蓮類胡蘿卜素裂解雙加氧酶4(NnCCD4)基因家族鑒定與功能分析
2023-09-06 07:13:24王志文楊寧寧秦莉莉曹瀧云李海鋒
核農學報 2023年10期
王志文 楊寧寧 張 晨 秦莉莉 曹瀧云 李海鋒 董 臣
(河南工業大學生物工程學院,河南 鄭州 450001)
蓮(Nelumbo nucifera)是我國重要的水生經濟作物,也是多年生草本蓮屬植物,具有廣泛的用途和價值[1]。蓮花以其獨特香氣深受人們喜愛,其花色有白色、黃色和紅色等,且與蓮類胡蘿卜素的積累有關[2]。類胡蘿卜素(carotenoid)是自然界中分布最廣的一類色素,迄今已發現近750 種類胡蘿卜素[3]。類胡蘿卜素是必要的光合色素和光保護劑,也是植物激素和植物香氣的前體物質[4]。
類胡蘿卜素裂解雙加氧酶(carotenoid cleavage dioxygenase,CCDs)是一類高度異構的多烯鏈氧化酶家族,在非血紅鐵Fe2+協助下,結合4個組氨酸發生催化反應[5]。在植物中,CCDs家族根據催化底物是否環氧化分為兩個亞家族:類胡蘿卜素裂解雙加氧酶和9-順式-環氧類胡蘿卜素雙加氧酶(nine-cis-epoxycarotenoid dioxygenase,NCED)。植物CCD 亞基因家族成員有5個,分別為CCD1、CCD2、CCD4、CCD7和CCD8;NCED亞家族成員有5 個,分別為NCED1、NCED3、NCED5、NCED6和NCED9。兩個基因家族之間的同源性和酶活性較低,底物專一性也存在差異[6-7]。
CCDs作為一種關鍵酶參與植物激素的生成,并在植物(葉、花、果實)顏色和香味形成中發揮重要作用[8]。CCD1 和CCD4 通過裂解作用,形成易于揮發的脫輔基類胡蘿卜素,產生香氣物質;CCD7 和CCD8 裂解產物是獨角金內酯形成的關鍵前體物質;NCEDs 是脫落酸(abscisic acid,ABA)形成的關鍵酶[9-10]。在植物體內同時存在多個CCD 家族基因,在不同的植物組織中發揮著各自的功能,共同調節植物類胡蘿卜素代謝[11]。
CCDs通過裂解不同底物和位點產生不同產物,從而發揮著獨特的生物學功能。目前研究發現,植物體內CCD1 和CCD4 能夠裂解多種類胡蘿卜素,產生不同的香氣物質,包括β-紫羅酮和香葉基丙酮等[12-13]。CCD1 定位于植物細胞質,CCD4 位于植物質體內[14]。考慮到質體是植物類胡蘿卜素合成的主要場所,本研究對蓮NnCCD4基因進行生物信息學分析,對NnCCD4基因表達模式進行研究,明確其酶促催化產生的香氣物質,以期為蓮屬植物的花香改良提供有效基因,為蓮種質資源開發提供依據。
1 材料與方法
1.1 材料與試劑
選取子蓮金色年華、金陵火都和太空蓮36 作為研究對象,種植于河南工業大學。收集太空蓮36 的蓮子、幼葉和成熟葉,以及金色年華(黃色)、金陵火都(紅色)和太空蓮36(白色)的花瓣。取樣完成后,立即在液氮中冷卻,后放置在-80 ℃冰箱中備用。每個材料均取3個生物學重復[15]。
