成功明,表達的重組蛋白有兩種形式,分別是包涵體蛋白和可溶性蛋白存在,分子量約為62 KD。SDS-PAGE電泳對經收集、破碎、和純化后的誘導菌液進行進行分析鑒定,結果顯示經洗脫下來的重組蛋白的純度較好,雜蛋白明顯減少,可作為抗原用于后續多克隆抗體制備實驗材料(圖4)。
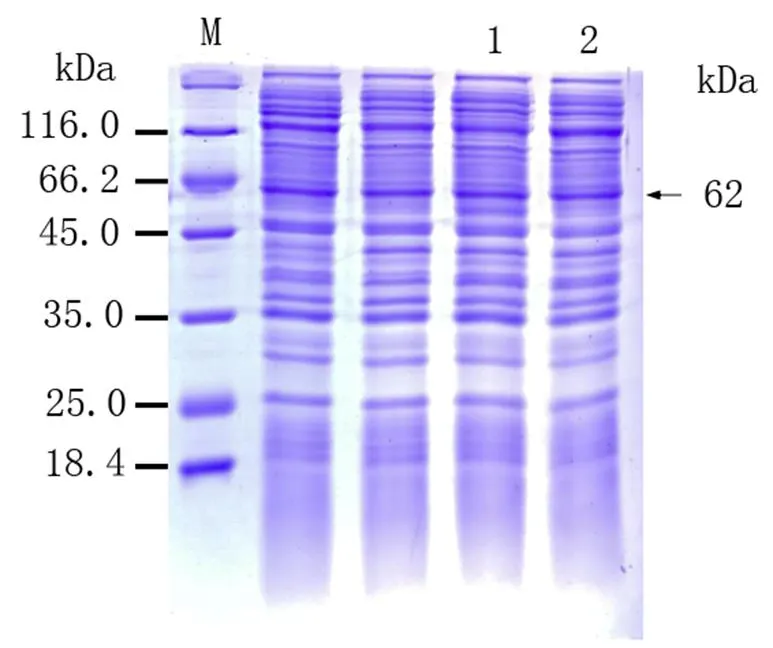
M:蛋白質Marker;1:沉淀中的目的蛋白;2:上清中的目的蛋白
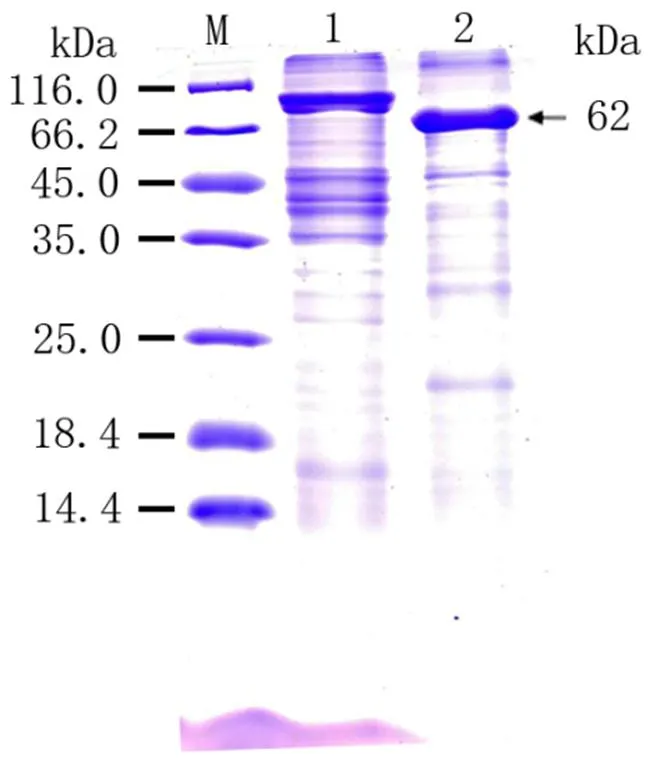
M:蛋白質Marker;1:其他蛋白;2:純化后的重組Rab11-FIP5蛋白
2.4 抗血清效價檢測
收集經至少4次免疫的兔源Rab11-FIP5抗血清,用間接酶聯免疫吸附方法檢測抗血清效價達到了1∶2 048 000(圖5),表明純化后的Rab11-FIP5重組白可以誘導日本大耳白兔產生效價高的多克隆抗體。
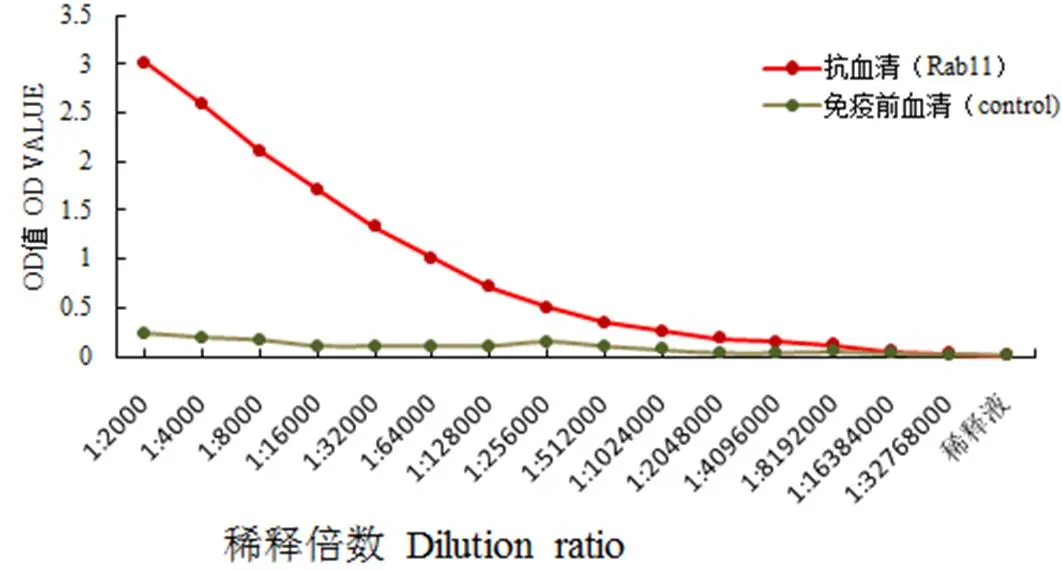
圖5 尼羅羅非魚Rab11-FIP5多肽多克隆抗體效價曲線
2.5 抗體特異性檢測結果
Western blotting結果顯示,日本大耳白兔免疫前所得抗血清為陰性對照,重組Rab11-FIP5蛋白樣品泳道檢測到蛋白分子量約62 kD蛋白(見圖6),蛋白條帶清晰,無雜帶,說明制備的抗體能夠識別并結合Rab11-FIP5蛋白,性能良好,可用于檢測尼羅羅非魚的Rab11-FIP5蛋白變化。
2.6 Rab11-FIP5多克隆抗體純化分析
將經效價分析和抗體鑒定后的抗血清進行純化,采用0.1 M檸檬酸液洗脫,用含有0.02% NaN3以及1 mM EDTA的PBS緩沖液收集。對純化后的抗體進行SDS-PAGE分析,結果表明純化效果較好,目的蛋白條帶清晰,無明顯雜帶(圖7)。
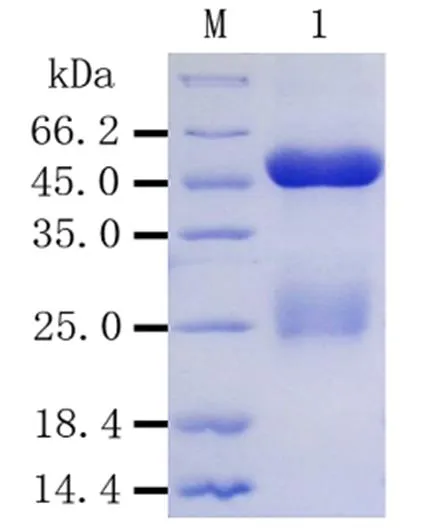
M:蛋白質Marker;1:純化的Rab11-FIP5抗體
3 討論
3.1 Rab11-FIP5的生物功能分析
Rab11-FIPs是一種高度保守的支架蛋白家族,隨著FIP功能的分子細節研究結果的公布,Rab11-FIPs被證實是細胞內多個內小體循環過程的關鍵調控者。此外,隨著研究的深入,FIP這一系列功能不斷擴大,Rab11-FIPs被證明在細胞分裂,細胞遷移,對胰島素、記憶過程、免疫和免疫的反應、炎癥反應、水的再吸收、生育、視網膜細胞發育和各種人類癌癥等生理過程中起作用[1,11-20]。由于這些細胞過程與疾病相關,因此在未來FIPs可能被確定為潛在的目標用于人類疾病的治療干預。弓形蟲在入侵哺乳動物細胞可自制膜結合室內繁殖,即寄生液泡(PV),而Ⅰ類(FIP1C、FIP2、FIP5)和Ⅱ類(FIP3、FIP4)在弓形蟲PV囊泡內存在不同含量的FIPs,說明哺乳動物Rab11參與PV蛋白形成[21]。