自然越冬過程中鹿蹄草膜質(zhì)過氧化及滲透調(diào)節(jié)物質(zhì)含量的變化研究
2023-09-06 00:11:52王延松白海東逯錦春李彥慧
安徽農(nóng)業(yè)科學(xué) 2023年16期
王延松,白海東,逯錦春,李彥慧*
(1.河北農(nóng)業(yè)大學(xué)園林與旅游學(xué)院,河北保定 071000;2.北京市園林設(shè)計(jì)工程有限公司,北京 100012)
近年來,隨著工業(yè)化、城市化的不斷發(fā)展,人們對城市園林綠化的要求越來越高。目前,我國北方地區(qū)地被植物的應(yīng)用對建設(shè)生態(tài)園林城市發(fā)揮著重要作用。北方氣候寒冷,喬木多高大,分枝點(diǎn)高,冬季落葉,適當(dāng)?shù)剡x擇常綠的地被植物不僅可以填補(bǔ)林下的景觀空缺,還能夠嚴(yán)密地覆蓋裸露地面,增加林下景觀層次,營造復(fù)合型植物群落,健全綠地生態(tài)系統(tǒng)。低溫影響植物的生長發(fā)育,也是限制植物分布和引種的重要因素。因此,急需對植物進(jìn)行耐寒性研究并找到耐寒性強(qiáng)的植物來提高生態(tài)效益,解決地被植物的問題日益緊迫。近年來已有對茶樹(Camelliasinensis)[1]、三角梅(Bougainvilleaglabra)[2]與睡蓮(Nymphaeatetragona)[3]等耐寒性的相關(guān)報(bào)道,但對耐寒性地被植物研究相對較少,因此深入研究地被植物的耐寒性,完善評價(jià)標(biāo)準(zhǔn)體系,并結(jié)合生態(tài)效益原則,對園林植物的科學(xué)配置提供指導(dǎo)非常有必要。
鹿蹄草(PyrolacallianthaH.Andr.)是鹿蹄草科(Pyrolaceae)鹿蹄草屬(Pyrola)多年生常綠野生草本花卉[4],其花大,廣開,花瓣呈白色或粉紅色,喜冷涼陰濕環(huán)境,其生命力頑強(qiáng),具有突出的耐寒性,是四季觀賞極佳的地被植物,在城市園林綠化中有巨大的發(fā)展前景。目前,關(guān)于其抗寒性已有部分研究,如陳忠等[5]對鹿蹄草在自然越冬過程中內(nèi)源激素的研究表明,ABA/GA比值與抗寒性呈正相關(guān)。趙輝新等[6]研究發(fā)現(xiàn),鹿蹄草的解剖結(jié)構(gòu)和葉片表皮形態(tài)與其常綠抗寒的特性之間有很強(qiáng)的相關(guān)性。趙耀新[7]對鹿蹄草進(jìn)行了形態(tài)解剖、組織培養(yǎng)研究,并對其引種試驗(yàn)進(jìn)行了綜合性分析。基于前人對鹿蹄草的研究,筆者以鹿蹄草的葉片和根狀莖為試驗(yàn)材料,測定從越冬前到越冬后的地上部分葉片和地下部分根狀莖的膜質(zhì)過氧化產(chǎn)物、保護(hù)酶活性與滲透調(diào)節(jié)物質(zhì)含量,旨在明確鹿蹄草地上部分和地下部分的生理生化機(jī)制,為耐寒植物培育、引種馴化及豐富園林地被植物資源提供理論基礎(chǔ)和指導(dǎo)依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料供試材料為鹿蹄草活體植物材料,采自河北省塞罕壩機(jī)械總場陰河林場白水盤區(qū)域。