褪黑素調控水楊酸和乙烯代謝維持沙果貯藏品質
2023-09-07 00:43:44袁瑞敏王佳傲李湘鵬李建揮
核農學報 2023年9期
關鍵詞:生物
袁瑞敏 彭 靜 王佳傲 李湘鵬 李建揮 閔 婷 林 瓊,*
(1中國農業科學院農產品加工研究所,北京 100193;2湖南省植物園,湖南 長沙 410116;3武漢輕工大學食品科學與工程學院, 湖北 武漢 430023)
褪黑素(melatonin, MT)是存在于動植物體內、具有較強生物活性的多功能胺類分子[1-2],于1958 年首次在牛松果腺中被發現[3],參與植物成熟衰老等一系列生命活動的調控[4-5],可提高植物對生物脅迫的抵抗性及對冷害、高鹽等非生物脅迫的耐受性[6-7]。研究表明,MT 可通過自身的清除性,誘導果實中超氧化物歧化酶(superoxide dismutase, SOD)、谷胱甘肽還原酶(glutathione reductase, GR)及抗壞血酸過氧化物酶(ascorbate peroxidase, APX)等抗氧化酶和酚類物質、還原型谷胱甘肽等非酶抗氧化劑來消除柑橘[8]、番茄[9]、石榴[10]果實及大白菜[11]中過多的活性氧,保護其免受氧化損傷,維持活性氧代謝,延緩果蔬衰老以保持較好的品質。Wei等[12]研究表明,MT處理能誘導香蕉果實中MT、水楊酸(salicylic acid, SA)和乙烯含量的積累,提高香蕉對枯萎病的抗性。采前MT 噴施處理番茄果實能顯著提高其對灰葡萄孢菌侵染的抵抗能力,有效提高番茄果實的貯藏品質[13-14]。此外,MT 在延緩蘋果[15]、獼猴桃[16]、梨[17]果實及黃瓜[18]的后熟衰老進程中效果顯著,能抑制果實組織中呼吸速率及乙烯合成,降低果蔬失水和軟化的發生,更好地保持果實品質,但也能顯著提高杏[19]果實中的乙烯積累,加快果實成熟進程,可能是由于不同濃度MT 在不同果蔬中的表達效果有所差異。
沙果(MalusasiaticaNakai)為薔薇科仁果亞科蘋果屬植物的果實,正名花紅果[20]。其果皮鮮紅,果肉為淡黃色,富含多酚、有機酸、維生素、膳食纖維等成分,具有生津、止咳和抗氧化等功效[21]。乙烯信號對沙果果實這一典型的躍變型果實的成熟具有調控作用。乙烯是由S-腺苷甲硫氨酸通過1-氨基環丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)合成酶(ACC synthase, ACS)和ACC氧化酶(ACC oxidase,ACO)合成[22]。研究表明,MT 處理葡萄果實能誘導MYACS1表達,促進果實中乙烯的生物合成[23]。