木納格葡萄谷胱甘肽-S-轉移酶VvGST1基因克隆與序列分析
2023-09-12 09:19:36伊麗達娜迪力夏提
新疆農(nóng)業(yè)科學 2023年8期
關鍵詞:結構
王 曼,張 政,伊麗達娜·迪力夏提,吳 斌
(1.新疆農(nóng)業(yè)大學食品科學與藥學學院,烏魯木齊 830052;2.新疆農(nóng)業(yè)科學院農(nóng)產(chǎn)品貯藏加工研究所/新疆農(nóng)產(chǎn)品加工與保鮮重點實驗室,烏魯木齊 830091)
0 引 言
【研究意義】木納格(VitisviniferaL.cv Munage)是新疆主栽葡萄品種之一,其采后極易受到病原菌的侵染。隨著分子生物學和生物技術的運用,許多抗真菌基因已經(jīng)從多種植物中分離出來:如幾丁質(zhì)酶、β-1,3-葡聚糖酶和其它致病相關蛋白[1]。在植物與病原菌脅迫響應的過程中,植物進化出一系列復雜的機制來應對病原菌感染,包括病原菌感知、信號轉導和誘導特定抗性相關基因[2]。當植物受到病原微生物侵染時主要是通過防御基因的表達、氧化應激反應和代謝系統(tǒng)獲得抗性來抵抗病菌的入侵[3]。【前人研究進展】谷胱甘肽-S-轉移酶(glutathione-S-transferase,GST)具有多功能性,在植物體內(nèi)發(fā)揮解毒功能[4]。GST對于底物的特異性選擇是由兩個基本的功能域決定,分別是位于N端的GSH結合位點和位于C端的化合物底物的結合位點[5]。作為GSH途徑中的關鍵酶之一,具有催化還原性GSH和其他物質(zhì)(如疏水、親電底物)產(chǎn)生共價化合物的作用[6]。GST是一個多樣化的蛋白質(zhì)家族編碼的多基因家族。GSTs可作為替代底物催化各種反應,它們可參與生物解毒、應激響應和次級代謝等生理過程[7]。GST還是一種可溶性蛋白由兩個亞基組成,可催化GSH與疏水性物質(zhì)發(fā)生親電取代反應,轉運蛋白可將相關代謝物轉運至液泡中,進而降解有毒有害物質(zhì),從而減少對細胞的毒害作用[8]。植物生長發(fā)育調(diào)節(jié)和次級代謝產(chǎn)物的運輸?shù)冗^程需要GST家族基因的參與,其作用表現(xiàn)在抵御病原微生物侵染、排除外來化合物、抑制自身代謝產(chǎn)生的有毒物質(zhì)以及冷害等環(huán)境脅迫方面效果顯著[9]。【本研究切入點】目前,已對小麥[3]、水稻[10]和煙草[11]等植物中的GST基因進行了生物學分析并發(fā)現(xiàn)GST基因在提高植物對生物和非生物脅迫的抗性中發(fā)揮著重要的作用[12]。