該區(qū)域位于河北省圍場縣與內(nèi)蒙古赤峰市克什克騰旗和錫林郭勒盟多倫縣的接壤處(116°51′~117°39′ E,42°02′~42°36′ N),為內(nèi)蒙古高原東南緣與大興安嶺和冀北山地的交匯地帶。
1.2 試驗(yàn)方法2016年9月根據(jù)鹿蹄草的分布和長勢在該區(qū)域選定3個(gè)10 m×10 m的樣方,2016年10月至2017年5月每月中旬取樣。取樣時(shí)每樣方內(nèi)隨機(jī)選3組,每組取無病蟲害、長勢一致的鹿蹄草活體植物材料,放入冰塊保鮮的保溫箱內(nèi),運(yùn)回保定,用蒸餾水清洗,一部分用于葉色指標(biāo)測定,另一部分進(jìn)行液氮處理,待液氮處理完成,存入超低溫冰箱,用于生理指標(biāo)的測定,每個(gè)指標(biāo)3次重復(fù)。
1.3 指標(biāo)測定與方法丙二醛(MDA)含量采用巴比妥酸(TBA)法測定,超氧化物歧化酶(SOD)活性采用氮藍(lán)四唑(NBT)光化還原法測定,過氧化物酶(POD)活性采用愈創(chuàng)木酚法測定,過氧化氫酶(CAT)活性采用紫外吸收法測定[8],可溶性蛋白含量采用考馬斯亮藍(lán)G-250染色法測定,可溶性糖含量用蒽酮比色法測定,脯氨酸含量采用酸性茚三酮法測定[9]。
1.4 數(shù)據(jù)處理與分析試驗(yàn)數(shù)據(jù)用Excel軟件進(jìn)行記錄處理,使用SPSS 20.0軟件進(jìn)行顯著性分析。
2 結(jié)果與分析
2.1 自然越冬過程中鹿蹄草MDA含量動(dòng)態(tài)變化由圖1可知,葉片和根狀莖MDA含量在自然越冬過程中均呈現(xiàn)先上升后下降的變化趨勢。葉片的MDA含量在1月出現(xiàn)峰值,比10、11、12月分別升高了291.1%、208.1%、60.4%,與10、11月差異顯著(P<0.05),2、3、4、5月MDA含量逐漸降低,與1月相比,分別降低了42.8%、52.4%、52.4%、59.3%,且與3、4、5月差異顯著(P<0.05)。 根狀莖的MDA含量最大值出現(xiàn)在1月,比10、11、12月分別升高了116.8%、3.0%、1.8%。2、3、4、5月,根狀莖MDA含量逐漸降低,與1月相比,分別降低了4.5%、15.2%、43.6%、81.5%,但各月間差異均不顯著。除11月份外,其他月份葉片的MDA含量高于根狀莖。
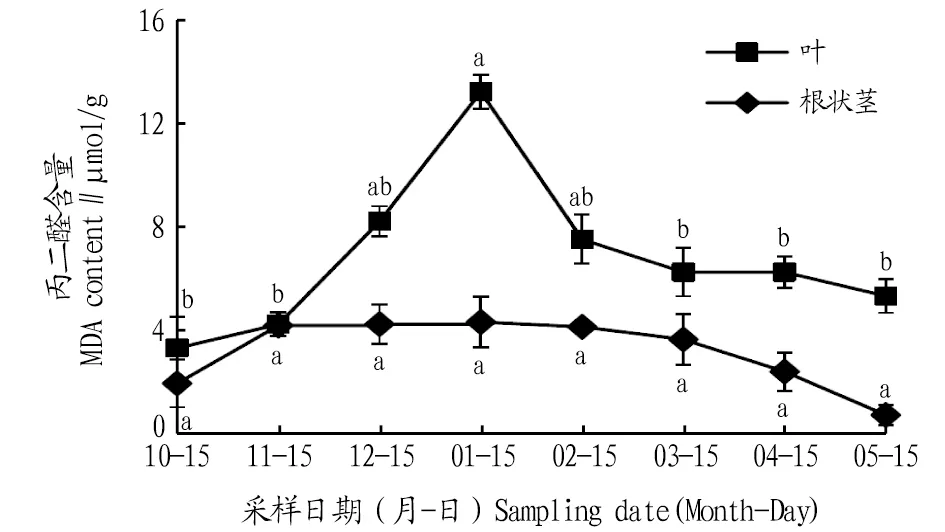
注:不同小寫字母表示同一部位不同月份間差異顯著(P<0.05)。Note:Different lowercase letters indicate significant differences between different months in the same location(P<0.05).