在研究前列腺癌機制時,證據顯示Rab11-FIP5表現與胞質α6β1整聯蛋白密切聯系,α6β1是參與腫瘤轉移的重要蛋白,推測α6β1整聯蛋白通過Rab11-FIP5參與前列腺癌細胞向富含層黏連蛋白組織的遷移[22]。最新證據發現RAB11FIP5過表達調節NK細胞參與HIV-1廣泛中和抗體的產生[23]。TRIM21可介導的多聚泛素化Rab11-FIP1和Rab11-FIP5協同促進pIgA跨膜轉運,介導黏膜的免疫反應[24]。我國是羅非魚生產大國,其產業前景非常廣闊,羅非魚近年病害嚴重,本研究旨在研究Rab11-FIP5對尼羅羅非魚的生物功能關系,通過制備其多克隆抗體,進一步研究其與尼羅羅非魚的某種生物功能是否關聯。
3.2 Rab11-FIP5的生物信息學分析
生物信息學分析是一種基于核酸和蛋白質序列分析其表達的結構功能的生物信息,從而進一步了解機體的基因組學和蛋白質組學信息的方法。本研究從尼羅羅非魚cDNA文庫中篩選出Rab11-FIP5基因,設計合成尼羅羅非魚Rab11-FIP5基因,并進行了生物信息學分析。Nt-Rab11-FIP5基因具有典型的Rab結構域,屬于Rab11蛋白家族。同時,該基因在結構上高度保守,可能與不同物種間的某種相似功能有關。蛋白預測分析表明Rab11-FIP5蛋白編碼氨基酸有605個,存在跨膜結構區域1個,為第90~113氨基酸序列段,推測Rab11-FIP5蛋白活動不局限于胞內,其可能活躍于胞內外參與機體的生命活動。其他抗原性分析,該基因編碼的蛋白序列無信號肽存在,局部蛋白親水性好,含17~149氨基酸序列段為超家族,綜上研究選取了150~605氨基酸序列段,用于抗原抗體的制備,抗原表位較好。因此選擇了上述基因序列進行克隆,開展多克隆抗體制備工作。
3.3 Rab11-FIP5多克隆抗體制備分析
原核表達系統因其易培養、生長快、操作方便且能產生足量的目標蛋白等優勢,常作為外源基因融合表達。本研究通過設計并克隆出編碼尼羅羅非魚Rab11-FIP5蛋白基因序列,連接到原核表達載體pET-B2m上進行誘導表達,以獲得Rab11-FIP5重組蛋白。在誘導環節中,誘導溫度、誘導時間和誘導劑濃度是影響重組目的蛋白表達的重要因素,提前誘導菌體太少,外源蛋白產量低;誘導太遲,細菌過老,也不利于基因表達。為使得表達的外源蛋白更具備活性,產量高且質量好,本次實驗選擇確定最佳實驗條件,即37 ℃、0.5 mmoL/L IPTG、A 600 mm=0.6時進行誘導尼羅羅非魚Rab11-FIP5蛋白表達。實驗中收集并離心誘導后的細菌進行裂解,獲得重組蛋白并進行SDS-PAGE電泳,結果顯示加上清液和沉淀泳道均約有分子量62 KD出現,說明該重組蛋白以可溶性蛋白和包涵體蛋白兩種形式表達。