但Onik等[15]發現MT處理富士蘋果能有效降低MdACO1和MdACS1的轉錄豐度,使其在貯藏期間的乙烯生成量減少。這與Yuan 等[24]的研究結果類似。水楊酸(salicylic acid, SA)是一種酚類物質,參與植物生長、發育和成熟衰老的調節[25-26]。前后關聯分支酸是MT與SA在生物合成代謝途徑中共同的起始物質,是合成苯丙氨酸、色氨酸的原料[27]。植物中的MT 由色氨酸通過鄰氨基苯甲酸或吲哚途徑合成[6],SA 則由苯丙氨酸解氨酶(phenylalanine ammonia-lyase, PAL)或異分支酸合酶(isochorismate synthase, ICS)途徑合成[28]。SA處理可以抑制蘋果[24]、獼猴桃[29]、梨[30]和枸杞[22]果實的采后成熟,與MT 在維持果蔬成熟過程中的作用類似,兩種植物激素具有較高的相似性。張靜等[27]對薺菜進行SA處理,發現芽苗組織能通過上調BjTDC1/2和BjT5H1/2的表達來富集MT;同時MT 處理冷藏桃[31]果實顯著提高了果肉中的SA含量。
鑒于此,本研究以沙果為試驗材料,從SA 和乙烯生物合成代謝兩方面研究MT 處理對沙果在常溫貯藏期間生理生化指標變化的影響,探討褪黑素處理后沙果品質及成熟的調控機制,旨在為揭示褪黑素調控沙果果實的機理奠定理論依據。
1 材料與方法
1.1 材料與試劑
新鮮沙果(MalusasiaticaNakai),于2021 年9 月采摘自內蒙古科右前旗,在24 h 內通過冷鏈運輸至中國農業科學院農產品加工研究所實驗室,放入4 ℃預冷箱預冷6 h 去除田間熱,選取成熟度一致(八成熟)、大小均勻、果柄新鮮完整、無病蟲害、無機械損傷、無腐爛變質的沙果果實作為試驗材料。
褪黑素,Sigma-Aldrich 上海有限公司;三氯甲烷、異戊醇,上海麥克林生化科技有限公司;焦炭酸二乙酯(diethy pyrocarbonate, DEPC)、β-巰基乙醇,生工生物工程(上海)股份有限公司;三羥甲基氨基甲烷[tris(hydroxymethyl)aminomethane, Tris]、聚乙烯吡咯烷酮(polyvinylpyrrolidone, PvP)、十二烷基硫酸鈉(sodium laurylsulfonate, SDS)、十六烷基三甲基溴化銨(hexadecyl trimethyl ammonium bromide, CTAB)、乙二胺四乙酸(ethylene diamine tetraacetic acid, EDTA)、氯化鋰、亞精胺、氯化鈉,國藥集團化學試劑北京有限公司。試劑均為分析純。植物水楊酸酶聯免疫分析試劑盒,北京威萊博生物技術有限公司;全式金試劑盒,北京全式金生物技術有限公司。