2.2 自然越冬過程中鹿蹄草保護(hù)酶活性動(dòng)態(tài)變化
2.2.1SOD活性動(dòng)態(tài)變化。由圖2可知,葉片和根狀莖SOD活性在自然越冬過程中均呈現(xiàn)先升高后降低的變化趨勢。葉片的SOD活性在1月出現(xiàn)峰值,比10、11、12月分別升高了45.1%、42.5%、1.8%,與10、11月差異顯著(P<0.05);進(jìn)入2、3、4、5月葉片SOD活性逐漸下降,與1月相比,分別降低了5.9%、17.1%、19.9%、37.1%,1月葉片SOD活性與3、4、5月差異顯著(P<0.05)。根狀莖的SOD活性在1月達(dá)到最大值,比10、11、12月分別升高了0.2%、1.0%、14.5%;進(jìn)入2、3、4、5月根狀莖的SOD活性逐漸降低,與1月相比,分別降低了13.4%、21.3%、29.5%、40.0%,1月根狀莖SOD活性與5月差異顯著(P<0.05)。在10、11月葉片和根狀莖的SOD活性相同,之后,葉片的SOD活性明顯高于根狀莖。
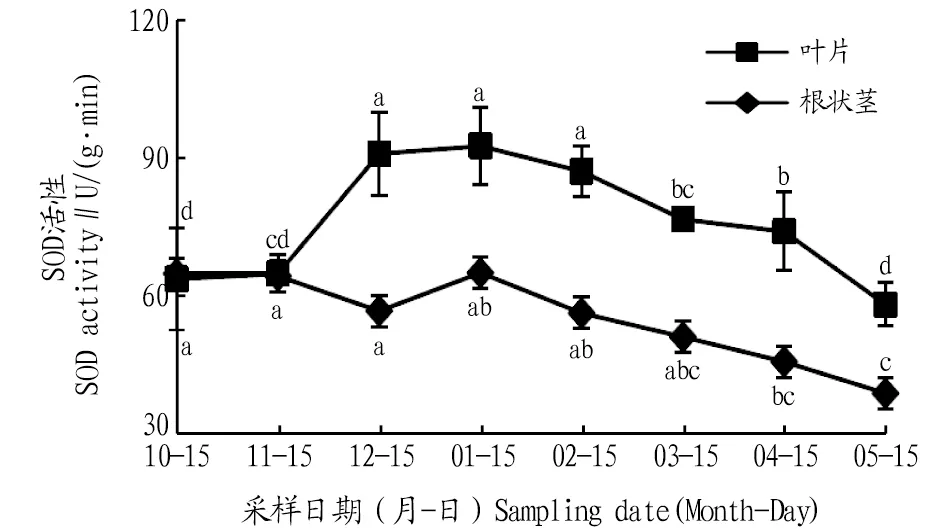
注:不同小寫字母表示同一部位不同月份間差異顯著(P<0.05)。Note:Different lowercase letters indicate significant differences between different months in the same location(P<0.05).