然而,關于GST基因參與調(diào)控果實采后抗性的研究較少,尤其是葡萄中VvGST1的分子生物學的研究鮮有報道。需分析木納格葡萄谷胱甘肽-S-轉移酶VvGST1基因克隆與序列分析。【擬解決的關鍵問題】以木納格葡萄為試材,對其VvGST1基因克隆并進行生物信息學分析,為研究VvGST1基因在鮮食葡萄中的抗病功能奠定分子基礎。
1 材料與方法
1.1 材 料
1.1.1 葡 萄
木納格葡萄采于新疆阿圖什市。挑選果梗新鮮、成熟度一致(TSS≥18%)、大小均一、無表面損傷無病蟲害的葡萄串。采收后的葡萄立即用冷鏈車運回實驗室,用液氮速凍果肉后置于-80℃保存?zhèn)溆谩?/p>
1.1.2 試 劑
瓊脂糖、2×EasyTaq?PCR SuperMix (+dye),生工生物工程(上海)股份有限公司;HyPure漿果RNA提取試劑盒,北京君諾德生物技術有限公司;M5 DL2000 DNA Marker,北京聚合美生物科技有限公司;5X All-In-One RT MasterMix (with AccuRT Genomic DNA Removal Kit),ABM公司。
1.1.3 主要儀器設備
臺式離心機,上海安亭科學儀器制造廠;PCR儀,美國Bio-Rad有限公司;漩渦混合器,海門市其林貝爾儀器制造有限公司;干式恒溫器,杭州美盛儀器有限公司;核酸定量儀,杭州奧盛有限公司;電泳儀電源、電泳槽、凝膠成像系統(tǒng),上海天能有限公司。
1.2 方 法
1.2.1 總RNA 提取與cDNA第一條鏈的合成
使用HyPure漿果RNA提取試劑盒,提取葡萄果實的總RNA。隨后,通過瓊脂糖凝膠電泳檢測RNA的完整性。cDNA合成采用FastKing cDNA第一鏈合成試劑盒。
1.2.2 葡萄果實VvGST1全長克隆
根據(jù)GenBank數(shù)據(jù)庫中公布的釀酒葡萄VvGST1基因核苷酸序列(序列號:NM_001281248.1),利用DNA man9.0 軟件設計引物,VvGST1-F:AAGAGGCTGGCAAGAAGGAAT,VvGST1-R:CTGTAATCCACCACCAAGTCATAG,引物由上海生工合成。經(jīng)PCR擴增試驗,獲得目的條帶。將獲得的目的條帶回收和核酸電泳檢測。純化后的目的片段進行連接。12 h后在4℃條件下轉化至DH5a,37℃過夜培養(yǎng),挑取菌體正常的單克隆,將擴增的菌液進行PCR鑒定,鑒定完成后每片段挑選3例陽性單克隆菌液雙向測序,測序列與參照序列進行對比,結果一致可用于后續(xù)分析。