細菌顏色試驗所用載體pAC-β 和pMAL-c5x均由河南工業大學生物工程學院董臣課題組保存。大腸桿菌菌株DH5α和BL21(DE3)感受態細胞購自生工生物工程(上海)股份有限公司。質粒提取試劑盒、異丙基硫代半乳糖苷(isopropyl β-D-thiogalactoside,IPTG)購自生工生物工程上海股份有限公司;植物總RNA 提取試劑盒購自北京天根生化科技有限公司;反轉錄試劑盒購自北京全式金生物科技有限公司;實時熒光定量PCR(quantitative real-time PCR,qRT-PCR)試劑盒2×ChamQ Universal SYBR qPCR Master Mix 購自南京諾唯贊生物科技股份有限公司;核酸測序及引物合成委托北京擎科生物科技有限公司進行。
1.2 儀器與設備
Mastercycler nexus X2 PCR擴增儀、Centrifuge 5418 R微量離心機、CryoCube F740超低溫冰箱,德國Eppendorf公司;7250 GC/Q-TOF 氣相色譜-質譜聯用儀,上海安捷倫科技有限公司。
1.3 試驗方法
1.3.1 蓮NnCCDs基因家族鑒定 使用NCBI(https://www. ncbi. nlm. nih. gov/)數據庫對蓮NnCCDs 編碼蛋白序列(E 值<10-6)進行在線搜索,獲得NnCCDs 蛋白序列。使用Expasy(https://www.expasy.org/)在線網站分析NnCCD4分子量和等電點等理化性質[16]。
1.3.2 蓮NnCCD4編碼蛋白序列比對及系統進化分析 在NCBI數據庫中檢索并下載蓮NnCCDs及其他高等植物CCD蛋白序列,分析蓮NnCCD4蛋白序列的相似性及其結構特征。利用DNAMAN 6.0 軟件對NnCCD4與其他物種的CCD4 進行多序列比對[17]。使用軟件MEGA 7.0 通過最大簡約法(maximum parsimony,MP)繪制高等植物CCDs家族的系統進化樹[18]。
1.3.3 蓮NnCCD4原核表達載體構建 原核表達載體構建采用一步克隆法[19]。設計特異性引物,引物兩端含有15 bp 載體序列,對目的基因進行克隆(表1)。采用pMAL-c5x 作為原核表達載體,使用限制性內切酶BamHⅠ對pMAL-c5x 載體進行酶切,將插入片段與酶切載體進行電泳檢測。膠回收進行重組反應后,轉化大腸桿菌DH5α感受態細胞,挑取陽性克隆[20]。通過質粒提取試劑盒抽提原核表達載體pMALNnCCD4a、pMAL-NnCCD4b、pMAL-NnCCD4c。

表1 引物序列Table 1 Primer sequences