包涵體出現的原因可能是Rab11-FIP5蛋白表達過高,或者是原核表達載體pET-B2m缺少蛋白正確折疊的影響因子,也有缺乏蛋白翻譯修飾的酶類等因素。當然,包涵體的形成并不影響抗多克隆抗體的制備,反而更有助于外源蛋白的表達與純化[22]。親和層析法是融合蛋白純化的方式之一,且其不受尿素干擾,可直接在變性條件下純化蛋白。為除去包涵體中的脂質和部分膜蛋白等雜質,對其進行洗滌和純化,本研究利用Ni-NTA樹脂層析柱純化重組蛋白,獲得了純度高達85%的重組蛋白。
免疫壞節,選擇日本大耳白兔作為免疫動物,該研究中用500 μg純化的重組蛋白和弗氏完全佐劑(1∶1)混勻后注射日本大白耳兔腹部皮下進行免疫。首次免疫后,動物機體正進行抗原刺激、識別抗原及效應B細胞增殖階段,大約7天可檢測到抗體,在10天左右達到最大值,如果第二次注入抗原時間過短,極易造成免疫抑制,因此間隔10~20天為好,以保持刺激強度,增強抗體效價。本實驗選擇間隔為兩周,共免疫5~6次,提高大耳白兔的免疫應答水平。本研究將獲得的兔抗Rab11-FIP5多克隆抗體,通過Western blot 檢測驗證,結果顯示有清晰的條帶,表明成功制備了兔抗Rab11-FIP5多克隆抗體,同時為進一步深入研究Rab11-FIP5在尼羅羅非魚中的表達及生物功能研究提供了重要工具。
4 結論
本研究據EST序列設計得到了尼羅羅非魚Rab11-FIP5 cDNA序列,該序列含有Rab家族典型保守的結構域。成功構建了尼羅羅非魚Rab11-FIP5原核表達載體,重組質粒轉入大腸桿菌B21并進行誘導表達,表達產物純化后免疫日本大耳兔制得了效價高、特異性強的多克隆抗體,為Rab11-FIP5在尼羅羅非魚的異免疫機制研究提供基礎工具,為今后其生長發育、病害防治等方面研究奠定了基礎。
[1]HORGAN C, MCCAFFREY M. The dynamic Rab11-FIPs [J]. Biochemical Society Transactions, 2009, 37(5): 1032.
[2]李琳. Rab11A效應因子FIP5在FLCN-Rab11A結合中的作用[D]. 楊凌: 西北農林科技大學,2019.
[3]ZHANGg H J, GAO Y, QIAN P P, et al. Expression analysis of Rab11 during zebrafish embryonic development.[J]. BMC Developmental Biology, 2019, 19(1): 25.
[4]張宇,韓芳,劉嵐萍,等. 大黃魚Rab11基因的克隆與表達分析[J]. 集美大學學報(自然科學版),2016,21(3): 167-174.
[5]夏正龍,黃雪娜,黃振遠,等. 羅氏沼蝦Rab11基因cDNA克隆及其組織表達分析[J]. 水生生物學報,2016,40(3): 443-450.