1.2.3VvGST1基因的生物信息學分析
將測序結果通過NCBI數(shù)據(jù)庫中的BLAST工具確定完整編碼框(http://blast.ncbi.nlm.nih.gov);使用蛋白質(zhì)查找開放閱讀框ORF Finder在線對比(https://www.ncbi.nlm.nih.gov/orffinder);利用ProtParam軟件分析蛋白質(zhì)分子結構和理化特性(http://web.expasy.org/prot-param);利用Prot Scale軟件分析蛋白質(zhì)的疏水性(http://web.expasy.org/protscale/);使用SignalP軟件對蛋白質(zhì)信號肽分析(https://services.healthtech.dtu.dk/);使用TMHMM軟件對蛋白質(zhì)跨膜結構域分析(https://services.healthtech.dtu.dk/);通過Softberry在線軟件的ProtComp 9.0預測VvGST1的亞細胞定位;使用SWISS-MODEL軟件對蛋白質(zhì)三級結構預測(http://swissmodel.Expasy.org/)。
2 結果與分析
2.1VvGST1保守序列的擴增及鑒定
研究表明,葡萄基因條帶特異性好,清晰度高。所獲得的VvGST1的片段長度為719 bp,目的基因長度與預測的結果一致。該條帶滿足測序反應的要求,已擴增到目的基因片段。隨機選其中一個條帶膠回收進行后續(xù)克隆實驗。所有的測序峰圖結果是較為單一的峰圖,背景較弱無雜峰亂峰,可以用于后續(xù)的分析。圖1
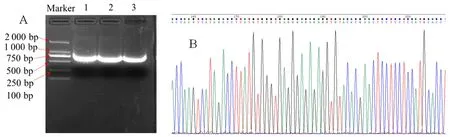
圖1 葡萄VvGST1基因PCR擴增電泳圖、測序峰圖和對比
2.2VvGST1氨基酸序列比對
研究表明,木納格葡萄中的VvGST1的氨基酸序列為保守序列。該序列與釀酒葡萄GST1(NM_001281248.1)的同源性高達97%,與梅(Prunus mume,XM_008228042.2)、甜杏仁(Prunus dulcis,XM_034357965.1)、桃(Prunus persica,XM_020561748.1)相似度均超過70%,分別為71.94%、71.78%和71.29%。克隆獲得的GST是葡萄果實中GST1基因。進一步與其他物種比對氨基酸序列,各物種間的GST基因具有較高的同源性。圖2
2.3VvGST1與其它植物GST氨基酸序列系統(tǒng)進化樹