1.3.4 蓮NnCCD4基因表達模式分析 通過植物總RNA 提取試劑盒提取蓮的總RNA[21]。使用反轉錄試劑盒合成cDNA,合成后的cDNA 于-20 ℃冰箱中保存[22]。設計特異性實時熒光定量引物(表1),使用DNA結合染料SYBR GreenⅠ,分析蓮NnCCD4基因在不同部位的差異性表達。反應程序:95 ℃預變性30 s;95 ℃變性10 s、55 ℃延伸30 s,40 個循環;溶解曲線95 ℃變性15 s,60 ℃延伸60 s,95 ℃變性15 s。以26 s作為內參基因,每個樣品設置3 次生物學重復,基因相對表達量用2-??Ct法計算(fold change),用Origin2017進行柱狀圖繪制及誤差分析[23]。
1.3.5 細菌顏色試驗 以pMAL-c5x 空載體作為對照,將重組質粒pMAL-NnCCD4b 與質粒pAC-β 共轉大腸桿菌BL21(DE3),37 ℃培養過夜。將共轉化的菌液使用100 mL TB 液體培養基進行培養,加入濃度為1 mmol·L-1IPTG 誘導液,誘導時間為15~20 h,產生顏色變化[24]。再使用癸酸乙酯作為內標物,將其加入到待分析的樣品混合物中,利用氣相色譜-質譜聯用技術(gas chromatography-mass spectrometry,GC-MS),并結合固相微萃取技術(solid-phase microextraction,SPME)頂空吸附上機,對蓮NnCCD4b基因共轉化菌的揮發性物質進行檢測[13,25],分析揮發性類胡蘿卜素衍生香氣物質的組分和相對含量。
用氣相色譜-質譜聯用(SPME-GC-MS)頂空固相微萃取檢測技術共轉化菌的揮發性物質,具體方法如下:
(1)設置程序:進樣口溫度為250 ℃,初始柱溫箱溫度為50 ℃保持5 min,以25 ℃·min-1升到250 ℃保持5 min 后按1∶20 的比例分流進樣;將氦氣流速保持在1 mL·min-1,不分流時間為3 min;電離方式為EI,電離能量70 eV,離子源溫度為200 ℃,DB-1701 色譜柱(30 m×0.25 mm×0.25 μm);
(2)向裝有誘導好菌液的錐形瓶中插入固相微萃取萃取頭(SPME,50/30 μm),放入60 ℃恒溫水浴鍋中30 min進行頂空吸附萃取;
(3)結束后將SPME 萃取頭插入儀器中進行解析,5 min后拔出,點擊開始鍵run;
(4)對其檢測產物進行數據分析,通過在標準品色譜圖中檢測到內標物和香氣物質的時間來確定標準品的出峰時間、出峰位置和樣品含量,從而確定樣品中內標物和香氣物質的出峰時間。
1.4 數據分析
試驗處理重復3 次,數據統計進行3 次生物學重復。采用Origin2017 軟件進行數據處理及作圖,并應用獨立樣本的t檢驗對變量進行顯著性差異分析。
2 結果與分析
2.1 蓮NnCCD4編碼蛋白的理化性質分析
從蓮基因組中成功獲得3 個CCD基因,分別命名為NnCCD4a、NnCCD4b和NnCCD4c。NnCCD4a、NnCCD4b和NnCCD4c分別含有1 914、1 812 和1 647 bp 的開放閱讀框,編碼蛋白各含有637、603和548個氨基酸。利用Expasy 在線網站分析NnCCD4 蛋白的分子量、等電點、編碼長度以及氨基酸組成等特征(表2)。