1.2 儀器與設備
GY-4果實硬度計,浙江托普云農科技股份有限公司;F-950 型便攜式C2H4/CO2/O2分析儀,美國Felix 公司;YP502N 電子天平,上海精密科學儀器有限公司;BIORAD680型全自動酶標儀,美國Biorad公司;Nanodrop 2000 分光光度儀,北京龍躍生物科技發展有限公司;ABI7500實時定量PCR儀,美國Perkin Elmer公司。
1.3 試驗方法
1.3.1 處理方法 試驗前將預冷的沙果用清水沖洗浮塵,然后立即放入0.1% NaClO(v/v)水溶液浸泡5 min進行減菌處理,接著撈出控干水分,將沙果平均分成兩組進行處理,每組150 個果實,分別為褪黑素(MT)處理組和對照組(CK)。MT 組沙果浸沒到1.0 mmol·L-1MT 溶液中處理15 min;CK 組除溶液為蒸餾水,其他條件與MT 組相同。處理后將沙果果實撈出,輕輕甩掉表面的溶液,控干水分后置于(20±1) ℃的恒溫箱中貯藏28 d。在貯藏3、7、14、21、28 d取樣點隨機取不同處理的沙果果實進行測定,每個指標測定時在不同處理組中進行3次重復取樣。
1.3.2 品質指標的測定 參考苑智華[31]的方法并稍作修改,隨機挑選9 個沙果并削去其赤道上間隔等距的3 個位置約1 mm 的果皮。將8 mm 圓柱形探針垂直于沙果的赤道表面,以2 mm·s-1的速度向下用力均勻地壓入沙果的果肉中,穿透深度為10 mm,記錄硬度計顯示數據。在進行下一個位置的測定前需要將硬度計進行調零操作。結果用牛頓(N)表示。
沙果果實的失重率采用稱重法[32]測定。在試驗期間,MT 與CK 組中各9 個沙果果實被反復評估。計算見式(1):
式中,W為沙果果實重量損失;m0為初始沙果果實重量;mn為不同取樣日稱量的沙果果實重量。
1.3.3 呼吸速率和乙烯釋放量的測定 參考Li等[33]的方法并稍作修改。將兩處理組的沙果果實密封在兩個密閉性較好的1.5 L 帶蓋塑料容器中(每個盒子3個沙果)。蓋上蓋子則開始計時,在20 ℃下密閉處理2 h。然后通過C2H4/CO2/O2分析儀分析塑料容器中的上層氣體樣本,得到CO2和乙烯(C2H4)濃度。每個處理重復取樣3次,每個重復測定呼吸速率和乙烯釋放量3次。結果分別以mL·kg-1·h-1和μg·kg-1·h-1表示。
1.3.4 SA 含量的測定 使用植物水楊酸酶聯免疫分析試劑盒根據雙抗體夾心法測定樣品中SA含量。稱取0.2 g粉末鮮樣,加入磷酸緩沖鹽溶液(phosphate buffer saline, PBS)緩沖液1.8 mL 渦旋混勻后,在4 ℃條件下5 000× g離心15 min得到SA 粗提液,每個樣品重復測定3次。按照說明書進行后續試驗。