研究表明,構建棉花(XM01260636.1)、桃(XM020561748.1)、梅(XM008228041.2)、櫻桃(XM021977290.1)、甜杏仁(XM034357965.1)和板栗(DQ673317.1)等植物的GST氨基酸序列進行系統(tǒng)進化樹,克隆的VvGST1基因編碼蛋白與棉花屬GST的親緣關系最近且屬于為Tua類GST基因。圖3
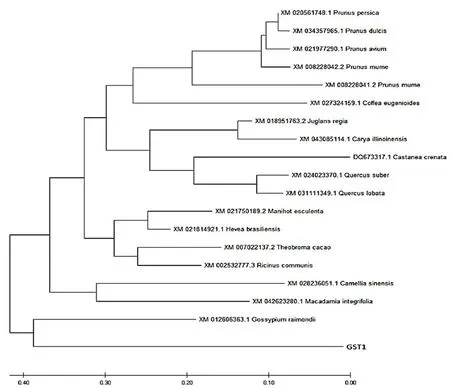
圖3 VvGST1與其它植物GST氨基酸序列系統(tǒng)進化樹
2.4 VvGST1蛋白質(zhì)的氨基酸理化特性
研究表明,VvGST1蛋白質(zhì)分子式為C1178H1799N293O321S11;相對分子質(zhì)量為25.55 kDa;原子總數(shù)3 602;等電點為6.32;推導半衰期在30 h左右。其中,不穩(wěn)定指數(shù)39.22,該蛋白質(zhì)具有穩(wěn)定性。VvGST1氨基酸數(shù)量為221個,氨基酸種類為20種。相對含量較高為亮氨酸(9.5%)、賴氨酸(9.0%)和谷氨酸(8.1%);氨基酸相對含量較低的是色氨酸(2.3%)和谷氨酰胺(1.8%)。其中帶負電荷的天冬氨酸和谷氨酰胺殘基總數(shù)是30,帶正電荷的精氨酸和賴氨酸殘基總數(shù)是28。圖4
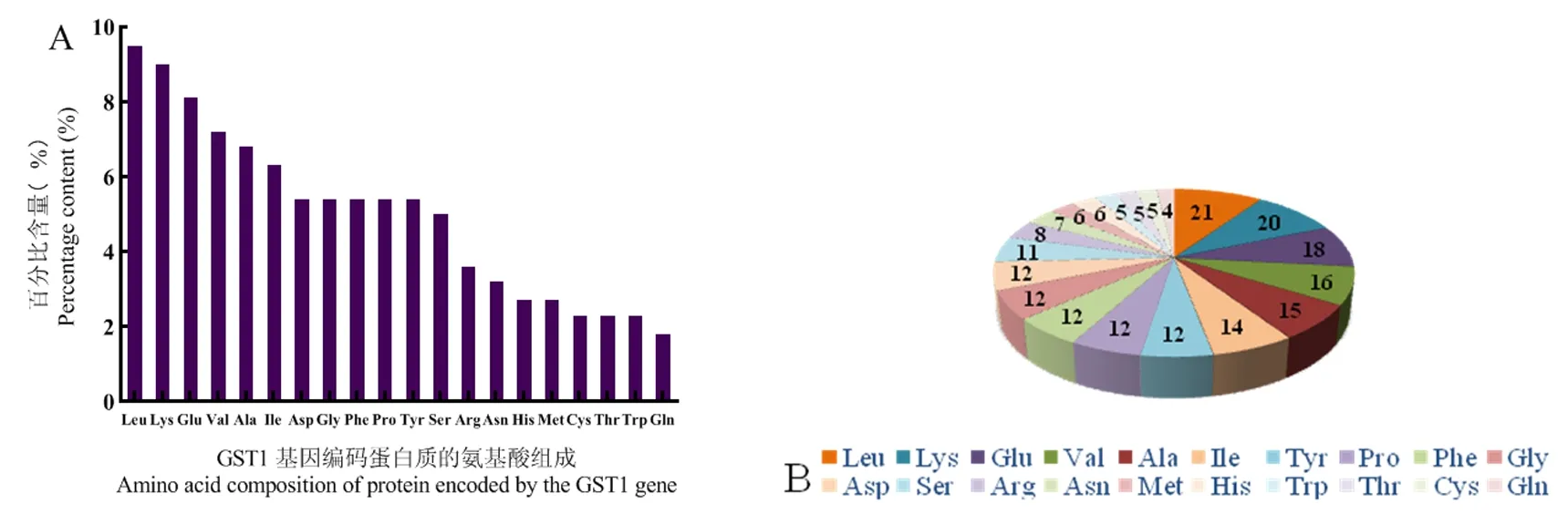
圖4 VvGST1基因編碼蛋白質(zhì)的氨基酸組成
2.5 VvGST1蛋白親疏水性
研究表明,在整個肽鏈中大多數(shù)氨基酸數(shù)值在0 ~ -2.5,分布均勻,負值越小,表示該氨基酸疏水性越強,且總親水性平均數(shù)為-0.175,脂肪指數(shù)為89.55,該葡萄果實中VvGST1為親水性蛋白。圖5

圖5 VvGST1蛋白的親疏水性
2.6 VvGST1蛋白信號肽預測
研究表明,木納格葡萄VvGST1沒有明顯的區(qū)段存在信號肽,該序列沒有預測到信號肽,意味VvGST1不會產(chǎn)生蛋白運轉。圖6

圖6 VvGST1蛋白信號肽預測
2.7 VvGST1蛋白跨膜結構域預測
研究表明,木納格葡萄VvGST1蛋白整條鏈上所有位點均沒有存在跨膜結構的可能性,整個蛋白的所有氨基酸膜外的概率均在0.9以上,存在于細胞基質(zhì)中,說明該蛋白亞單位不含跨膜結構域,是膜外蛋白,為典型的基質(zhì)蛋白。圖7
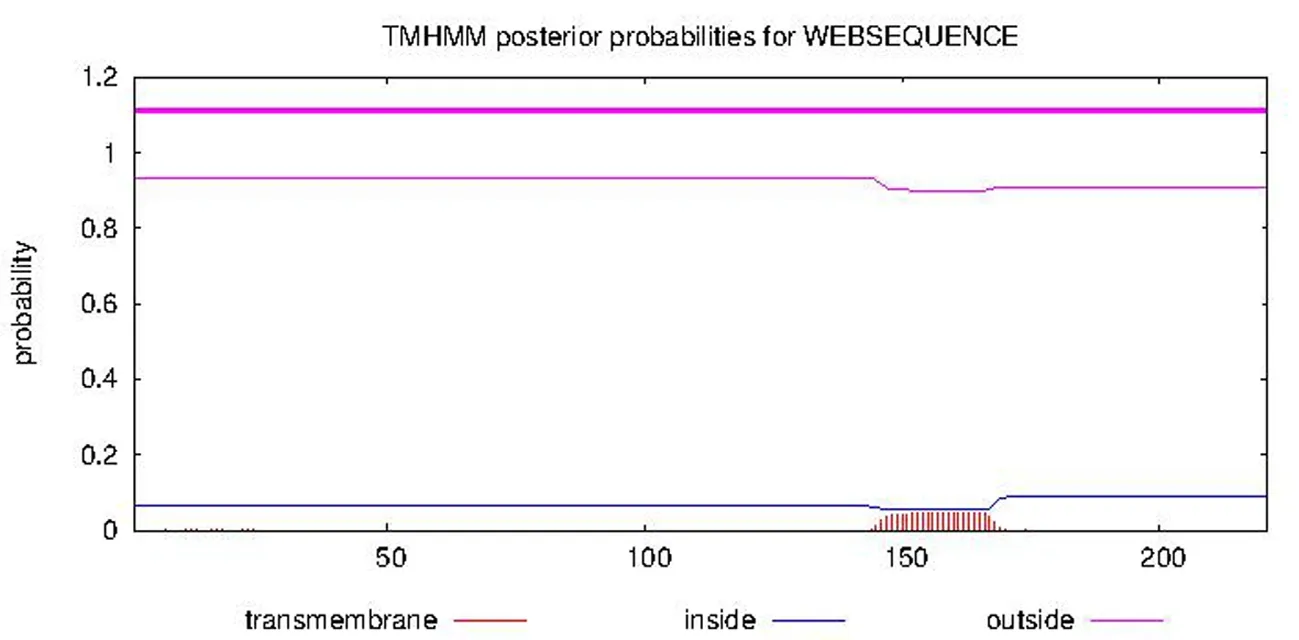