表2 蓮NnCCD4編碼蛋白的理化性質分析Table 2 Analysis of physicochemical properties of lotus NnCCD4-encoding protein
2.2 蓮NnCCD4編碼蛋白的序列比對分析
使用DNAMAN6.0 軟件對不同高等植物,如香瓜(Cucumis melo)、牽牛(Ipomoea nil)、蘋果(Malus domestica)、煙草(Nicotiana tabacum)、番茄(Solanum lycopersicum)、馬鈴薯(Solanum tuberosum)、葡萄(Vitis vinifera)、蓮的CCD4 蛋白序列進行比對(圖1)。結果表明,氨基酸序列多重比對一致性為82.98%;NnCCD4與MdCCD4 的同源性為76.48%;與VvCCD4 的同源性為74.64%;與NtaCCD4的同源性為73.08%;與SlCCD4的同源性為71.85%;與InCCD4 的同源性為72.31%;與StCCD4 的同源性為72.24%;與CmCCD4 的同源性為72.09%。說明不同物種的CCD4 蛋白序列具有較高的同源性,NnCCD4 與蘋果和葡萄之間具有較近的同源關系。
2.3 CCDs家族的系統進化分析
利用MEGA 7.0 軟件通過最大簡約法構建CCDs家族的系統進化樹(圖2)。結果表明,NnCCD4a、NnCCD4b和NnCCD4c屬于CCD 基因家族中CCD4 亞家族,不同亞基因家族之間進化關系存在差異。NnCCD4a、NnCCD4b與NnCCD4c與其他物種的CCD4亞家族同源性較低。說明NnCCD4在進化過程中保守性不強,不同種屬之間存在差異。
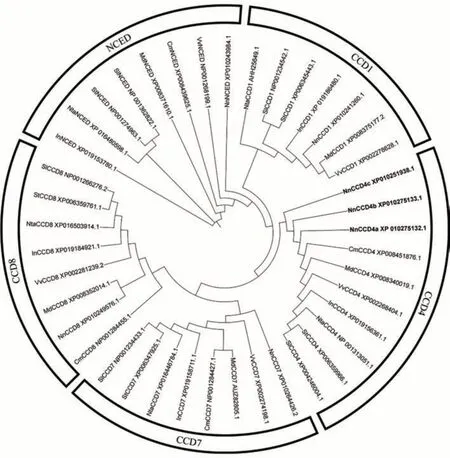
圖2 蓮NnCCD4與其他亞家族的系統進化樹Fig.2 Phylogenetic trees of NnCCD4 and other subfamilys
2.4 qRT-PCR分析蓮NnCCD4基因家族的表達模式
qRT-PCR結果表明,NnCCD4a、NnCCD4b、NnCCD4c在成熟葉片和花中的表達量普遍高于蓮子和幼葉(圖3)。其中NnCCD4a在成熟葉片中表達量較高;NnCCD4b在不同顏色的花瓣中的表達量較高,且白花表達量最高,紅花次之,黃花則相對偏低;而NnCCD4c相較于NnCCD4a、NnCCD4b在不同部位的表達量均偏低。上述結果表明,NnCCD4 家族成員在蓮不同組織器官發育中可能起到不同的作用,其中NnCCD4b在蓮花的發育過程中發揮著重要作用。
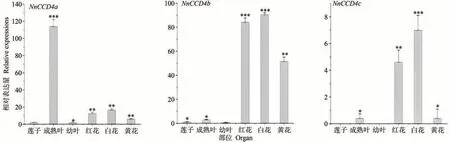
圖3 NnCCD4基因在蓮的不同部位中的表達量Fig.3 The amount of expression of the NnCCD4 gene in different organs of the lotus