2.2.2POD活性動(dòng)態(tài)變化。從圖3可知,葉片POD活性在自然越冬過程中呈現(xiàn)先上升后下降的波動(dòng)變化趨勢,在1月達(dá)到最大值,比10、11、12月分別升高42.8%、24.9%、42.8%;進(jìn)入2、3、4、5月葉片POD活性降低后升高,與1月份相比,分別降低了30.0%、39.9%、49.9%、9.9%,且差異不顯著(P>0.05)。根狀莖POD活性在自然越冬過程中呈現(xiàn)先下降后上升再下降的變化趨勢,11月比10月下降了52.1%,二者間差異顯著(P<0.05),1月比11、12月分別升高了90.9%、31.2%,且與11月差異顯著(P<0.05);進(jìn)入2、3、4、5月根狀莖POD活性逐漸降低,與1月份相比,分別降低了52.3%、76.1%、76.1%、85.7%。
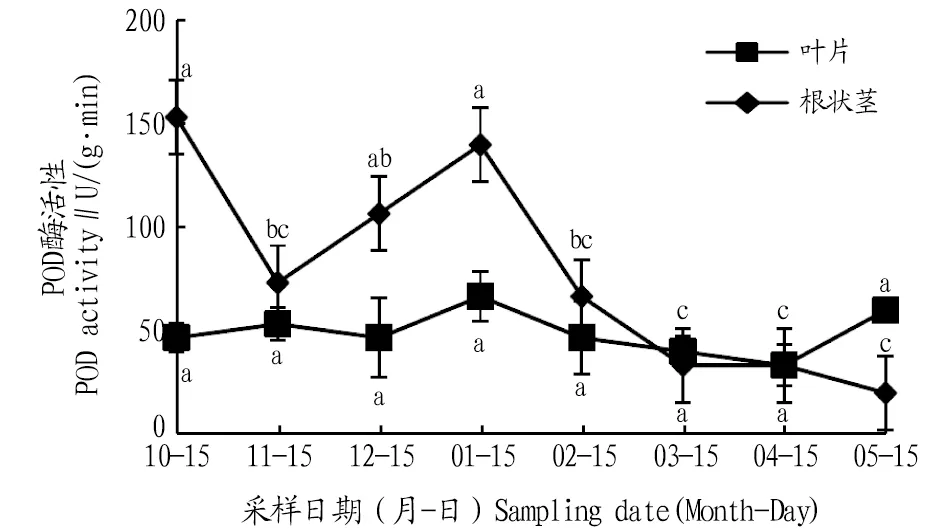
注:不同小寫字母表示同一部位不同月份間差異顯著(P<0.05)。Note:Different lowercase letters indicate significant differences between different months in the same location(P<0.05).
2.2.3CAT活性動(dòng)態(tài)變化。由圖4可知,其葉片和根狀莖CAT活性在自然越冬過程中均呈現(xiàn)先上升后下降的變化趨勢。葉片CAT活性在1月達(dá)到最大值,比10、11、12月分別上升了115.0%、61.1%、35.2%,且與10、11月差異顯著(P<0.05);進(jìn)入2、3、4、5月,葉片CAT活性逐漸降低,與1月份相比,分別降低了19.3%、19.3%、19.3%、58.4%,且1月與5月間差異顯著(P<0.05)。根狀莖CAT活性在1月達(dá)到最大值,比10、11、12月分別升高了231.1%、119.1%、88.6%,且差異顯著(P<0.05);進(jìn)入2、3、4、5月,根狀莖CAT活性逐漸下降,與1月相比,分別下降了32.8%、73.1%、85.2%、92.6%,且均差異顯著(P<0.05)。在各月份中葉片的CAT活性高于根狀莖。
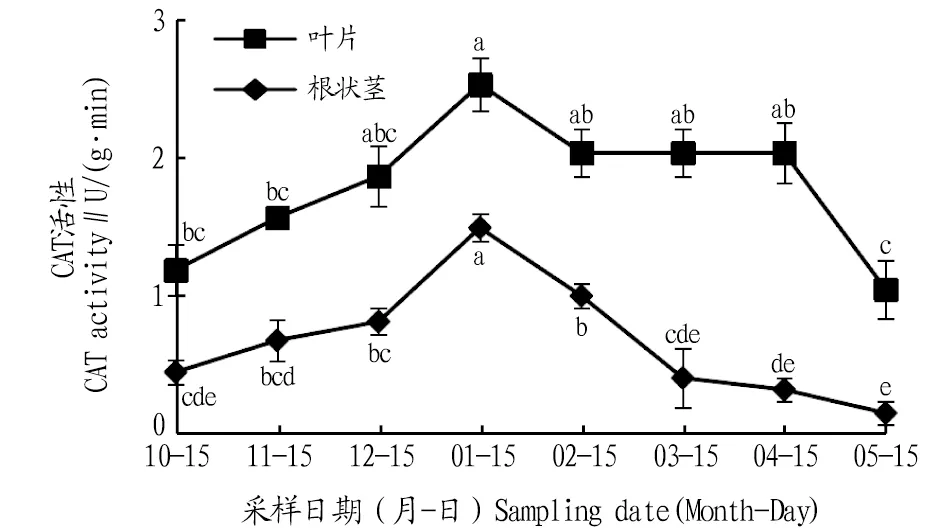
注:不同小寫字母表示同一部位不同月份間差異顯著(P<0.05)。Note:Different lowercase letters indicate significant differences between different months in the same location(P<0.05).