1.3.5 基因表達水平的檢測 使用Primer Premier 5.0 軟件對沙果果實相關基因進行qPCR 特異引物的設計,引物的合成由北京擎科生物科技有限公司完成,引物序列見表1。使用傳統的十六烷基三甲基溴化銨(CTAB)法從沙果果實中提取RNA。采用全式金試劑盒進行RNA 逆轉錄,合成cDNA。參考Yuan 等[24]的方法構建實時熒光定量聚合酶鏈反應(quantitative real time polymerase chain reaction, qRT-PCR)程序:Step 1:94 ℃ 孵育30 s;Step 2:94 ℃ 孵育5 s;Step 3:60 ℃ 孵育30 s;Step 4:跳回Step 2,40次循環;Step 5:95 ℃ 孵育15 s;Step 6:60 ℃ 采集34 s,通過7500 Fast Real-Time PCR System檢測各代謝途徑關鍵基因的表達水平。

表1 qRT-PCR分析的引物序列Table 1 Primer sequences used for qRT-PCR analysis
1.4 數據分析
使用SPSS 25.0軟件進行數據分析,并表示為3 個重復的平均值±標準誤差,各組間差異通過單因素方差分析(ANOVA)進行評估,P<0.05 表示差異顯著。使用OriginPro 9.0軟件繪制圖表。
2 結果與分析
2.1 MT處理對沙果果實水楊酸含量的影響
由圖1 可知,貯藏過程中,MT 處理組和CK 組沙果果實中SA含量表現為先升高后降低,均在貯藏14 d達到峰值,分別為0.84和0.71 mg·kg-1,MT較CK顯著增長了19.18%(P<0.05)。何雪瑩等[34]研究表明,不同品種蘋果果實在成熟過程中的SA含量基本呈現先升后降的趨勢。貯藏7 d后,MT處理組SA含量始終顯著高于CK;貯藏28 d,MT處理組SA含量是CK的1.32倍。上述結果表明,外源MT 處理沙果果實能夠使果肉中SA 含量累積,這與苑智華[31]在冷藏桃果實中的研究結果類似。

圖1 MT處理對沙果果實水楊酸含量的影響Fig.1 Effects of MT treatment on SA content of shaguo
2.2 MT處理對沙果果實乙烯釋放量的影響