2.5 NnCCD4蛋白的功能及酶切產物分析
考慮到NnCCD4b主要表達于不同顏色蓮的花瓣中,以NnCCD4b作為研究對象,分析其酶切產物。質粒pAC-β能夠在大腸桿菌中合成β-胡蘿卜素,使菌體產生黃色。如圖4-A 所示,由于pMAL-c5x 空載體不攜帶NnCCD4基因,所以共轉化菌用IPTG 誘導后,由于β-胡蘿卜素積累而呈現出深黃色。與對照相比,重組質粒pMAL-c5x-NnCCD4b 和pAC-β 共轉化菌中的顏色明顯變淺,說明NnCCD4b 有催化活性,能夠切割β-胡蘿卜素,使顏色變淺。
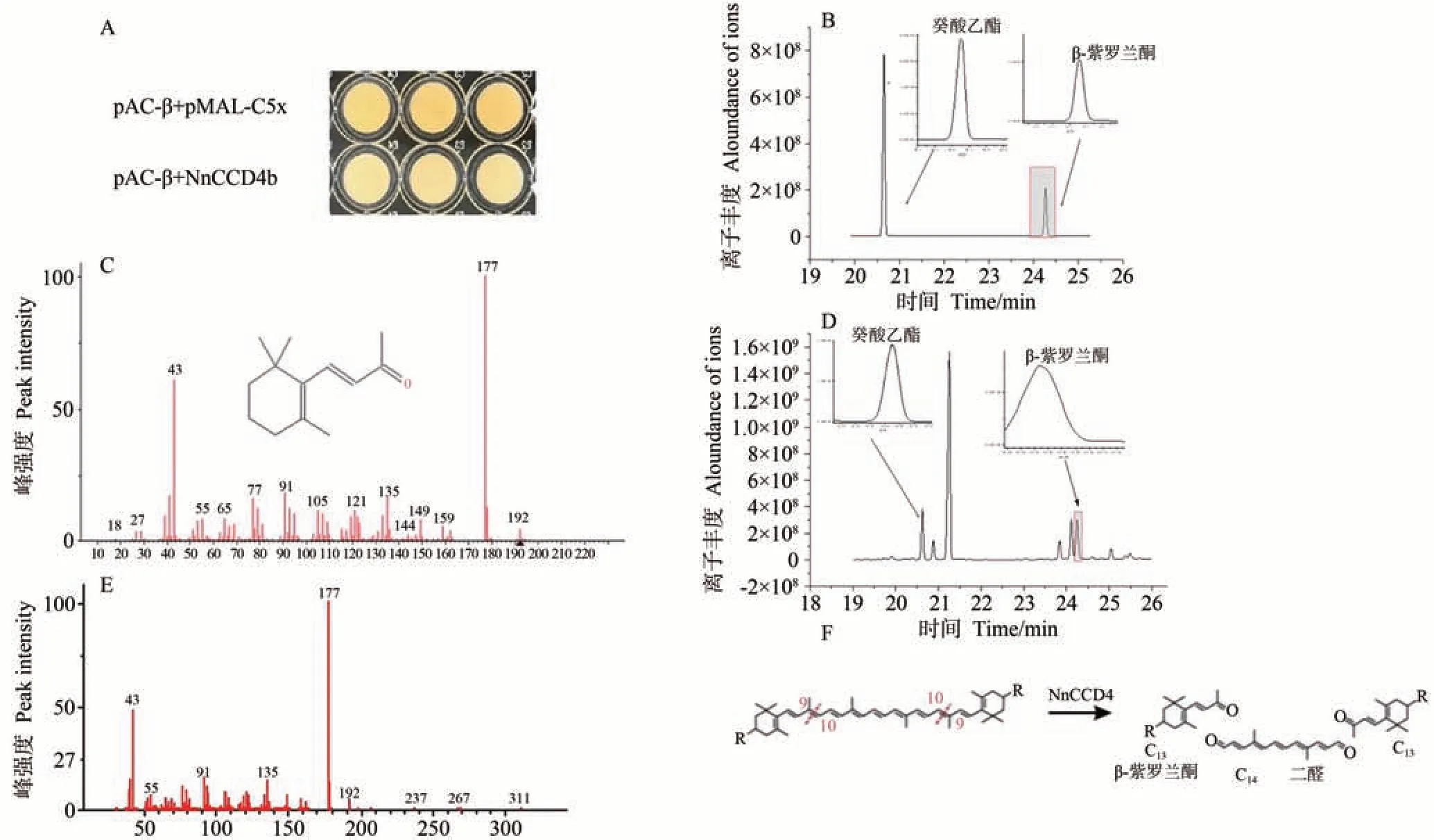
圖4 標準品與蓮NnCCD4b的類胡蘿卜素含量檢測色譜圖Fig.4 Chromatogram of carotenoid content detection of standard with NnCCD4b
為了進一步確定其裂解類胡蘿卜素產物,以pMALc5x作為對照,使用癸酸乙酯作為內標物,利用GC-MS技術,并結合SPME頂空吸附上機,對蓮NnCCD4b共轉化菌的揮發性物質進行檢測,分析揮發性類胡蘿卜素衍生香氣物質的組分和相對含量。在標準品色譜圖20.69 min 處檢測到了內標物癸酸乙酯,24.27 min 處檢測到了香氣物質β-紫羅蘭酮,從而確定了標準品的出峰時間、出峰位置和樣品含量(圖4-B)。根據總離子流色譜圖,通過檢索NIST98譜圖庫,并結合標準質譜譜圖,在重組質粒pMAL-c5x-NnCCD4b和pAC-β共轉化菌中進行檢測,分析色譜圖發現了揮發性物質β-紫羅蘭酮(圖4-D),且質譜圖(圖4-E)與標準品的質譜圖(圖4-C)一致。說明NnCCD4b 蛋白可以分解β-胡蘿卜素的9,10/9′,10′雙鍵,通過降解不同類胡蘿卜素底物的不同雙鍵,生成13 個碳原子的β-紫羅蘭酮(圖4-F)。
3 討論
本研究對NnCCD4基因家族保守基序、系統發育和表達模式進行了分析,通過同源克隆從蓮基因組中獲得了3 個NnCCD4 基因家族成員:NnCCD4a、NnCCD4b和NnCCD4c,結果表明,NnCCD4 基因家族的蛋白相對分子質量大小在61.49~70.14 kDa,pI值在5.55~6.52,表明NnCCD4 家族成員間在蛋白質大小、pI 值等特征參數上差異不大。克隆得到的3 個NnCCD4 基因家族成員與其他植物中已報道的CCD4基因相似[26]。NCBI比對結果表明,NnCCD4基因具有CCDs 家族基因相似結構特征的保守結構域。對蛋白序列進行研究發現,NnCCD4 與其他物種CCD4 家族成員的蛋白序列有較高的同源性。此外,NnCCD4c與NnCCD4a、NnCCD4b的親緣關系較遠,說明不同亞基因家族之間親緣關系存在差異。通過分析CCDs 的家族進化樹,發現NnCCD4在進化過程中保守性不強,不同種屬之間存在差異,這與岳遠征等[27]和王贊等[28]的研究結果一致。