2.3 自然越冬過程中鹿蹄草滲透調(diào)節(jié)物質(zhì)動(dòng)態(tài)變化
2.3.1脯氨酸含量動(dòng)態(tài)變化。由圖5可知,其葉片和根狀莖脯氨酸含量在自然越冬過程中均呈先上升后下降的變化趨勢。根狀莖脯氨酸含量在1月達(dá)到最大值,比10、11、12月分別升高了46.5%、21.7%、2.5%,且與10、11月差異顯著(P<0.05),進(jìn)入2、3、4、5月,脯氨酸含量顯著下降,與1月相比,分別下降了7.9%、17.8%、24.3%、34.7%,且均差異顯著(P<0.05)。葉片脯氨酸含量在1月達(dá)到最大值,比10、11、12月分別升高了58.3%、45.4%、17.4%,且與10月差異顯著(P<0.05);進(jìn)入2、3、4、5月脯氨酸含量逐漸下降,與1月相比,分別下降了27.9%、28.3%、34.9%、37.2%,且1月與4、5月差異顯著(P<0.05)。
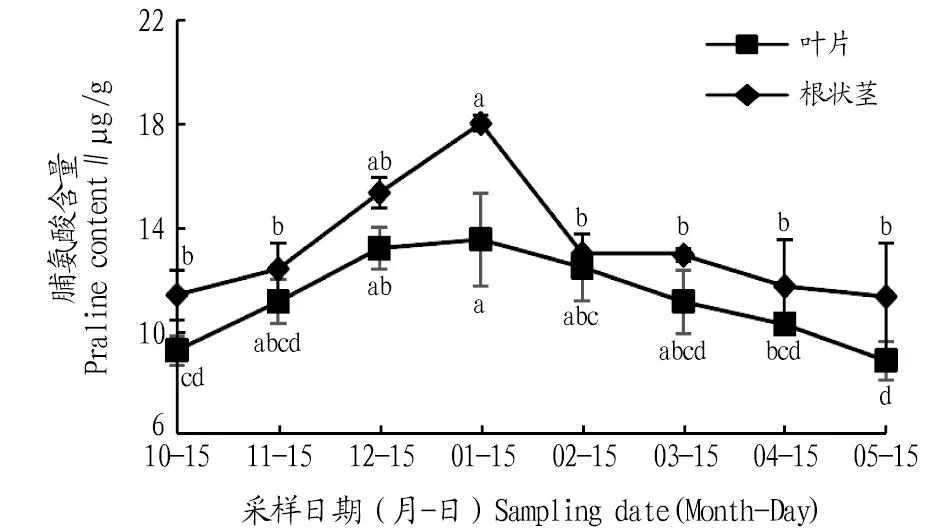
注:不同小寫字母表示同一部位不同月份間差異顯著(P<0.05)。Note:Different lowercase letters indicate significant differences between different months in the same location(P<0.05).