呼吸躍變型果實成熟最典型的特征是果實采收后仍然需要等待后熟才能完全成熟,后熟的過程伴隨著呼吸作用驟然升高以及大量乙烯的生成[35]。沙果作為典型的呼吸躍變型水果,其生長發育、成熟衰老和貯藏時間的長短主要依賴于乙烯的生成和催化[36]。由圖2 可知,貯藏期間,沙果果實乙烯釋放量呈現先急速上升至最高值,后隨著果實成熟而緩慢下降的變化。CK 組沙果果實的乙烯高峰出現在貯藏14 d,達到70.34 μg·kg-1·h-1。MT 處理組果實的乙烯高峰也出現在貯藏14 d,但僅為65.17 μg·kg-1·h-1,較CK 顯著降低了7.36%。同時,在整個貯藏期間其乙烯釋放量始終處于較低水平。
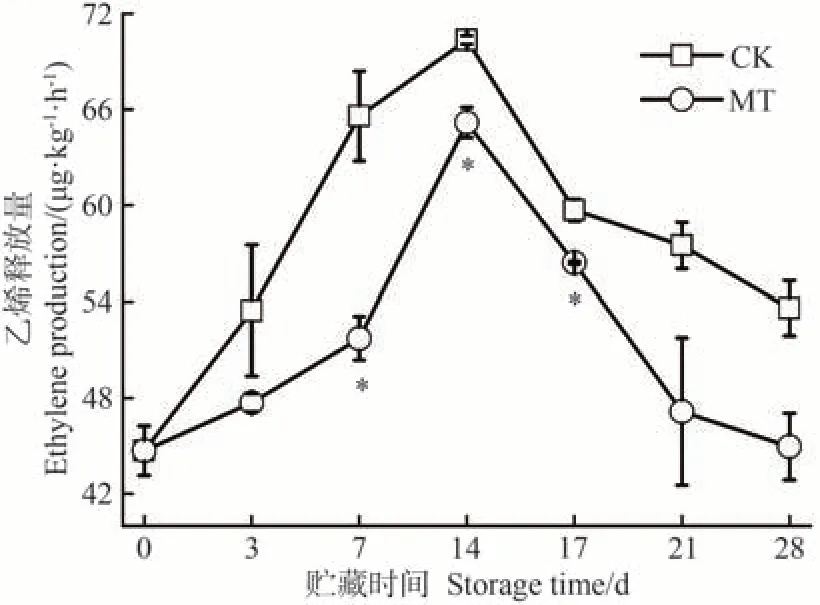
圖2 MT處理對沙果果實乙烯釋放量的影響Fig.2 Effects of MT treatment on ethylene production of shaguo
2.3 MT處理對沙果果實水楊酸合成關鍵基因的影響
在SA的兩條生物合成途徑中,ICS及PAL是其積累的關鍵酶。如圖3所示,在貯藏期間,MdPAL和MdICS2在沙果果實中表達水平的變化趨勢與SA含量一致。貯藏3、7和14 d時,MT處理組MdPAL相對表達量相較于CK組顯著增加,其中貯藏14 d時,MT處理組MdPAL相對表達量顯著增加38.29%。MT組MdICS2表達水平在貯藏3 d時較CK組顯著增加了16.96%(P<0.05),其表達量在貯藏14 d達到峰值,可見MT處理誘導了水楊酸生物合成關鍵基因的上調,這與SA含量顯著提高相一致(圖1)。外源MT在貯藏過程中能夠通過促進MdPAL和MdICS2的基因表達量,增加沙果果實中SA的生物合成。研究表明,SA及其代謝衍生物可以抑制乙烯前體的合成,最終降低乙烯產量[24,37]。因此推測MT可誘導沙果果實中SA合成,SA 與MT 均能抑制采后果實的乙烯生物合成途徑[15,24],可能是由于MT和SA單獨或共同作用抑制了乙烯生物合成和乙烯釋放,最終達到延緩果實成熟的目的。
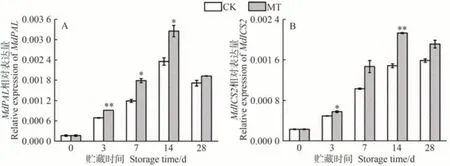
圖3 MT處理對沙果果實水楊酸合成關鍵基因MdPAL(A)和 MdICS2(B)相對表達量的影響Fig.3 Effects of MT treatment on the relative expression of MdPAL (A) and MdICS2 (B) in shaguo
2.4 MT處理對沙果果實乙烯合成關鍵基因的影響