注:膜外(outside)、膜內(nèi)(inside)和跨膜區(qū)域(transmembrane)
2.8 VvGST1蛋白亞細胞定位預測
研究表明,由神經(jīng)網(wǎng)絡預測在細胞外的分值為1.0,預測VvGST1蛋白整體在細胞質(zhì)的分值為8.64,該蛋白可能是多定位的。利用MultiLocDB中進行SBLAST搜索,該蛋白質(zhì)的位置可能在細胞質(zhì)和細胞核中。表1
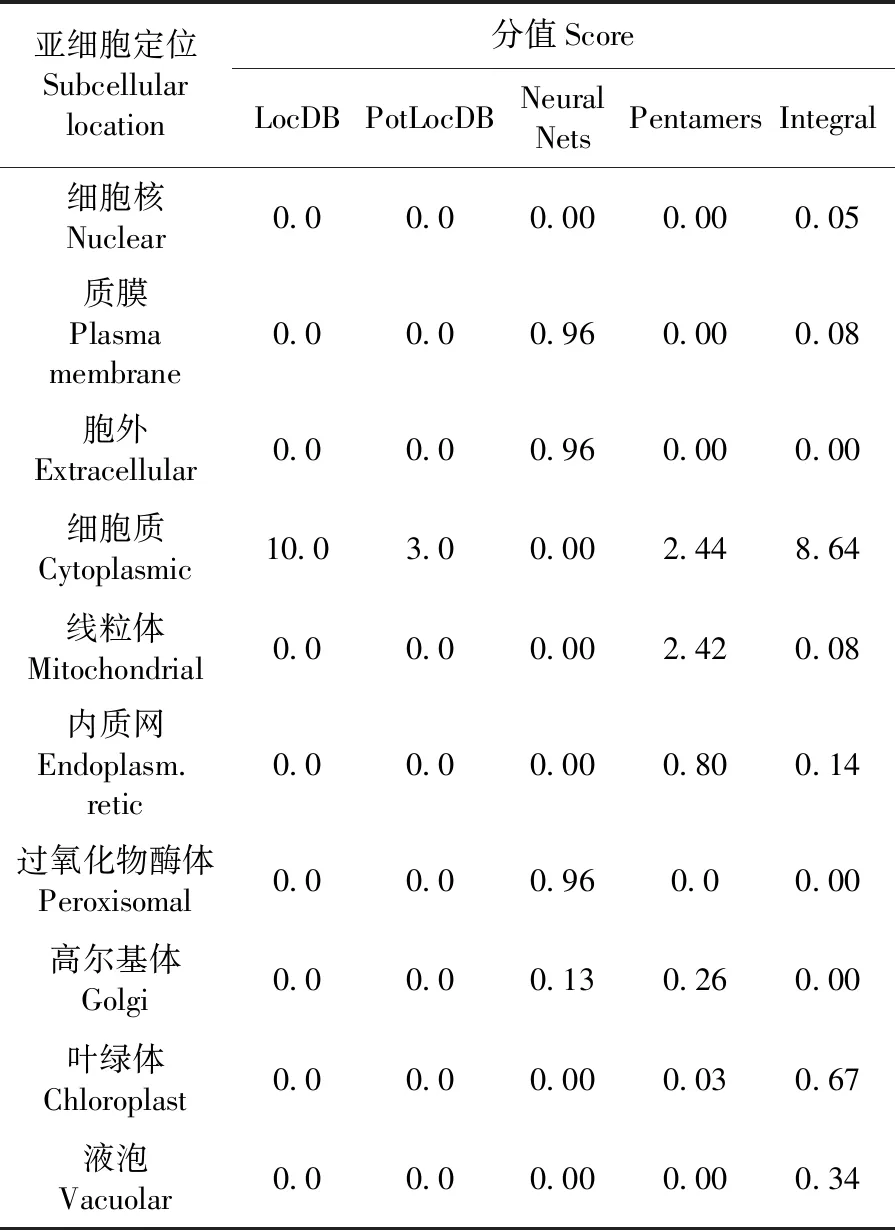
表1 VvGST1亞細胞定位預測
2.9 VvGST1蛋白二級和三級結構的構建
研究表明,VvGST1蛋白主要由4種二級結構組成,包括無規(guī)則卷曲、α-螺旋、延伸鏈和β-折疊等,其中α-螺旋是主要的二級結構占51.58%,其次無規(guī)則卷曲占29.84%,延伸鏈占13.75%,β-折疊占4.98%。圖8

注:藍色:α-螺旋;綠色:β-折疊;黃色:無規(guī)則卷曲;紅色:延伸鏈
VvGST1蛋白質(zhì)的三維結構模型類似山峰型,蛋白三維結構模型與模板的一致性為60.37%,全球性模型質(zhì)量評估(GMQE)值為0.81,越接近1,該蛋白三級結構建模質(zhì)量較好。該蛋白的三級結構主要是由α-螺旋組成,該蛋白具有穩(wěn)定的結果。圖9

圖9 VvGST1蛋白的三級結構預測
3 討 論
蛋白二級結構是指沿一定方向折疊或盤旋而形成的特定多肽結構,包括α-螺旋、β-折疊、無規(guī)則卷曲和延伸鏈等[13]。灰霉病是由灰葡萄孢引起的一種真菌性采后病害,是葡萄采后貯藏期間引起腐爛的主要病原菌之一,引起葡萄漿果采后品質(zhì)的快速下降[14]。開展抗灰霉病相關基因的挖掘對于葡萄采后抵御病原菌侵染具有重要作用。GST基因在植物響應生物與非生物脅迫方面發(fā)揮著重要作用,并具有一定的耐受能力[15]。GST家族可根據(jù)結構特征和序列的同源性分類分別是Phi、Tua、Lambda、Theta、Zeta、DHAR和TCHQD,其中植物所特有的是Phi和Tua類基因[16]。
試驗從木納格葡萄果實中成功克隆了VvGST1全長序列。該序列為完整的開放閱讀框,保守的編碼長度是719 bp,其氨基酸蛋白編碼長度與山葡萄[17]、荔枝[18]相似。VvGST1與其他物種氨基酸序列多重對比和進化樹系統(tǒng)構建可看出,研究克隆的VvGST1基因編碼蛋白與棉花屬GST的親緣關系最近且屬于Tua類GST基因。不同物種間GST的氨基酸序列在進化過程中是保守的。研究表明,氨基酸保守性越高,就越可能是功能位點,其突變對功能影響就越大[19]。