2.3.2可溶性蛋白含量動(dòng)態(tài)變化。由圖6可知,葉片和根狀莖可溶性蛋白含量在自然越冬過程中均呈現(xiàn)先上升后下降的變化趨勢。葉片的可溶性蛋白含量在1月達(dá)到最大值,比10、11、12月分別升高了35.0%、26.2%、0.5%,且與10、11、12月差異不顯著(P>0.05);進(jìn)入2、3、4、5月可溶性蛋白含量逐漸下降,與1月相比,分別下降了3.2%、7.8%、10%、10.3%,1月葉片可溶性蛋白含量與4、5月差異顯著(P<0.05)。根狀莖的可溶性蛋白含量在1月達(dá)到最大值,比10、11、12月分別升高15.1%、11.8%、0.4%,且與10、11、12月差異顯著(P<0.05);進(jìn)入2、3、4、5月,可溶性蛋白含量逐漸下降,與1月相比,分別下降了1.5%、2.4%、3.6%、8.6%,且1月根狀莖可溶性蛋白含量與4、5月差異顯著(P<0.05)。根狀莖的可溶性蛋白含量始終高于葉片。
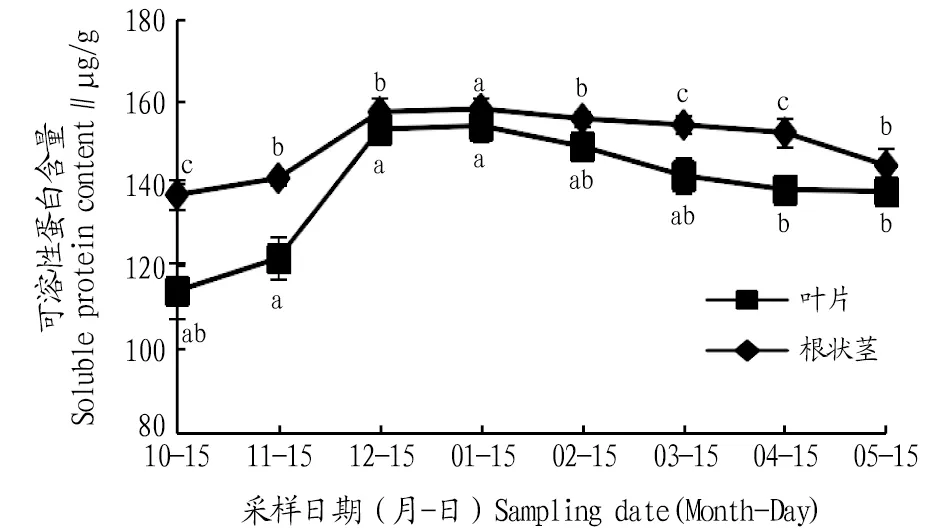
注:不同小寫字母表示同一部位不同月份間差異顯著(P<0.05)。Note:Different lowercase letters indicate significant differences between different months in the same location(P<0.05).
2.3.3可溶性糖含量動(dòng)態(tài)變化。由圖7可知,葉片和根狀莖可溶性糖含量在自然越冬過程中均呈先上升后下降的變化趨勢。葉片可溶性糖含量在1月達(dá)到最大值,比10、11、12月分別升高了48.3%、10.1%、5.1%,且與10、11、12月差異顯著(P<0.05);進(jìn)入2、3、4、5月可溶性糖含量逐漸下降,與1月相比,分別下降了11.9%、28.4%、28.6%、48.7%,且差異顯著(P<0.05)。根狀莖可溶性糖含量在1月達(dá)到最大值,比10、11、12月分別上升了40.1%、15.4%、4.8%;進(jìn)入2、3、4、5月,可溶性糖含量逐漸下降,與1月相比,分別下降了33.2%、38.7%、61.2%、65.4%,1月根狀莖可溶性糖含量與4、5月差異顯著(P<0.05)。各月葉片可溶性糖含量高于根狀莖。
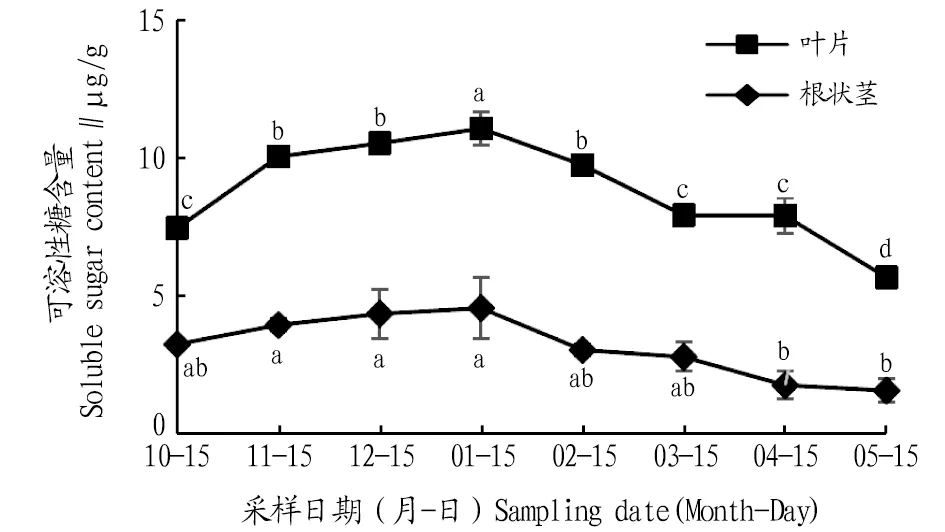
注:不同小寫字母表示同一部位不同月份間差異顯著(P<0.05)。Note:Different lowercase letters indicate significant differences between different months in the same location(P<0.05).