乙烯生物合成的關鍵酶主要為ACS 和ACO。在采摘后的蘋果果實中,沉默MdACS1[38]和MdACO1[39]會導致果實無法發生呼吸躍變而出現無法成熟的表型,因此ACS1和ACO1被認為是調控蘋果果實乙烯生物合成中重要的基因。如圖4 所示,沙果果實中MdACS1和MdACO1的基因表達量在貯藏前期迅速上升,與乙烯釋放量變化趨勢一致(圖2)。MdACS1和MdACO1在MT 處理組的沙果果實成熟過程中表現出相似的表達模式,在整個貯藏過程中均受到了顯著抑制,貯藏28 d 時,兩基因的相對表達水平相較于CK組分別顯著降低了15.39%和15.31%。上述結果表明,外源MT 在貯藏過程中下調MdACS1和MdACO1的表達水平,從而抑制了沙果果實乙烯的釋放,緩解了果實成熟。
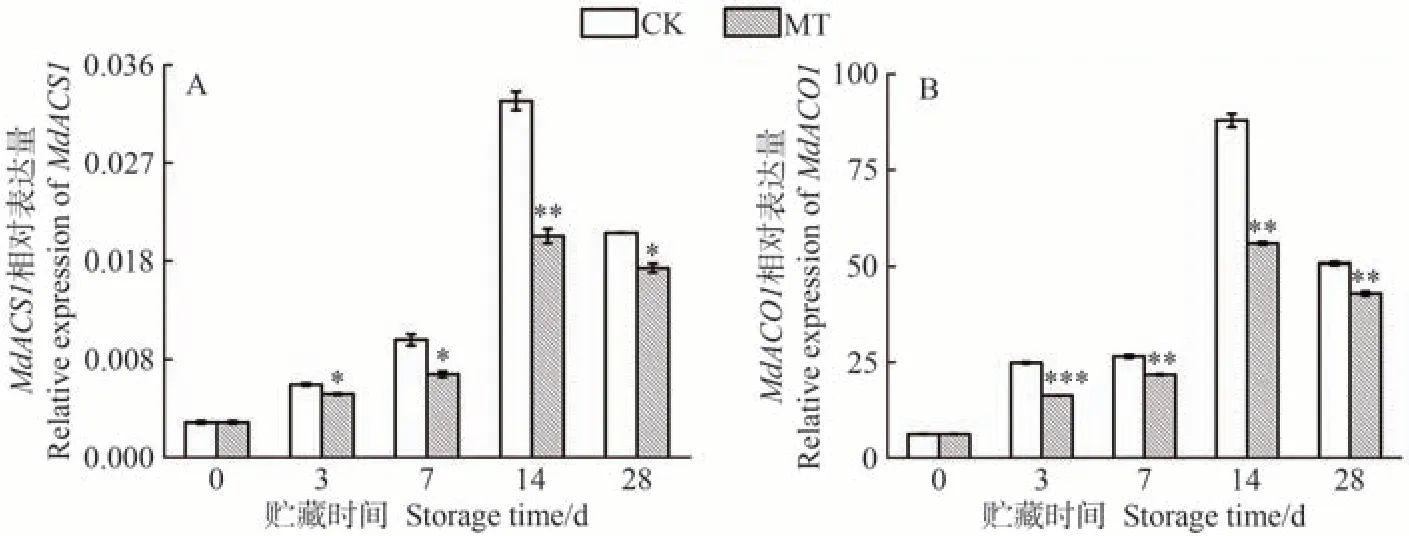
圖4 MT處理對沙果果實乙烯合成關鍵基因MdACS1(A)和MdACO1(B)相對表達量的影響Fig.4 Effects of MT treatment on the relative expression of MdACS1 (A) and MdACO1 (B) in shaguo
2.5 MT處理對沙果果實失重率的影響
沙果果實采收后,在酶的催化下通過消耗營養物質來維持生命活動會使果實組織含水量下降,直接影響其新鮮程度,進而影響商品價值。失重率越小,果實水分保持越好,品質更佳。由圖5可知,貯藏期間,CK和MT 處理組果實的失重率均持續上升,MT 處理組果實失重率始終低于CK。在貯藏早期,MT處理組與CK的失重率有顯著差異,隨著果實逐漸成熟,差異逐漸減小。貯藏17 d時,CK組果實失重率是MT處理組的1.34倍。貯藏28 d 時,CK 和MT 處理組失重率分別達到4.87%和3.68%。上述結果表明,MT處理能夠減少沙果果實在貯藏期間的失水,這與MT 處理延緩獼猴桃[16]和蜂糖李[40]果實失重率上升的研究結果相似。
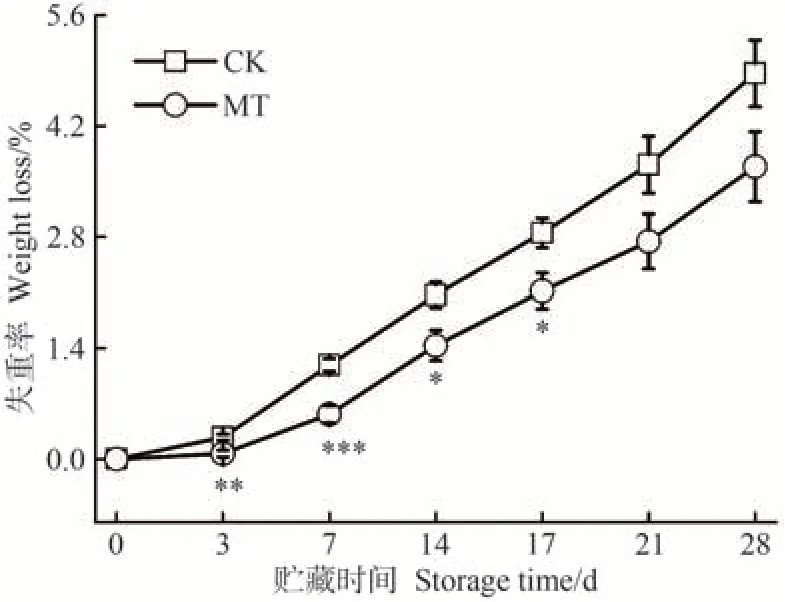
圖5 MT處理對沙果果實失重率的影響Fig.5 Effects of MT treatment on weight loss of shaguo
2.6 MT處理對沙果果實硬度的影響
沙果果實在采后貯藏期間,伴隨著果膠類物質分解及淀粉降解等成熟進程變化會出現發綿變軟、果肉質地疏松等現象[33,41],造成沙果果實品質劣變。硬度被廣泛用作成熟度和水果消費質量屬性的判斷指標。由圖6 可知,貯藏0 d 時,沙果果實的硬度為48.36 N,隨著沙果采后貯藏時間的延長,CK 和MT 處理組沙果果肉變軟趨勢加劇,到貯藏28 d,果實硬度分別為17.86和24.01 N,MT 組果實硬度是CK 組的1.35 倍。上述結果表明,MT處理采后沙果果實在維持其果實硬度上有顯著效果,有效緩解了沙果果實軟化現象的出現,在芒果[42]、富士蘋果[15]中進行MT處理也有相同表現。