[6]WANG D, BUYON J P, ZHU W, et al. Defining a novel 75-kDa phosphoprotein associated with SS-A/Ro and identification of distinct human autoantibodies[J]. The Journal of Clinical Investigation, 1999, 104(9): 1265-1275.
[7]WELSH G I, LENEY S E, BETHAN L L, et al. Rip11 is a Rab11- and AS160-Rab GAP-binding protein required for insulin-stimulated glucose uptake in adipocytes[J]. Journal of Cell Science, 2007, 120(120): 4197-4208.
[8]SCHONTEICH E, WILSON G M, BURDEN J, et al. The Rip11/Rab11-FIP5 and kinesin II complex regulates endocytic protein recycling[J]. Journal of Cell Science, 2008, 121(22): 3824-3833.
[9]YOON J, GARO J, LEE M, et al. Rab11fip5 regulates telencephalon development via ephrinB1 recycling.[J]. Development, 2021, 148(3): dev196527.
[10]張永德,林勇,馮鵬霏,等. 尼羅羅非魚Lck多克隆抗體的制備及鑒定[J]. 南方農業學報,2018,49(11): 2304-2310.
[11]MUTO A, AOKI Y, WATANABE S. Mouse Rab11-FIP4 regulates proliferation and differentiation of retinal progenitors in a Rab11-independent manner[J]. Developmental Dynamics, 2010, 236(1): 214-225.
[12]HORGAN C P, OLELSY A, ZHDANOV A V, et al. Rab11-FIP3 Is Critical for the Structural Integrity of the Endosomal Recycling Compartment[J]. Traffic, 2007, 8(4): 414-430.