3 討論與結(jié)論
3.1 討論MDA是膜質(zhì)過氧化的最終分解產(chǎn)物。常溫下,MDA含量較少,但遇到逆境時(shí),MDA含量增多,表明植物受傷越嚴(yán)重[10]。對于同一植物來說,低溫時(shí)植物地上部分與地下部分的MDA表現(xiàn)不同[11]。該研究發(fā)現(xiàn),在低溫狀態(tài)下葉片和根狀莖MDA含量上升幅度較大,1月含量達(dá)到最大,進(jìn)入2、3、4、5月,當(dāng)氣溫有所回升時(shí),MDA含量又明顯下降,說明初期機(jī)體自身代謝功能突然失去平衡,膜質(zhì)過氧化加強(qiáng),溫度升高后,植物體自我調(diào)節(jié)和抵抗能力增強(qiáng),使MDA含量降低。而根狀莖MDA含量變化較小,這可能是由于葉片暴露在地上部分,細(xì)胞活動(dòng)較為旺盛,溫度等自然環(huán)境對葉片中細(xì)胞膜影響較大;而根系在地下部分受土層保護(hù),受外界環(huán)境影響較小,活動(dòng)較為穩(wěn)定。
遇到逆境條件,植物體內(nèi)產(chǎn)生的大量活性氧,會打破正常條件下活性氧的動(dòng)態(tài)平衡,從而對植物造成傷害。SOD、POD、CAT以及其他保護(hù)酶類是植物體內(nèi)重要的活性氧清除酶,逆境條件下植物體內(nèi)的SOD、POD活性均會上升[12]。有研究也已證實(shí)保護(hù)酶在植物體內(nèi)的濃度和活性高低與植物抗寒性強(qiáng)弱有密切關(guān)系,抗寒性強(qiáng)的植物具有更強(qiáng)的抗氧化能力[13]。該研究發(fā)現(xiàn),葉片和根狀莖的SOD、POD、CAT活性在1月達(dá)到最大,這主要是由于氣溫較低,植物體自身酶活性發(fā)揮最大,以抵御低溫帶來的傷害,而在試驗(yàn)期各月葉片的SOD、POD、CAT活性不同,說明葉片暴露于外部,受環(huán)境影響較大。在12月到翌年1月,植物處于抗寒保持期,葉片與根狀莖差異顯著,這是由于該時(shí)期葉片各酶活性變化較為劇烈,而根系處于地下部分較穩(wěn)定。
脯氨酸是植物體內(nèi)廣泛存在的一種滲透保護(hù)物質(zhì),植物體通過脯氨酸代謝,使體內(nèi)積累脯氨酸,增強(qiáng)對響應(yīng)和抵御低溫脅迫及生長發(fā)育的多重保護(hù)功能[14]。越來越多的學(xué)者認(rèn)為,高等植物體內(nèi)的脯氨酸代謝不僅僅局限于簡單、靜態(tài)的細(xì)胞內(nèi)的脯氨酸積累過程,而且存在不同器官間脯氨酸的長距離運(yùn)輸和流通[15]。該研究發(fā)現(xiàn),鹿蹄草根狀莖和葉片的脯氨酸含量呈先上升后下降趨勢,在氣溫最低時(shí)達(dá)到最大值,且葉片中的游離脯氨酸含量的相對增加量始終小于根狀莖,這是由于同化物在根莖中合成,運(yùn)輸時(shí)“就近分配”、優(yōu)先參與根系滲透作用有關(guān)。