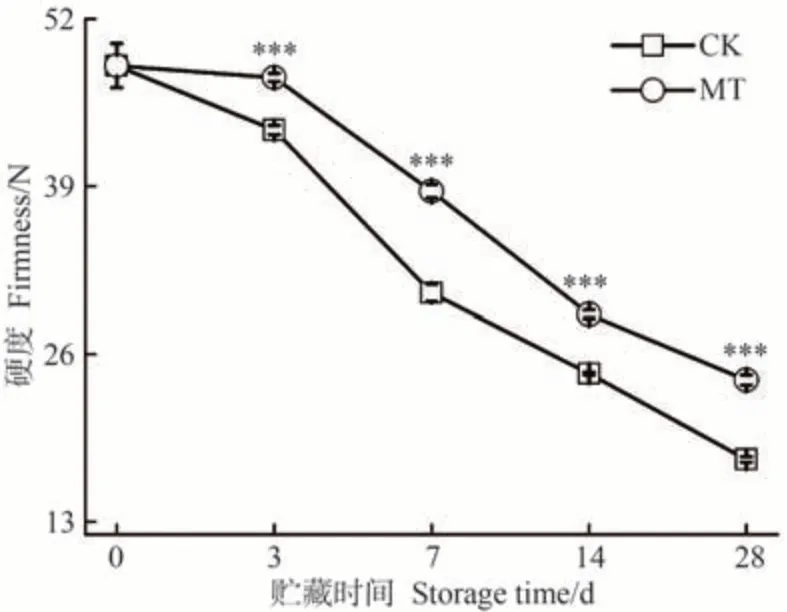
圖6 MT處理對沙果果實硬度的影響Fig.6 Effects of MT treatment on the firmness of shaguo
2.7 MT處理對沙果果實呼吸速率的影響
呼吸代謝是果蔬采后貯藏過程中最基本的生命活動,也是評價果蔬貯藏時間長短的重要指標之一,與果實中多種有機大分子的生物合成、分解代謝密切相關,能通過氧化不同的碳底物,為果實在采后貯藏期間的生命代謝提供營養物質[43]。此外,消耗果實中糖分、有機酸等有機物質會降低果實的食用品質,縮短采后貯藏時間,加快果蔬成熟衰老。由圖7可知,MT處理組能夠有效降低沙果果實的呼吸速率,與CK組變化趨勢相似,在貯藏前7 d迅速上升,中后期緩慢下降。兩組呼吸速率高峰均在貯藏7 d出現,CK組達到52.07 mL·kg-1·h-1,而MT 處理組僅為42.45 mL·kg-1·h-1,其變化趨勢與乙烯釋放速率相似。因此,MT處理可有效抑制沙果果實呼吸強度,減少有機養分的消耗,維持其貯藏品質。
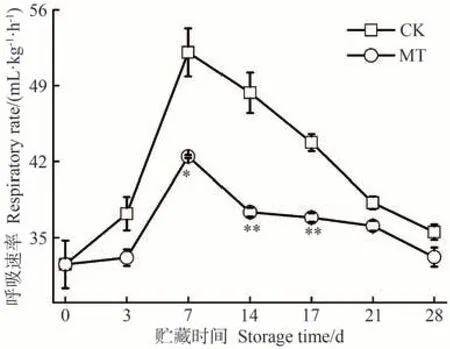
圖7 MT處理對沙果果實呼吸速率的影響Fig.7 Effects of MT treatment on the respiration rate of shaguo
3 討論
MT 作為參與多種植物活性成分合成的吲哚胺激素[1-3],在延緩果蔬衰老、維持果蔬品質方面發揮著重要作用[15-19]。Onik 等[15]使用0.1、1、10 mmol·L-1MT溶液處理不同種類蘋果,發現1 mmol·L-1MT在常溫下能更好地降低乙烯釋放量。因此本研究選用同樣濃度的MT 對沙果果實進行研究,結果發現,用1 mmol·L-1MT 處理沙果果實后,果實組織中SA 含量較對照組顯著提高。在對SA 生物合成途徑進行分析后發現,MT處理上調了SA 合成關鍵基因MdPAL和MdICS2的表達水平,促進了反式肉桂酸和異分支酸的合成,有利于果實組織中SA 的生物合成。研究表明,MT 與SA 在生物合成代謝途徑中有共同的起始物質[27]。SA 作為一種高效、無毒、低成本且無殘留的保鮮劑,在維持果蔬品質中被廣泛使用[25-26]。SA 也是乙烯生物合成中的有效抑制劑,其衍生物能通過調節乙烯生物合成和信號傳導影響果實成熟和衰老,抑制乙烯前體的合成,并減少乙烯的產生[24],在獼猴桃[29]、梨[30]和枸杞[22]中均有報道。