理化特性分析表明VvGST1共編碼氨基酸221個,包含20種氨基酸,pI 6.32;相對分子質(zhì)量為25.55 kDa。VvGST1蛋白不穩(wěn)定指數(shù)為39.22,表明該蛋白質(zhì)具有穩(wěn)定性。分子的親水性是指其有能力形成氫鍵的位點,并且和非極性或其他疏水性溶液對比,親水性分子更易溶解在水里面[20]。VvGST1蛋白是具有穩(wěn)定性的親水性蛋白,該蛋白不能進行膜內(nèi)外運動,不含跨膜結構域,沒有預測到信號肽。從亞細胞定位可以發(fā)現(xiàn)該蛋白存在細胞質(zhì)中與跨膜結構域預測結果一致。三級結構是指蛋白質(zhì)通過充分旋轉折疊的、并具有生物活性的、能完成生物代謝功能的一個完整球蛋白的三維立體結構[20]。VvGST1蛋白的二級結構和三維結構模型主要由α-螺旋組成,其中三級結構模型中GMQE值為0.81,當越接近1時建模質(zhì)量越好。VvGST1基因編碼蛋白質(zhì)的二級和三級結構十分穩(wěn)定。這與蓖麻克隆GST蛋白三維結構相似,該蛋白可能與蓖麻蛋白具有的功能相似[21]。葡萄VvGST1含有豐富的α-螺旋結構,其可能指導VvGST1生物合成能夠響應于生物脅迫因子或非生物脅迫因子的誘導,參與細胞對外界脅迫的抗性反應[5]。蘋果受到輪紋病侵染時VvGST1基因表達上調(diào)表達并克隆了抗蘋果輪紋病相關的MdGSTU1[9]。在水稻和小麥中分別分離出了參與植物抗病脅迫響應的GST相關基因[22];擬南芥與番茄中的多個GST被證明了能夠對真菌侵染做出響應提高抗性能力[23]。植物GSTs的表達受到非生物和生物脅迫的激活。
4 結 論
在木納格葡萄中克隆了1個GST1基因,該基因片段長度為719 bp,具有完整的開放閱讀框。VvGST1基因編碼蛋白質(zhì)的相對分子質(zhì)量為25.55 kDa;理論等電點為6.32;分子式為C1178H1799N293O321S11;原子總數(shù)3 602;理論推導半衰期為30 h左右;不穩(wěn)定指數(shù)為39.22。VvGST1基因編碼的氨基酸種類及個數(shù)分別為20種和221個。VvGST1蛋白的二級和三級結構構建分析主要是由α-螺旋組成,該蛋白具有十分穩(wěn)定的結構。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數(shù)理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