植物的生長發育離不開CCD亞家族各成員的表達。Rubio等[29]證明番紅花(Crocus sativus)CsCCD4b僅在柱頭組織中表達;王昊等[30]證明芹菜(Apium graveolens)AgCCD4在葉片中高表達,葉柄和根中的表達量低,說明CCD 亞家族基因在植物不同組織和器官中具有表達特異性。本研究結果顯示,3 個NnCCD4基因在蓮子、葉片和花中的表達量都存在明顯差異:NnCCD4c表達量最低;對比葉片的2 個生長時期發現,在成熟葉片中NnCCD4a表達量明顯高于幼葉中的表達量。在3種不同顏色的花中,NnCCD4b表達量遠高于NnCCD4a和NnCCD4c,且白花表達量最高,紅花次之,黃花則相對偏低。
CCD 基因家族成員是一類具有RPE65 保守結構域的基因家族,大多存在于植物中,如葡萄基因組中鑒定出19 個CCD[31]、水稻中鑒定出11個CCD[32]、擬南芥和番茄中均鑒定到9 個CCD[6,33]。CCD 亞家族各成員編碼的酶在類胡蘿卜素的代謝過程中發揮了重要作用,目前在番紅花、桂花和矮牽牛中均已證實CCD1 和CCD4可參與類胡蘿卜素的降解,導致類胡蘿卜素含量發生變化,裂解產生獨特的香氣物質。Baldermann等[34]證明桂花OfCCD1可裂解β-胡蘿卜素產生β-紫羅蘭酮、假紫羅蘭酮以及香葉基丙酮。Simkin等[35]證明矮牽牛花中PhCCD1的表達量增加會使β-紫羅蘭酮的含量相應升高。Rubio等[29]證明番紅花CsCCD4柱頭發育期間會裂解出β-胡蘿卜素,產生β-紫羅蘭酮。Song等[36]證明在轉基因水稻中過表達擬南芥AtCCD4 會使水稻中β-胡蘿卜素降低74%,且β-紫羅蘭酮的含量是非轉基因水稻的2 倍。本研究通過GC-MS 和SPME 頂空吸附上機對蓮NnCCD4b共轉化菌的揮發性物質進行檢測,結果表明NnCCD4 可以催化β-胡蘿卜素氧化裂解,產生香氣物質β-紫羅蘭酮。這與Baldermann 等[34]和Simkin 等[35]的研究結果一致。本研究表明NnCCD4很可能是蓮代謝途徑里的一個關鍵基因,這為今后植物品質改良提供了一定的理論依據,為后續進一步研究蓮NnCCD4基因家族功能特性提供了理論基礎。
4 結論
本研究以NnCCD4 為研究對象,通過生物信息學手段對其氨基酸序列進行分析,發現NnCCD4 與其他物種家族成員同源性較高,且NnCCD4在不同亞基因家族之間親緣關系存在差異;采用qRT-PCR 對不同生長時期NnCCD4基因的表達量進行分析,證明了蓮不同組織部位中不同基因之間存在表達差異,通過構建原核表達載體,進一步研究NnCCD4 蛋白的酶活性以及酶切產物,從分子水平上揭示了NnCCD4 降解類胡蘿卜素產生香氣物質對蓮品質的影響。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