Yuan 等[24]研究表明,SA 能通過降低王林蘋果果實中的乙烯生物合成而減緩貨架期間果實的品質劣變。因此猜測沙果果實中乙烯含量的下降可能是由于MT 處理激活了SA 生物合成,造成SA 積累,進一步抑制了乙烯的生物合成,從而減緩了采后沙果果實的衰老后熟進程,維持了硬度和貯藏品質。進一步對MT處理后沙果果實的乙烯生物合成關鍵基因MdACS1和MdACO1進行分析后發現,MT 處理組基因相對表達量較CK 顯著降低,這與富士蘋果[15]中MT 處理結果一致。說明沙果果實中乙烯釋放的下降也可能是由MT直接造成的。SA 與MT 合成關鍵酶基因表達密切相關,但由于MT 在植物中的生物合成代謝途徑尚不完全清楚,且SA在植物中的形成可能由多種因素共同調控,因此仍需進一步研究。
采摘后的果蔬仍可以通過旺盛的呼吸代謝消耗自身的營養物質,為果實的呼吸躍變準備能量[33]。呼吸躍變型果實中的呼吸速率會先呈現一個迅速上升的過程,造成果實衰老并伴隨品質的下降,主要表現為果實蒸騰作用使組織含水量下降、果實褐變與軟化、乙烯釋放高峰的出現等,影響果實的商品價值[35,41,43]。本研究表明,在CK和MT處理組的乙烯釋放高峰時期,沙果果實硬度迅速下降,說明乙烯的釋放加速了果實的軟化進程。沙果作為典型的呼吸躍變型果實,也證實了乙烯是控制沙果果實成熟衰老的關鍵所在。與CK 相比,MT處理組顯著降低了乙烯釋放的峰值,并抑制了其呼吸高峰的峰值,更有效地延緩了沙果果實中硬度的下降和失水現象的發生,與MT 處理降低獼猴桃[16]、梨[17]和黃瓜[18]失水和軟化的結果類似。上述結果表明,MT處理有效抑制了采后沙果果實中的生命活動,減緩了相關代謝進程,引發了一系列生理生化活動的變化,延緩了沙果果實的衰老,在一定程度上保持了果實的硬度和含水量。
4 結論
本研究結果表明,MT處理的沙果果實激活了SA生物合成關鍵基因MdPAL和MdICS2的表達水平,誘導了果實中SA 的積累。同時,乙烯生物合成關鍵基因MdACS1和MdACO1均受到了不同程度的下調,與乙烯含量變化趨勢一致,表明MT 通過調控MdACS1和MdACO1基因表達抑制乙烯積累,減緩乙烯信號對沙果果實的成熟作用,同時延緩硬度下降,失重率的上升以及抑制呼吸作用,有效減少營養物質消耗,維持了果實品質。綜上,MT 能夠通過上調沙果果實中MdPAL和MdICS2水平來積累SA,下調MdACS1和MdACO1表達以抑制乙烯生物合成,推遲貯藏期間果實的成熟衰老進程,維持果實品質。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