可溶性蛋白含量的積累可作為滲透調(diào)節(jié)物質(zhì)和防脫水劑,從而有利于植物抗寒[16]。該研究發(fā)現(xiàn),葉片和根狀莖的可溶性蛋白含量呈先上升后下降的趨勢。在1月,可溶性蛋白的積累量達(dá)到最大,這可能是由于鹿蹄草在低溫初期體內(nèi)蛋白質(zhì)合成受到抑制,而后產(chǎn)生大量抗逆性蛋白,使得體內(nèi)滲透調(diào)節(jié)物質(zhì)增加。10、11、12月和翌年3、4月葉片和根狀莖的可溶性蛋白含量與4、5月差異顯著(P<0.05),根狀莖的可溶性蛋白含量略高于葉片,這表明在低溫傷害較輕時(shí),葉片在滲透調(diào)節(jié)方面起主導(dǎo)作用,低溫強(qiáng)度增大時(shí)根狀莖部分泌的功能性蛋白增多,滲透調(diào)節(jié)轉(zhuǎn)變?yōu)槿~部和根莖部共同作用。
可溶性糖對維持細(xì)胞膜在低溫下的正常功能有著重要作用[17]。該試驗(yàn)中,隨著低溫時(shí)間的延長,鹿蹄草可溶性糖含量呈先上升后下降的趨勢,且葉片的可溶性糖相對含量大幅高于根莖,分析可知鹿蹄草的可溶性糖對低溫的變化更敏感,且低溫時(shí)可溶性糖含量增加,提高了植物細(xì)胞的滲透調(diào)節(jié)能力,同時(shí)也降低了細(xì)胞原生質(zhì)的凝固點(diǎn),進(jìn)而增強(qiáng)了抗凍能力。
3.2 結(jié)論自然越冬過程中鹿蹄草葉片和根狀莖中MDA含量及SOD、POD、CAT活性均表現(xiàn)出先上升后下降的變化趨勢。葉片MDA含量1月比上一年10月上升291.1%。葉片和根狀莖SOD活性1月比5月分別上升了58.8%、66.7%。根狀莖POD活性1月比4月上升了320.0%。根狀莖CAT活性1月是5月的13.5倍,各月CAT活性葉片高于根狀莖,說明葉片的細(xì)胞保護(hù)酶活動(dòng)較為劇烈,根狀莖活動(dòng)較為穩(wěn)定。鹿蹄草葉片和根狀莖中脯氨酸、可溶性蛋白、可溶性糖含量均表現(xiàn)為先上升后下降的變化趨勢。葉片和根狀莖的脯氨酸含量1月比5月分別上升了53.2%、37.2%。葉片和根狀莖的可溶性蛋白含量1月比上一年10月分別上升了35.0%、15.1%,葉片可溶性糖含量1月比5月上升了94.9%。各月根狀莖的脯氨酸與可溶性蛋白含量始終高于葉片,而葉片中的可溶性糖含量則高于根狀莖,由此可知脯氨酸和可溶性蛋白的合成部位為根狀莖,可溶性糖的合成部位為葉片。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52

- 安徽農(nóng)業(yè)科學(xué)的其它文章
- 鄉(xiāng)村振興背景下涉農(nóng)學(xué)科專業(yè)平臺團(tuán)隊(duì)課程一體化建設(shè)的探索與實(shí)踐
——以凱里學(xué)院園藝專業(yè)為例 - 地方涉農(nóng)院校“新農(nóng)科”建設(shè)探索與實(shí)踐
——以遼東學(xué)院農(nóng)學(xué)院為例 - 農(nóng)村生態(tài)資源價(jià)值實(shí)現(xiàn)路徑及其法治保障研究
- 大學(xué)生掛職視角下徐州市銅山區(qū)家庭農(nóng)場創(chuàng)新發(fā)展研究
- 共同富裕背景下閑置宅基地有償退出利用機(jī)制研究
- 新安江流域水生態(tài)環(huán)境形勢分析及對策研究