[13]WELSH G I, LENEY S E, LLOYD-LEWIS B, et al. Rip11 is a Rab11- and AS160-RabGAP-binding protein required for insulin-stimulated glucose uptake in adipocytes[J]. Journal of Cell Science, 2007, 120(23): 4197-4208.
[14]FAN G H, LAPIERRE L A, GOLDENRING J R, et al. Rab11-family interacting protein 2 and myosin Vb are required for CXCR2 recycling and receptor-mediated chemotaxis[J]. Molecular Biology of the Cell, 2004, 15(5): 2456-2469
[15]WANG Z, EDWARDS J G, RILEY N, et al. Myosin Vb mobilizes recycling endosomes and AMPA receptors for postsynaptic plasticity[J]. Cell, 2008, 135(3): 535-548.
[16]LINDSAY A J, MCCAFFRET M W. Purification and functional properties of Rab11-FIP3[J]. Methods in Enzymology, 2005, 403(403): 499-512.
[17]DAMIANI M T, PAVAROTTI M, LEIVA N, et al. Rab coupling protein associates with phagosomes and regulates recycling from the phagosomal compartment[J]. Traffic, 2010, 5(10): 785-797.
[18]BEHRENDS U, SCHNEIDER I, ROSSLER S, et al. Novel tumor antigens identified by autologous antibody screening of childhood medulloblastoma cDNA libraries.[J]. International Journal of Cancer Journal International Du Cancer, 2003, 106(2): 244-251.
[19]GARCIA M J, POLE J C M , CHIN S F, et al. A1 Mb minimal amplicon at 8p11-12 in breast cancer identifies new candidate oncogenes[J]. Oncogene, 2005, 24(33): 5235-5245.
[20]PATILV S. Rab coupling protein (RCP): a novel target of progesterone action in primate endometrium[J]. Journal of Molecular Endocrinology, 2005, 35(2): 357-372.
[21]HARTMAN E J, ASADY B, JULIA D R, et al.. The Rab11-family interacting proteins reveal selective interaction of mammalian recycling endosomes with the Toxoplasma parasitophorous vacuole in a Rab11-and Arf6-dependent manner[J]. Molecular Biology of the Cell, 2022, 33(5): ar34.
[22]DAS L, GARD J M C, PREKERIS R, et al. Novel regulation of integrin trafficking by Rab11-FIP5 in aggressive prostate cancer[J]. Molecular Cancer Research, 2018, 16(8): 1319-1331.
[23]BRADLEY T, PEPPA D, PEDROZA P I, et al. Rab11-FIP5 expression and altered natural killer cell function are associated with induction of HIV broadly neutralizing antibody responses[J]. Cell, 2018, 175(2): 387-399.
[24]Fan X X, ZHOU D H, ZHAO B L, et al. Rab11-FIP1 and Rab11-FIP5 regulate pIgR/pIgA transcytosis through TRIM21-mediated polyubiquitination[J]. International Journal of Molecular Sciences, 2021, 22(19): 10466-10466.
Prokaryotic Expression of Rab11-FIP5 in Nile Tilapia and Preparation of Its Polyclonal Antibody
In order to study the gene function of Rab11 effect factor FIP5 (Rab11-FIp5) in nile tilapia, the Rab11-FiP5 gene sequence was designed for prokaryotic expression vector, and the prokaryotic expression vector pET-B2m-Rab11-FIP5 was constructed, and then the fusion protein Rab11-Fip5 was induced. After purification, Rab11-FIP5 polyclonal antibody was prepared by immunizing Japanese rabbits with large eared rabbits. The target protein was analyzed by ELISA and Western Blot. Results: Western Blot analysis showed that there was a clear target band with a single band, and the molecular weight was about 62 KD, consistent with the expected molecular weight. The titer of the antibody detected by ELISA was 1:048000, indicating that the titer met the antibody standard and had good performance. Conclusions: Rab11-FIP5 polyclonal antibody was successfully prepared, which provides a basic tool for the localization and functional study of Nt-Rab11-FIP5.
nile tilapia; prokaryotic expression; purification; polyclonal antibody
S917.4
A
1008-1151(2023)07-0064-05
2023-01-26
國家自然科學基金(31372553;31760765)。
宋漫玲(1991-),女,廣西產研院生物制造技術研究所有限公司工程師,碩士,研究方向為水生動物遺傳育種。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
